Potential Role of Pig UCP3 in Modulating Adipocyte Browning via the Beta-Adrenergic Receptor Signaling Pathway

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Animal Ethics
2.2. In Vitro Pig Preadipocyte Isolation and Browning
2.3. RNA Preparation and Real-Time PCR
2.4. ADRB Inhibition during Browning
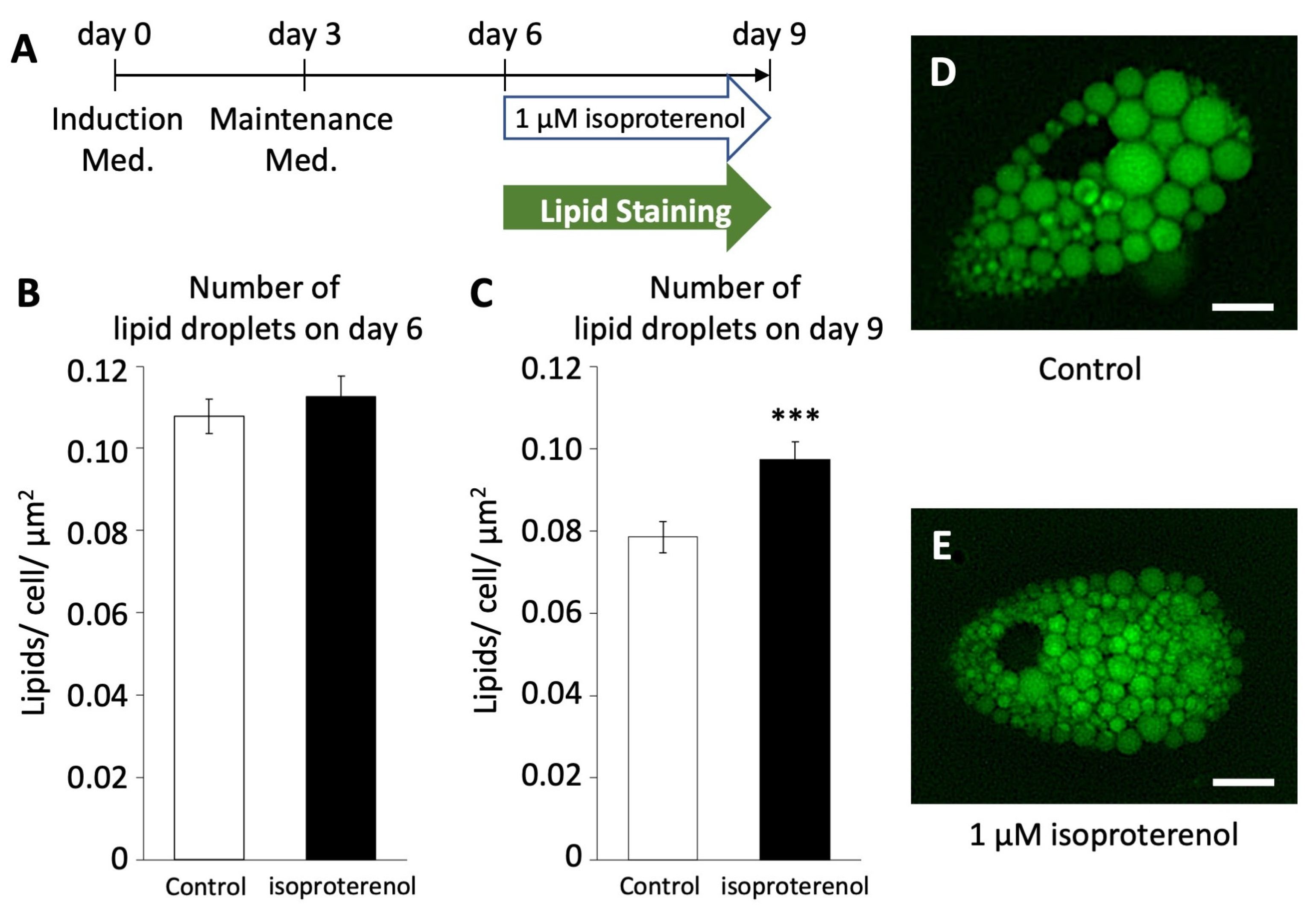
2.5. Lipid Staining after Browning
2.6. Analysis of Mitochondrial Copy Number and Expression of COX
2.7. Alignment Analysis
- Pig chromosomes 8 and 9 including UCP1 (XM_021100543.1), UCP2 (NM_214289.1), UCP3 (NM_214049.1), PPARγ (NM_214379.1), and PGC-1α (NM_213963.2);
- Human chromosomes 4 and 11 including UCP1 (NM_021833.5), UCP2 (NM_001381943.1), UCP3 (NM_003356.4), PPARγ (NM_138712.5), and PGC-1α (NM_001330751.2);
- Mouse chromosomes 7 and 8 including UCP1 (NM_009463.3), UCP2 (NM_011671.6), UCP3 (NM_009464.3), PPARγ (NM_001127330.3), and PGC-1α (NM_008904.3);
- Cattle chromosomes 7, 15, and 17 including UCP1 (NM_001166528.1), UCP2 (NM_001033611.2), UCP3 (NM_174210.1), PPARγ (NM_181024.2), and PGC-1α (NM_177945.3);
- Macaca mulatta chromosomes 2, 5, and 14 including UCP1 (XM_001090457.4), UCP2 (NM_001195393.1), UCP3 (XM_015115192.2), PPARγ (NM_001032860.1), and PGC-1α (XM_028848369.1);
- Chicken chromosomes 1, 4, and 12 including UCP3 (NM_204107.2), PPARγ (NM_001001460.2), and PGC-1α (NM_001006457.2);
- Zebrafish chromosomes 1, 7, 10, and 11 including UCP1 (NM_199523.2), UCP2 (NM_131176.1), UCP3 (NM_200353.2), PPARγ (NM_131467.1), and PGC-1α (XM_017357138.2).
2.8. Plasmids
2.9. Transfection and Luciferase Assay
2.10. Statistical Analyses
3. Results
3.1. Isoproterenol-Activated Gene Expression of PGC-1α and UCP3 in Pig Adipocytes
3.2. PPARγ and PGC-1α Activates Pig UCP3 Promoter
3.3. Gene Alignment: Human Closer to Pig Than Mouse
3.4. UCP3 Is Activated via ADRB Signaling
3.5. Isoproterenol-Induced Lipid Droplet Fragmentation in Pig Adipocytes
3.6. Enhancement of Mitochondrial Function in Pig Adipocytes by Isoproterenol
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, T.; Kuang, S. Adipocyte Dedifferentiation in Health and Diseases. Clin. Sci. 2019, 133, 2107–2119. [Google Scholar] [CrossRef] [PubMed]
- Zwick, R.K.; Guerrero-Juarez, C.F.; Horsley, V.; Plikus, M.V. Anatomical, Physiological, and Functional Diversity of Adipose Tissue. Cell Metab. 2018, 27, 68–83. [Google Scholar] [CrossRef] [PubMed]
- Bargut, T.C.L.; Souza-Mello, V.; Aguila, M.B.; Mandarim-De-Lacerda, C.A. Browning of White Adipose Tissue: Lessons from Experimental Models. Horm. Mol. Biol. Clin. Investig. 2017, 31. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.M.; Sanchez-Gurmaches, J.; Guertin, D.A. Brown Adipose Tissue Development and Metabolism. In Handbook of Experimental Pharmacology; Springer: New York, NY, USA, 2019; Volume 251, pp. 3–36. [Google Scholar]
- Heaton, J.M. The Distribution of Brown Adipose Tissue in the Human. J. Anat. 1972, 112, 35–39. [Google Scholar] [PubMed]
- Yoneshiro, T.; Saito, M. Activation and Recruitment of Brown Adipose Tissue as Anti-obesity Regimens in Humans. Ann. Med. 2015, 47, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Christian, M. Hormonal Factors in the Control of the Browning of White Adipose Tissue. Horm. Mol. Biol. Clin. Investig. 2017, 31, 20170017. [Google Scholar] [CrossRef] [PubMed]
- Patsouris, D.; Qi, P.; Abdullahi, A.; Stanojcic, M.; Chen, P.; Parousis, A.; Amini-Nik, S.; Jeschke, M.G. Burn Induces Browning of the Subcutaneous White Adipose Tissue in Mice and Humans. Cell Rep. 2015, 13, 1538–1544. [Google Scholar] [CrossRef] [PubMed]
- Pinto, Y.O.; Festuccia, W.T.L.; Magdalon, J. The Involvement of the Adrenergic Nervous System in Activating Human Brown Adipose Tissue and Browning. Hormones 2022, 21, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; You, W.; Zhou, Y.; Chen, W.; Wang, Y.; Shan, T. Cold-Induced Lipid Dynamics and Transcriptional Programs in White Adipose Tissue. BMC Biol. 2019, 17, 74. [Google Scholar] [CrossRef]
- Choi, M.; Mukherjee, S.; Yun, J.W. Trigonelline Induces Browning in 3T3-L1 White Adipocytes. Phytother. Res. 2021, 35, 1113–1124. [Google Scholar] [CrossRef]
- Ohno, H.; Shinoda, K.; Spiegelman, B.M.; Kajimura, S. PPARγ Agonists Induce a White-to-Brown Fat Conversion through Stabilization of PRDM16 Protein. Cell Metab. 2012, 15, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Conroe, H.M.; Estall, J.; Kajimura, S.; Frontini, A.; Ishibashi, J.; Cohen, P.; Cinti, S.; Spiegelman, B.M. Prdm16 Determines the Thermogenic Program of Subcutaneous White Adipose Tissue in Mice. J. Clin. Investig. 2011, 121, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, C.; Sul, H.S. Signaling Pathways Regulating Thermogenesis. Front. Endocrinol. 2021, 12, 595020. [Google Scholar] [CrossRef] [PubMed]
- Erlanson-Albertsson, C. The Role of Uncoupling Proteins in the Regulation of Metabolism. Acta Physiol. Scand. 2003, 178, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Bouillaud, F.; Alves-Guerra, M.C.; Ricquier, D. UCPs, at the Interface between Bioenergetics and Metabolism. Biochim. Biophys. Acta 2016, 1863, 2443–2456. [Google Scholar] [CrossRef] [PubMed]
- Hass, D.T.; Barnstable, C.J. Uncoupling Proteins in the Mitochondrial Defense against Oxidative Stress. Prog. Retin. Eye Res. 2021, 83, 100941. [Google Scholar] [CrossRef]
- Kaisanlahti, A.; Glumoff, T. Browning of White Fat: Agents and Implications for Beige Adipose Tissue to Type 2 Diabetes. J. Physiol. Biochem. 2019, 75, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Chandrasekera, P.C.; Pippin, J.J. Leptin- and Leptin Receptor-Deficient Rodent Models: Relevance for Human Type 2 Diabetes. Curr. Diabetes Rev. 2014, 10, 131–145. [Google Scholar] [CrossRef]
- Dufau, J.; Shen, J.X.; Couchet, M.; De Castro Barbosa, T.; Mejhert, N.; Massier, L.; Griseti, E.; Mouisel, E.; Amri, E.Z.; Lauschke, V.M.; et al. In Vitro and Ex Vivo Models of Adipocytes. Am. J. Physiol. Cell Physiol. 2021, 320, C822–C841. [Google Scholar] [CrossRef]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the Pig as a Human Biomedical Model. Sci. Transl. Med. 2021, 13, eabd5758. [Google Scholar] [CrossRef]
- Meurens, F.; Summerfield, A.; Nauwynck, H.; Saif, L.; Gerdts, V. The Pig: A Model for Human Infectious Diseases. Trends Microbiol. 2012, 20, 50–57. [Google Scholar] [CrossRef]
- Cooper, D.K.C.; Gaston, R.; Eckhoff, D.; Ladowski, J.; Yamamoto, T.; Wang, L.; Iwase, H.; Hara, H.; Tector, M.; Tector, A.J. Xenotransplantation-The Current Status and Prospects. Br. Med. Bull. 2018, 125, 5–14. [Google Scholar] [CrossRef]
- Ibrahim, Z.; Busch, J.; Awwad, M.; Wagner, R.; Wells, K.; Cooper, D.K.C. Selected Physiologic Compatibilities and Incompatibilities between Human and Porcine Organ Systems. Xenotransplantation 2006, 13, 488–499. [Google Scholar] [CrossRef]
- Simon, A.R.; Warrens, A.N.; Sykes, M. Efficacy of Adhesive Interactions in Pig-to-Human Xenotransplantation. Immunol. Today 1999, 20, 323–330. [Google Scholar] [CrossRef]
- Wang, W.; He, W.; Ruan, Y.; Geng, Q. First Pig-to-Human Heart Transplantation. Innovation 2022, 3, 100223. [Google Scholar] [CrossRef]
- Li, J.; Zhao, T.; Guan, D.; Pan, Z.; Bai, Z.; Teng, J.; Zhang, Z.; Zheng, Z.; Zeng, J.; Zhou, H.; et al. Learning Functional Conservation between Human and Pig to Decipher Evolutionary Mechanisms Underlying Gene Expression and Complex Traits. Cell Genom. 2023, 3, 100390. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Yao, Y.; Yin, H.; Cai, Z.; Wang, Y.; Bai, L.; Kern, C.; Halstead, M.; Chanthavixay, G.; Trakooljul, N.; et al. Pig Genome Functional Annotation Enhances the Biological Interpretation of Complex Traits and Human Disease. Nat. Commun. 2021, 12, 5848. [Google Scholar] [CrossRef] [PubMed]
- Berg, F.; Gustafson, U.; Andersson, L. The Uncoupling Protein 1 Gene (UCP1) Is Disrupted in the Pig Lineage: A Genetic Explanation for Poor Thermoregulation in Piglets. PLoS Genet. 2006, 2, e129. [Google Scholar] [CrossRef] [PubMed]
- Gaudry, M.J.; Jastroch, M.; Treberg, J.R.; Hofreiter, M.; Paijmans, J.L.A.; Starrett, J.; Wales, N.; Signore, A.V.; Springer, M.S.; Campbell, K.L. Inactivation of Thermogenic UCP1 as a Historical Contingency in Multiple Placental Mammal Clades. Sci. Adv. 2017, 3, e1602878. [Google Scholar] [CrossRef]
- Hou, L.; Shi, J.; Cao, L.; Xu, G.; Hu, C.; Wang, C. Pig Has No Uncoupling Protein 1. Biochem. Biophys. Res. Commun. 2017, 487, 795–800. [Google Scholar] [CrossRef]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J.; et al. Reconstitution of UCP1 Using CRISPR/Cas9 in the White Adipose Tissue of Pigs Decreases Fat Deposition and Improves Thermogenic Capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474–E9482. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Liu, H.Y.; Chang, Y.T.; Cheng, Y.H.; Mersmann, H.J.; Kuo, W.H.; Ding, S.T. Isolation and Differentiation of Adipose-Derived Stem Cells from Porcine Subcutaneous Adipose Tissues. J. Vis. Exp. 2016, 109, e53886. [Google Scholar] [CrossRef]
- Matsumoto, T.; Kano, K.; Kondo, D.; Fukuda, N.; Iribe, Y.; Tanaka, N.; Matsubara, Y.; Sakuma, T.; Satomi, A.; Otaki, M.; et al. Mature Adipocyte-Derived Dedifferentiated Fat Cells Exhibit Multilineage Potential. J. Cell Physiol. 2008, 215, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Nobusue, H.; Endo, T.; Kano, K. Establishment of a Preadipocyte Cell Line Derived from Mature Adipocytes of GFP Transgenic Mice and Formation of Adipose Tissue. Cell Tissue Res. 2008, 332, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.N.; Yang, J.Y.; England, E.; Yin, A.; Baile, C.A.; Rayalam, S. Isoproterenol Increases Uncoupling, Glycolysis, and Markers of Beiging in Mature 3T3-L1 Adipocytes. PLoS ONE 2015, 10, e0138344. [Google Scholar] [CrossRef]
- Kim, S.; Yoneda, E.; Tomita, K.; Kayano, M.; Watanabe, H.; Sasaki, M.; Shimizu, T.; Muranishi, Y. LPS Administration during Fertilization Affects Epigenetic Inheritance during Embryonic Development. Animals 2023, 13, 1135. [Google Scholar] [CrossRef]
- Ye, Y.; Zhong, H.; Dou, L.; Song, W.; Dong, C.; Lu, W.; Dong, K.; Li, K.; Li, J.; He, L.; et al. Propranolol Inhibits the Angiogenic Capacity of Hemangioma Endothelia via Blocking β-Adrenoceptor in Mast Cell. Pediatr. Res. 2022, 92, 424–429. [Google Scholar] [CrossRef]
- Pantziarka, P.; Bouche, G.; Sukhatme, V.; Meheus, L.; Rooman, I.; Sukhatme, V.P. Repurposing Drugs in Oncology (ReDO)-Propranolol as an Anti-Cancer Agent. Ecancermedicalscience 2016, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- England, R.W.; Hardy, K.L.; Kitajewski, A.M.; Wong, A.; Kitajewski, J.K.; Shawber, C.J.; Wu, J.K. Propranolol Promotes Accelerated and Dysregulated Adipogenesis in Hemangioma Stem Cells. Ann. Plast. Surg. 2014, 73 (Suppl. S1), S119–S124. [Google Scholar] [CrossRef]
- Yazawa, T.; Inaoka, Y.; Okada, R.; Mizutani, T.; Yamazaki, Y.; Usami, Y.; Kuribayashi, M.; Orisaka, M.; Umezawa, A.; Miyamoto, K. PPAR-γ Coactivator-1α Regulates Progesterone Production in Ovarian Granulosa Cells with SF-1 and LRH-1. Mol. Endocrinol. 2010, 24, 485–496. [Google Scholar] [CrossRef]
- Chhimpa, N.; Singh, N.; Puri, N.; Kayath, H.P. The Novel Role of Mitochondrial Citrate Synthase and Citrate in the Pathophysiology of Alzheimer’s Disease. J. Alzheimers Dis. 2023, 94, S453–S472. [Google Scholar] [CrossRef] [PubMed]
- Rak, M.; Bénit, P.P.; Chrétien, D.; Bouchereau, J.; Schiff, M.; El-Khoury, R.; Tzagoloff, A.; Rustin, P. Mitochondrial Cytochrome c Oxidase Deficiency. Clin. Sci. 2016, 130, 393–407. [Google Scholar] [CrossRef] [PubMed]
- Timón-Gómez, A.; Nývltová, E.; Abriata, L.A.; Vila, A.J.; Hosler, J.; Barrientos, A. Mitochondrial Cytochrome c Oxidase Biogenesis: Recent Developments. Semin. Cell Dev. Biol. 2018, 76, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Markussen, L.K.; Isidor, M.S.; Breining, P.; Andersen, E.S.; Rasmussen, N.E.; Petersen, L.I.; Pedersen, S.B.; Richelsen, B.; Hansen, J.B. Characterization of Immortalized Human Brown and White Pre-Adipocyte Cell Models from a Single Donor. PLoS ONE 2017, 12, e0185624. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.P.; Nomura, W.; Takahashi, H.; Inoue, K.; Kawada, T.; Goto, T. Methylglyoxal Attenuates Isoproterenol-Induced Increase in Uncoupling Protein 1 Expression through Activation of JNK Signaling Pathway in Beige Adipocytes. Biochem. Biophys. Rep. 2021, 28, 101127. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Kuo, C.H.; Leu, Y.L.; Wang, S.H. Corylin Reduces Obesity and Insulin Resistance and Promotes Adipose Tissue Browning through SIRT-1 and Β3-AR Activation. Pharmacol. Res. 2021, 164, 105291. [Google Scholar] [CrossRef]
- Piao, Z.; Zhai, B.; Jiang, X.; Dong, M.; Yan, C.; Lin, J.; Jin, W. Reduced Adiposity by Compensatory WAT Browning upon IBAT Removal in Mice. Biochem. Biophys. Res. Commun. 2018, 501, 807–813. [Google Scholar] [CrossRef]
- Ducharme, N.A.; Bickel, P.E. Lipid Droplets in Lipogenesis and Lipolysis. Endocrinology 2008, 149, 942–949. [Google Scholar] [CrossRef]
- Schott, M.B.; Weller, S.G.; Schulze, R.J.; Krueger, E.W.; Drizyte-Miller, K.; Casey, C.A.; McNiven, M.A. Lipid Droplet Size Directs Lipolysis and Lipophagy Catabolism in Hepatocytes. J. Cell Biol. 2019, 218, 3320–3335. [Google Scholar] [CrossRef]
- Lin, J.; Cao, C.; Tao, C.; Ye, R.; Dong, M.; Zheng, Q.; Wang, C.; Jiang, X.; Qin, G.; Yan, C.; et al. Cold Adaptation in Pigs Depends on UCP3 in Beige Adipocytes. J. Mol. Cell Biol. 2017, 9, 364–375. [Google Scholar] [CrossRef]
- Cypess, A.M.; Weiner, L.S.; Roberts-Toler, C.; Franquet Elía, E.F.; Kessler, S.H.; Kahn, P.A.; English, J.; Chatman, K.; Trauger, S.A.; Doria, A.; et al. Activation of Human Brown Adipose Tissue by a Β3-Adrenergic Receptor Agonist. Cell Metab. 2015, 21, 33–38. [Google Scholar] [CrossRef]
- Mottillo, E.P.; Balasubramanian, P.; Lee, Y.H.; Weng, C.; Kershaw, E.E.; Granneman, J.G. Coupling of Lipolysis and de Novo Lipogenesis in Brown, Beige, and White Adipose Tissues during Chronic Β3-Adrenergic Receptor Activation. J. Lipid Res. 2014, 55, 2276–2286. [Google Scholar] [CrossRef]
- Mukherjee, S.; Yun, J.W. Prednisone Stimulates White Adipocyte Browning via Β3-AR/P38 MAPK/ERK Signaling Pathway. Life Sci. 2022, 288, 120204. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Tian, S.; Lin, Y.; Tang, Q.; Zhou, X.; Li, D.; Yeung, C.K.L.; Che, T.; Jin, L.; et al. Comprehensive Variation Discovery and Recovery of Missing Sequence in the Pig Genome Using Multiple De Novo Assemblies. Genome Res. 2017, 27, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, C.L.; Csikasz, R.I.; Chernogubova, E.; Yamamoto, D.L.; Hogberg, H.T.; Amri, E.Z.; Hutchinson, D.S.; Bengtsson, T. β₁-Adrenergic Receptors Increase UCP1 in Human MADS Brown Adipocytes and Rescue Cold-Acclimated β₃-Adrenergic Receptor-Knockout Mice via Nonshivering Thermogenesis. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1108–E1118. [Google Scholar] [CrossRef]
- Riis-Vestergaard, M.J.; Richelsen, B.; Bruun, J.M.; Li, W.; Hansen, J.B.; Pedersen, S.B. Beta-1 and Not Beta-3 Adrenergic Receptors May Be the Primary Regulator of Human Brown Adipocyte Metabolism. J. Clin. Endocrinol. Metab. 2020, 105, E994–E1005. [Google Scholar] [CrossRef]
- Pohl, E.E.; Rupprecht, A.; Macher, G.; Hilse, K.E. Important Trends in UCP3 Investigation. Front. Physiol. 2019, 10, 470. [Google Scholar] [CrossRef]
- Gong, D.W.; Monemdjou, S.; Gavrilova, O.; Leon, L.R.; Marcus-Samuels, B.; Chou, C.J.; Everett, C.; Kozak, L.P.; Li, C.; Deng, C.; et al. Lack of Obesity and Normal Response to Fasting and Thyroid Hormone in Mice Lacking Uncoupling Protein-3. J. Biol. Chem. 2000, 275, 16251–16257. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Puig, A.J.; Grujic, D.; Zhang, C.Y.; Hagen, T.; Boss, O.; Ido, Y.; Szczepanik, A.; Wade, J.; Mootha, V.; Cortright, R.; et al. Energy Metabolism in Uncoupling Protein 3 Gene Knockout Mice. J. Biol. Chem. 2000, 275, 16258–16266. [Google Scholar] [CrossRef]
- Nabben, M.; van Bree, B.W.J.; Lenaers, E.; Hoeks, J.; Hesselink, M.K.C.; Schaart, G.; Gijbels, M.J.J.; Glatz, J.F.C.; da Silva, G.J.J.; de Windt, L.J.; et al. Lack of UCP3 Does Not Affect Skeletal Muscle Mitochondrial Function under Lipid-Challenged Conditions, but Leads to Sudden Cardiac Death. Basic. Res. Cardiol. 2014, 109, 447. [Google Scholar] [CrossRef]
- Cortes de Oliveira, C.; Nicoletti, C.F.; Pinhel, M.A.S.; de Oliveira, B.A.P.; Quinhoneiro, D.C.G.; Noronha, N.Y.; Fassini, P.G.; Marchini, J.S.; da Silva Júnior, W.A.; Salgado Júnior, W.; et al. Influence of Expression of UCP3, PLIN1 and PPARG2 on the Oxidation of Substrates after Hypocaloric Dietary Intervention. Clin. Nutr. 2018, 37, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, C.; Sibille, B.; Solanes, G.; Darimont, C.; Macé, K.; Villarroya, F.; Gómez-Foix, A.M. Overexpression of UCP3 in Cultured Human Muscle Lowers Mitochondrial Membrane Potential, Raises ATP/ADP Ratio, and Favors Fatty Acid vs. Glucose Oxidation. FASEB J. 2001, 15, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Dridi, S.; Onagbesan, O.; Swennen, Q.; Buyse, J.; Decuypere, E.; Taouis, M. Gene Expression, Tissue Distribution and Potential Physiological Role of Uncoupling Protein in Avian Species. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2004, 139, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Emre, Y.; Hurtaud, C.; Ricquier, D.; Bouillaud, F.; Hughes, J.; Criscuolo, F. Avian UCP: The Killjoy in the Evolution of the Mitochondrial Uncoupling Proteins. J. Mol. Evol. 2007, 65, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Joubert, R.; Métayer Coustard, S.M.; Swennen, Q.; Sibut, V.; Crochet, S.; Cailleau-Audouin, E.; Buyse, J.; Decuypere, E.; Wrutniak-Cabello, C.; Cabello, G.; et al. The Beta-Adrenergic System Is Involved in the Regulation of the Expression of Avian Uncoupling Protein in the Chicken. Domest. Anim. Endocrinol. 2010, 38, 115–125. [Google Scholar] [CrossRef]
- Sotome, R.; Hirasawa, A.; Kikusato, M.; Amo, T.; Furukawa, K.; Kuriyagawa, A.; Watanabe, K.; Collin, A.; Shirakawa, H.; Hirakawa, R.; et al. In Vivo Emergence of Beige-Like Fat in Chickens as Physiological Adaptation to Cold Environments. Amino Acids 2021, 53, 381–393. [Google Scholar] [CrossRef]
 ) control, (
) control, ( ) 0.01 µM, (
) 0.01 µM, ( ) 0.1 µM, (
) 0.1 µM, ( ) 1 µM, (
) 1 µM, ( ) 10 µM, and (
) 10 µM, and ( ) 100 µM. The values are shown as mean ± SEM. * p < 0.05 and ** p < 0.01.
) control, () 0.01 µM, () 0.1 µM, () 1 µM, () 10 µM, and () 100 µM. The values are shown as mean ± SEM. * p < 0.05 and ** p < 0.01.
) 100 µM. The values are shown as mean ± SEM. * p < 0.05 and ** p < 0.01.
) control, () 0.01 µM, () 0.1 µM, () 1 µM, () 10 µM, and () 100 µM. The values are shown as mean ± SEM. * p < 0.05 and ** p < 0.01.





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, S.; Yazawa, T.; Koide, A.; Yoneda, E.; Aoki, R.; Okazaki, T.; Tomita, K.; Watanabe, H.; Muroi, Y.; Testuka, M.; et al. Potential Role of Pig UCP3 in Modulating Adipocyte Browning via the Beta-Adrenergic Receptor Signaling Pathway. Biology 2024, 13, 284. https://doi.org/10.3390/biology13050284
Kim S, Yazawa T, Koide A, Yoneda E, Aoki R, Okazaki T, Tomita K, Watanabe H, Muroi Y, Testuka M, et al. Potential Role of Pig UCP3 in Modulating Adipocyte Browning via the Beta-Adrenergic Receptor Signaling Pathway. Biology. 2024; 13(5):284. https://doi.org/10.3390/biology13050284
Chicago/Turabian StyleKim, Sangwoo, Takashi Yazawa, Akari Koide, Erina Yoneda, Risa Aoki, Tatsuki Okazaki, Kisaki Tomita, Hiroyuki Watanabe, Yoshikage Muroi, Masafumi Testuka, and et al. 2024. "Potential Role of Pig UCP3 in Modulating Adipocyte Browning via the Beta-Adrenergic Receptor Signaling Pathway" Biology 13, no. 5: 284. https://doi.org/10.3390/biology13050284