

Willow (Salix acmophylla Boiss.) Leaf and Branch Extracts Inhibit In Vitro Sporulation of Coccidia (Eimeria spp.) from Goats
Abstract
:
1. Introduction
2. Materials and Methods
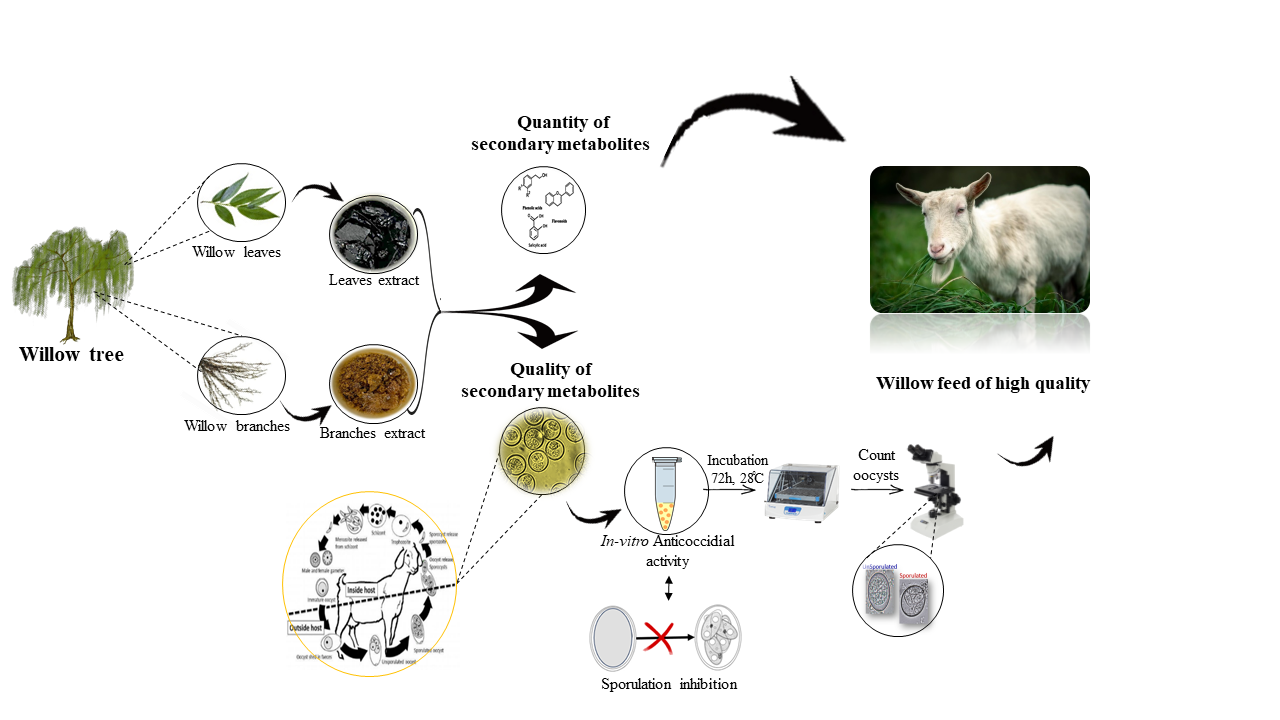
2.1. Plant Material and Sample Preparation
2.2. Quantity and Quality of Secondary Metabolites in Willow Leaves and Branches
2.2.1. Preparation of Plant Extracts
2.2.2. Phenolic Acids and Flavonoid Assays
2.2.3. Coccid Sporulation Test
2.3. Data Analysis
3. Results
3.1. Yield (%) of Ethanolic Extracts from Willow Leaves and Branches
3.2. Total Phenolic Content of Willow Leaf and Branch Extracts Obtained from the Different Genotypes
3.3. Total Flavonoid Content of Extracts from Leaves and Branches of the Different Willow Genotypes
3.4. Salicylic Acid Content of Willow Leaves and Branches
3.5. Anticoccidial Activity of Willow Extracts Obtained from Leaves and Branches at Different Concentrations of the Three Genotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lauron-Moreau, A.; Pitre, F.E.; Argus, G.W.; Labrecque, M.; Brouillet, L. Phylogenetic relationships of American willows (Salix L., Salicaceae). PLoS ONE 2015, 10, e0121965. [Google Scholar]
- Argus, G.W. Salix (Salicaceae) distribution maps and a synopsis of their classification in North America, north of Mexico. Harv. Pap. Bot. 2007, 12, 335–369. [Google Scholar] [CrossRef]
- Isebrands, J.G.; Richardson, J. Poplars and Willows: Trees for Society and the Environment; The Food and Agriculture Organization of the United Nations and CABI: Boston, MA, USA, 2014; Volume 634. [Google Scholar]
- Mahdi, J.G. Medicinal potential of willow: A chemical perspective of aspirin discovery. J. Saudi Chem. Soc. 2010, 14, 317–322. [Google Scholar] [CrossRef]
- Kuzovkina, Y.A.; Knee, M.; Quigley, M.F. Effects of soil compaction and flooding on the growth of 12 willow (Salix L.) species. J. Environ. Hortic. 2004, 22, 155–160. [Google Scholar] [CrossRef]
- Muklada, H.; Klein, J.; Glasser, T.; Dvash, L.; Azaizeh, H.; Halabi, N.; Davidovich-Rikanati, R.; Lewinsohn, E.; Landau, S.Y. Initial evaluation of willow (Salix acmophylla) irrigated with treated wastewater as a fodder crop for dairy goats. Small Rum. Res. 2017, 163, 783. [Google Scholar] [CrossRef]
- Larsen, S.U.; Lærke, P.E.; Jørgensen, U. Harvest of green willow biomass for feed—Effects of harvest time and frequency on yield, nutrient concentration, silage quality, and regrowth. Acta Agric. Scand. 2020, 70, 532–540. [Google Scholar] [CrossRef]
- Saller, R.; Melzer, J.; Felder, M. Pain relief with a proprietary extract of Willow bark in rheumatology. An Open Trial. Schweiz. Z. Für Ganzheitsmed. 2008, 20, 156–162. [Google Scholar] [CrossRef]
- Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B. Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochemistry 2011, 72, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.R.; Aboud, A.S. Antimicrobial activities of aqueous and methanolic extracts from salvia officinalis and Salix acmophylla used in the treatment of wound infection isolates. IBN AL-Haitham J. Pure Appl. Sci. 2017, 23, 25–39. [Google Scholar]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- El-Shazly, A.; El-Sayed, A.; Fikrey, E. Bioactive secondary metabolites from Salix tetrasperma Roxb. Z. Fur Naturforsch. 2012, 67, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.Y.; Chan, C.L.; Yang, Q.Q.; Li, H.B.; Zhang, D.; Ge, Y.Y.; Corke, H. Bioactive compounds and beneficial functions of sprouted grains. In Sprouted Grains; AACC International Press: St Paul, MN, USA, 2019; pp. 91–246. [Google Scholar]
- Lindroth, R.L.; Pajutee, M.S. Chemical analysis of phenolic glycosides: Art, facts, and artifacts. Oecologia 1987, 74, 144–148. [Google Scholar] [CrossRef]
- Kammerer, B.; Kahlich, R.; Biegert, C.; Gleiter, C.H.; Heide, L. HPLC-MS/MS analysis of willow bark extracts contained in pharmaceutical preparations. Phytochem. Anal. 2005, 16, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Fu, Y.; Zhang, W.; Su, G.; Liu, B.; Guo, M.; Li, F.; Liang, D.; Liu, Z.; Zhang, X.; et al. Salidroside attenuates inflammatory responses by suppressing nuclear factor-κB and mitogen-activated protein kinases activation in lipopolysaccharide-induced mastitis in mice. Inflamm. Res. 2013, 62, 9–15. [Google Scholar] [CrossRef]
- Farney, J.K.; Mamedova, L.K.; Coetzee, J.F.; Minton, J.E.; Hollis, L.C.; Bradford, B.J. Sodium salicylate treatment in early lactation increases whole-lactation milk and milk fat yield in mature dairy cows. J. Dairy Sci. 2013, 96, 7709–7718. [Google Scholar] [CrossRef]
- Nyman, T.; Julkunen-Tiitto, R. Chemical variation within and among six northern willow species. Phytochemistry 2005, 66, 2836–2843. [Google Scholar] [CrossRef]
- Li, X.; Liu, Z.; Zhang, X.F.; Wang, L.J.; Zheng, Y.N.; Yuan, C.C.; Sun, G.Z. Isolation and characterization of phenolic compounds from the leaves of Salix matsudana. Molecules 2008, 13, 1530–1537. [Google Scholar] [CrossRef] [PubMed]
- Sulaiman, G.M.; Hussien, N.N.; Marzoog, T.R.; Awad, H.A. Phenolic content, antioxidant, antimicrobial and cytotoxic activities of ethanolic extract of Salix alba. Am. J. Biochem. Biotechnol. 2013, 9, 41–46. [Google Scholar] [CrossRef]
- El-Wakil, E.; El-Sayed, S.; El-Sayed, M.M.; Abdel-Lateef, E. Identification of the chemical composition of the methanolic extract of Salix tetrasperma Roxb. using LC-ESI-MS and evaluation its potential as antioxidant agent. Pharma Chem. 2015, 7, 168–177. [Google Scholar]
- Hoste, H.; Jackson, F.; Athanasiadou, S.; Thamsborg, S.M.; Hoskin, S.O. The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 2006, 22, 253–261. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Unraveling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Hernandez, P.M.; Salem, A.Z.M.; Elghandour, M.Y.; Cipriano-Salazar, M.; Cruz-Lagunas, B.; Camacho, L.M. Anthelmintic effects of Salix babylonica L. and Leucaena leucocephala Lam. extracts in growing lambs. Trop. Anim. Health Prod. 2014, 46, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Glazer, I.; Salame, L.; Dvash, L.; Muklada, H.; Azaizeh, H.; Mreny, R.; Landau, S. Effects of tannin-rich host plants on the infection and establishment of the entomopathogenic nematode Heterorhabditis bacteriophora. J. Invertebr. Pathol. 2015, 128, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Awwad, S.; Markovics, A.; Halahlih, F.; Yazbak, A.; Haj-Zaroubi, M.; Muklada, H.; Klein, J.D.; Azaizeh, H. Effect of irrigation water source on secondary metabolites in Salix acmophylla and their potential to impair exsheathment of gastro-intestinal nematodes. Ann. Agric. Crop Sci. 2021, 6, 1079. [Google Scholar] [CrossRef]
- Rottenberg, A.; Zohary, D.; Nevo, E. Patterns of isozyme diversity and vegetative reproduction of willows in Israel. Int. J. Plant Sci. 1999, 160, 561–566. [Google Scholar] [CrossRef]
- Muklada, H.; Davidovich-Rikanati, R.; Wilkerson, D.G.; Klein, J.D.; Deutch-Traubaum, T.; Zou, J.; Awabdeh, S.; Sweidan, R.; Landau, S.Y.; Schwartz, A.; et al. Genotypic diversity in willow (Salix spp.) is associated with chemical and morphological polymorphism, suggesting human-assisted dissemination in the Eastern Mediterranean. Biochem. Syst. Ecol. 2020, 91, 104081. [Google Scholar] [CrossRef]
- Hussein Muklada, H.; Davidovich-Rikanati, R.; Awabdeh, S.; Weinberg, Z.G.; Hen, Y.; Deutch, T.; Klein, J.D.; Voet, H.; Lewinsohn, E.; Landau, S.Y. Ensiling willow (Salix acmophylla) fodder modifies the contents of plant specialized metabolites, but not nutritional attributes. Anim. Feed. Sci. Technol. 2021, 278, 115019. [Google Scholar] [CrossRef]
- Keeton, S.T.N.; Navarre, C.B. Coccidiosis in Large and Small Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bangoura, B.; Bardsley, K.D. Ruminant coccidiosis. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 187–203. [Google Scholar] [CrossRef]
- Mohamaden, W.I.; Sallam, N.H.; Abouelhassan, E.M. Prevalence of Eimeria species among sheep and goats in Suez Governorate, Egypt. Int. J. Vet. Sci. Med. 2018, 6, 65–72. [Google Scholar] [CrossRef]
- Juszczak, M.; Sadowska, N.; Udała, J. Parasites of the digestive tract of sheep and goats from organic farms in Western Pomerania, Poland Ann. Parasitol. 2019, 65, 245–250. [Google Scholar]
- Rivero-Perez, N.; Hernández-Alvarado, J.L.; Valladares-Carranza, B.; Delgadillo-Ruiz, L.; Ojeda-Ramírez, D.; Sosa-Gutiérrez, C.G.; Morales-Ubaldo, A.L.; Vega-Sanchez, V.; Zaragoza-Bastida, A. Salix babylonica L. as a Natural Anticoccidial Alternative in Growing Rabbits. Evid. Based Complement. Altern. Med. 2019, 2019, 2107231. [Google Scholar] [CrossRef] [PubMed]
- Azaizeh, H.; Halahleh, F.; Abbas, N.; Markovics, A.; Muklada, H.; Ungar, E.D.; Landau, S.Y. Polyphenols from Pistacia lentiscus and Phillyrea latifolia impair the exsheathment of gastro-intestinal nematode larvae. Vet. Parasitol. 2013, 191, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Zhishen, J.; Mengcheng, T.; Jianming, W. Determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Warrier, R.; Paul, M.; Vineetha, M. Estimation of salicylic acid in Eucalyptus leaves using spectrophotometric methods. Gen. Plant Physiol. 2013, 3, 90–97. [Google Scholar]
- Zhang, G.; Li He, L.; Hu, M. Optimized ultrasonic-assisted extraction of flavonoids from Prunella vulgaris L. and evaluation of antioxidant activities in vitro. Innov. Food Sci. Emerg. Technol. 2011, 12, 18–25. [Google Scholar] [CrossRef]
- Ekawasti, F.; Nurcahyo, W.; Wardhana, A.H.; Shibahara, T.; Tokoro, M.; Sasai, K.; Matsubayashi, M. Molecular characterization of highly pathogenic Eimeria species among beef cattle on Java Island, Indonesia. Parasitol. Int. 2019, 72, 101927. [Google Scholar] [CrossRef] [PubMed]
- Tawfeek, N.; Mahmoud, M.F.; Hamdan, D.I.; Sobeh, M.; Farrag, N.; Wink, M.; El-Shazly, A.M. Phytochemistry, pharmacology and medicinal uses of plants of the genus Salix: An updated review. Front. Pharmacol. 2021, 12, 593856. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R. Phenolic constituents of Salix: A chemotaxonomic survey of further Finnish species. Phytochemistry 1989, 28, 2115–2125. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. Herbal Medicines; Royal Pharmaceutical Society: London, UK, 2002. [Google Scholar]
- Shah, Z.A.; Hameed, A.; Ahmed, A.; Simjee, S.U.; Jabeen, A.; Ullah, A.; Shaheen, F. Cytotoxic and anti-inflammatory salicin glycosides from leaves of Salix acmophylla. Phytochem. Lett. 2016, 17, 107–113. [Google Scholar] [CrossRef]
- Donaldson, J.R.; Lindroth, R.L. Genetics, environment, and their interaction determine efficacy of chemical defense in trembling aspen. Ecology 2007, 88, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Salem, A.Z.M.; Elghandour, M.M.Y.; Kholif, A.E. Tree leaves of Salix babylonica extract as a natural anthelmintic for small-ruminant farms in a semiarid region in Mexico. Agrofor. Syst. 2016, 91, 111–122. [Google Scholar] [CrossRef]
- Jamil, M.; Aleem, M.T.; Shaukat, A.; Khan, A.; Mohsin, M.; Rehman, T.U.; Abbas, R.Z.; Saleemi, M.K.; Khatoonand, A.; Babar, W. Medicinal plants as an alternative to control poultry parasitic diseases. Life 2022, 12, 449. [Google Scholar] [CrossRef]
- Abbas, R.Z.; Saeed, Z.; Bosco, A.; Qamar, W.; Subhani, Z.; Sorin, C.M.; Kasli, M.A.F.; Munir, F. Botanical control of coccidiosis in ruminants. Pak. J. Agric. Sci. 2023, 60, 473–485. [Google Scholar]
- Soundararajan, C.; Nagarajan, K.; Satish, A.C.; Prakash, M.A. Use of tamarind seed coat powder for controlling coccidiosis in goats. Indian J. Small Rumin. 2019, 25, 247–250. [Google Scholar] [CrossRef]
- Rodríguez-Hernández, P.; Reyes-Palomo, C.; Sanz-Fernández, S.; Rufino-Moya, P.J.; Zafra, R.; Martínez-Moreno, F.J.; Rodríguez-Estévez, V.; Díaz-Gaona, C. Antiparasitic tannin-rich plants from the south of Europe for grazing livestock: A review. Animals 2023, 13, 201. [Google Scholar] [CrossRef]
- Mubashir, A.; Ghani, A.; Mubashar, A. Common medicinal plants effective in peptic ulcer treatment: A Nutritional Review. Inter. J. Agric. Biosci. 2022, 11, 70–74. [Google Scholar]
- Gorlenko, C.L.; Kiselev, H.Y.; Budanova, E.V.; Zamyatnin, A.A., Jr.; Ikryannikova, L.N. Plant secondary metabolites in the battle of drugs and drug-resistant bacteria: New heroes or worse clones of antibiotics? Antibiotics 2020, 9, 170. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Bush, K.; Harbarth, S.; Paul, M.; Rex, J.H.; Tacconelli, E.; Thwaites, G.E. Critical analysis of antibacterial agents in clinical development. Nat. Rev. Microbiol. 2020, 18, 286–298. [Google Scholar] [CrossRef]
- Landau, S.Y.; Glasser, T.A.; Zachut, M.; Klein, J.D.; Deutch-Traubman, T.; Voet, H.; Kra, G.; Davidovich-Rikanati, R. Milking performance and plant specialized metabolites in the milk of goats fed silage from willow (Salix acmophylla) irrigated with saline water. Livest. Sci. 2023, 270, 105205. [Google Scholar] [CrossRef]
- Awabdeh, S.; Sweidan, R.; Landau, S.Y. Growth performance and carcass characteristics of fattening Awassi lambs fed willow silage. Small Rumin. Res. 2022, 215, 106758. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Willow (Salix) Plant Tissue/Genotype | Extraction Yield (%) ±2Std | Total Phenols | Total Flavonoids | Salicylic Acid Content | |
|---|---|---|---|---|---|
| GAE * (mg)/Extract (g) ±2Std | TAE ** (mg)/Extract (g) ±2Std | Rutin (mg)/Extract (g) ±2Std | Salicylic Acid (mg)/Extract (g) ±2Std | ||
| Leaf 1 | 32 ± 0.00 | 72.3 ± 4.12 b *** | 64.9 ± 3.6 b | 129 ± 2.31 a | 59.8 ± 0.76 b |
| Leaf 2 | 31 ± 0.00 | 71.3 ± 2.72 b | 64.1 ± 2.45 b | 110 ± 9.25 b | 59.4 ± 2.25 b |
| Leaf 3 | 32 ± 0.01 | 83.5 ± 0.98 a | 75 ± 0.88 a | 128 ± 2.31 a | 75 ± 1.44 a |
| Branch 1 | 12 ± 0.00 | 93 ± 4.68 a | 83.5 ± 4.2 a | 210 ± 1.16 a | 40 ± 1.83 b |
| Branch 2 | 13 ± 0.01 | 78.1 ± 9.50 b | 70.1 ± 8.5 b | 165 ± 16.19 b | 44 ± 4.33 ab |
| Branch 3 | 11 ± 0.01 | 86.3 ± 2.34 b | 77.5 ± 2.1 ab | 177 ± 12.72 b | 52.1 ± 0.38 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haj-Zaroubi, M.; Mattar, N.; Awabdeh, S.; Sweidan, R.; Markovics, A.; Klein, J.D.; Azaizeh, H. Willow (Salix acmophylla Boiss.) Leaf and Branch Extracts Inhibit In Vitro Sporulation of Coccidia (Eimeria spp.) from Goats. Agriculture 2024, 14, 648. https://doi.org/10.3390/agriculture14050648
Haj-Zaroubi M, Mattar N, Awabdeh S, Sweidan R, Markovics A, Klein JD, Azaizeh H. Willow (Salix acmophylla Boiss.) Leaf and Branch Extracts Inhibit In Vitro Sporulation of Coccidia (Eimeria spp.) from Goats. Agriculture. 2024; 14(5):648. https://doi.org/10.3390/agriculture14050648
Chicago/Turabian StyleHaj-Zaroubi, Manal, Nariman Mattar, Sami Awabdeh, Rawad Sweidan, Alex Markovics, Joshua D. Klein, and Hassan Azaizeh. 2024. "Willow (Salix acmophylla Boiss.) Leaf and Branch Extracts Inhibit In Vitro Sporulation of Coccidia (Eimeria spp.) from Goats" Agriculture 14, no. 5: 648. https://doi.org/10.3390/agriculture14050648