Knockdown H19 Accelerated iPSCs Reprogramming through Epigenetic Modifications and Mesenchymal-to-Epithelial Transition

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Construction of H19-shRNA Expression Vector and Lentiviral Infection
2.3. Generation of iPS Cells
2.4. RT-PCR and RT-qPCR Expression Analysis
2.5. Alkaline Phosphatase Staining and Cell Proliferation Assay
2.6. Flow Cytometric Assay
2.7. Bisulfite Genomic Sequencing
2.8. In Vivo and In Vitro Differentiation
2.9. Statistical Analysis
3. Results
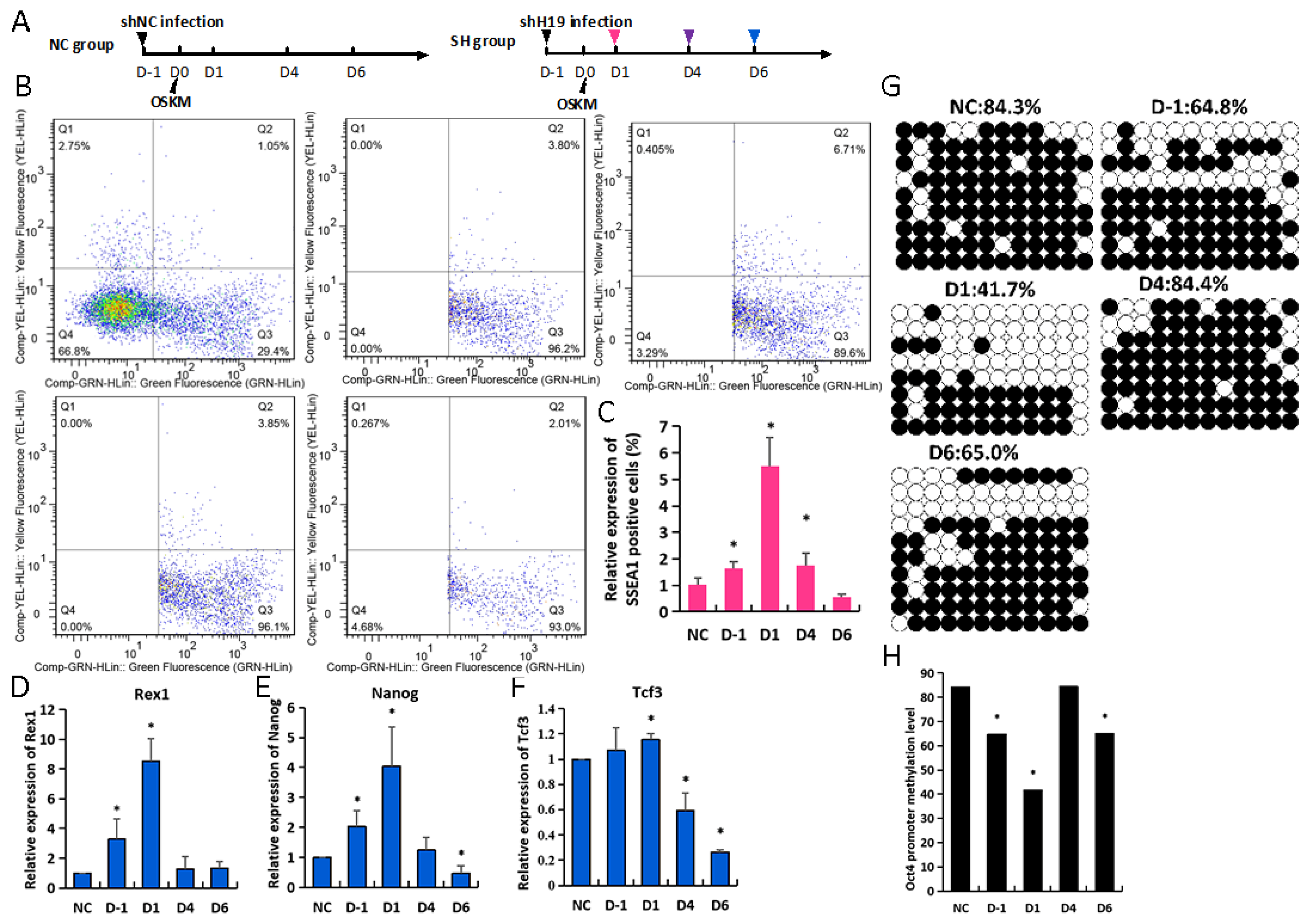
3.1. H19 Knockdown Contributes to iPSCs Reprogramming
3.2. H19 Knockdown Facilitates Expression of Pluripotent Genes during iPSCs Reprogramming
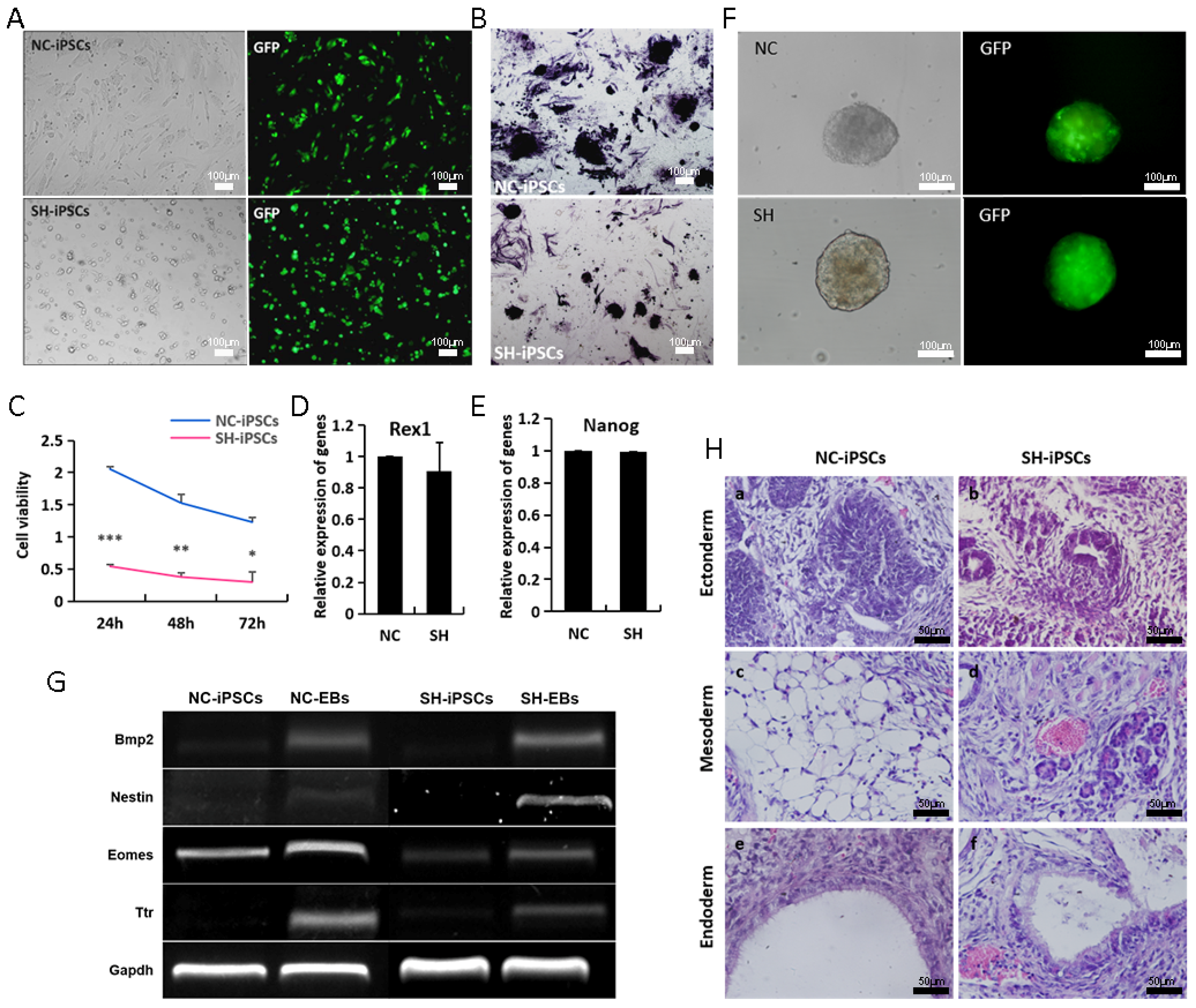
3.3. Dynamic Loss of H19 Leads to Diverse Reprogramming Efficiency
3.4. H19-Depleted iPSCs Exhibit Reduced Self-Renewal
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef]
- Wernig, M.; Meissner, A.; Foreman, R.; Brambrink, T.; Ku, M.; Hochedlinger, K.; Bernstein, B.E.; Jaenisch, R. In vitro reprogramming of fibroblasts into a pluripotent ES-cell-like state. Nature 2007, 448, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Hou, P.; Li, Y.; Zhang, X.; Liu, C.; Guan, J.; Li, H.; Zhao, T.; Ye, J.; Yang, W.; Liu, K.; et al. Pluripotent stem cells induced from mouse somatic cells by small-molecule compounds. Science 2013, 341, 651–654. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, I.M.; Fleischer, A.; Bachiller, D. Generation of mouse and human induced pluripotent stem cells (iPSC) from primary somatic cells. Stem Cell Rev. Rep. 2013, 9, 435–450. [Google Scholar] [CrossRef]
- Chang, S.; Fulmer, D.; Hur, S.K.; Thorvaldsen, J.L.; Li, L.; Lan, Y.; Rhon-Calderon, E.A.; Leu, N.A.; Chen, X.; Epstein, J.A.; et al. Dysregulated H19/Igf2 expression disrupts cardiac-placental axis during development of Silver-Russell syndrome-like mouse models. Elife 2022, 11, e78754. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Fan, Y.; Qin, C.; Xia, X.; Johnson, J.; Kallen, A.N. Absence of the long noncoding RNA H19 results in aberrant ovarian STAR and progesterone production. Mol. Cell Endocrinol. 2019, 490, 15–20. [Google Scholar] [CrossRef]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef]
- Martinet, C.; Monnier, P.; Louault, Y.; Benard, M.; Gabory, A.; Dandolo, L. H19 controls reactivation of the imprinted gene network during muscle regeneration. Development 2016, 143, 962–971. [Google Scholar] [CrossRef]
- Ogoyama, M.; Ohkuchi, A.; Takahashi, H.; Zhao, D.; Matsubara, S.; Takizawa, T. LncRNA H19-Derived miR-675-5p Accelerates the Invasion of Extravillous Trophoblast Cells by Inhibiting GATA2 and Subsequently Activating Matrix Metalloproteinases. Int. J. Mol. Sci. 2021, 22, 1237. [Google Scholar] [CrossRef]
- Park, K.S.; Rahat, B.; Lee, H.C.; Yu, Z.X.; Noeker, J.; Mitra, A.; Kean, C.M.; Knutsen, R.H.; Springer, D.; Gebert, C.M.; et al. Cardiac pathologies in mouse loss of imprinting models are due to misexpression of H19 long noncoding RNA. Elife 2021, 10, e67250. [Google Scholar] [CrossRef]
- Xu, J.; Xia, Y.; Zhang, H.; Guo, H.; Feng, K.; Zhang, C. Overexpression of long non-coding RNA H19 promotes invasion and autophagy via the PI3K/AKT/mTOR pathways in trophoblast cells. Biomed. Pharmacother. 2018, 101, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.; Piekos, S.C.; Chen, L.; Mishra, S.; Zhong, X.B. The role of H19, a long non-coding RNA, in mouse liver postnatal maturation. PLoS ONE 2017, 12, e0187557. [Google Scholar] [CrossRef] [PubMed]
- Wesemann, D.R.; Portuguese, A.J.; Magee, J.M.; Gallagher, M.P.; Zhou, X.; Panchakshari, R.A. Alt FW: Reprogramming IgH isotype-switched B cells to functional-grade induced pluripotent stem cells. Proc. Natl. Acad. Sci. USA 2012, 109, 13745–13750. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z. Igf2-H19, an imprinted tandem gene, is an important regulator of embryonic development, a guardian of proliferation of adult pluripotent stem cells, a regulator of longevity, and a ‘passkey’ to cancerogenesis. Folia Histochem. Cytobiol. 2012, 50, 171–179. [Google Scholar] [CrossRef]
- Liu, W.; Yin, J.; Kou, X.; Jiang, Y.; Gao, H.; Zhao, Y.; Huang, B.; He, W.; Wang, H.; Han, Z.; et al. Asymmetric reprogramming capacity of parental pronuclei in mouse zygotes. Cell Rep. 2014, 6, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Xie, Z.; Yin, Q.; Dong, R.; Yang, S.; Wu, Y.; Yang, L.; Li, J. Parthenogenetic haploid embryonic stem cells efficiently support mouse generation by oocyte injection. Cell Res. 2016, 26, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhao, Q.; Feng, Y.; Li, N.; Gu, Y.; Sun, R.; Duan, L.; Wu, Y.; Shan, Z.; Lei, L. Embryonic germ cell extracts erase imprinted genes and improve the efficiency of induced pluripotent stem cells. Sci. Rep. 2018, 8, 10955. [Google Scholar] [CrossRef] [PubMed]
- Zeira, E.; Abramovitch, R.; Meir, K.; Even Ram, S.; Gil, Y.; Bulvik, B.; Bromberg, Z.; Levkovitch, O.; Nahmansson, N.; Adar, R.; et al. The knockdown of H19lncRNA reveals its regulatory role in pluripotency and tumorigenesis of human embryonic carcinoma cells. Oncotarget 2015, 6, 34691–34703. [Google Scholar] [CrossRef] [PubMed]
- Stadtfeld, M.; Apostolou, E.; Akutsu, H.; Fukuda, A.; Follett, P.; Natesan, S.; Kono, T.; Shioda, T.; Hochedlinger, K. Aberrant silencing of imprinted genes on chromosome 12qF1 in mouse induced pluripotent stem cells. Nature 2010, 465, 175–181. [Google Scholar] [CrossRef]
- Cai, X.; Cullen, B.R. The imprinted H19 noncoding RNA is a primary microRNA precursor. RNA 2007, 13, 313–316. [Google Scholar] [CrossRef]
- Hashemi, M.; Moosavi, M.S.; Abed, H.M.; Dehghani, M.; Aalipour, M.; Heydari, E.A.; Behroozaghdam, M.; Entezari, M.; Salimimoghadam, S.; Gunduz, E.S.; et al. Long non-coding RNA (lncRNA) H19 in human cancer: From proliferation and metastasis to therapy. Pharmacol. Res. 2022, 184, 106418. [Google Scholar] [CrossRef] [PubMed]
- Shermane Lim, Y.W.; Xiang, X.; Garg, M.; Le, M.T.; Li-Ann Wong, A.; Wang, L.; Goh, B.C. The double-edged sword of H19 lncRNA: Insights into cancer therapy. Cancer Lett. 2021, 500, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, E.A.; Cheeran, E.A.; Salamah, J.; Lee, B.K. Long noncoding RNA H19 in ovarian biology and placenta development. Cell Biochem. Funct. 2024, 42, e3907. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, H.; Miyajima, Y.; Fukamizu, A.; Tanimoto, K. Orientation of mouse H19 ICR affects imprinted H19 gene expression through promoter methylation-dependent and -independent mechanisms. Commun. Biol. 2021, 4, 1410. [Google Scholar] [CrossRef]
- Luo, H.; Li, X.; Tian, G.G.; Li, D.; Hou, C.; Ding, X.; Hou, L.; Lyu, Q.; Yang, Y.; Cooney, A.J.; et al. Offspring production of ovarian organoids derived from spermatogonial stem cells by defined factors with chromatin reorganization. J. Adv. Res. 2021, 33, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Tian, E.; Chen, Z.X.; Sun, G.; Ye, P.; Yang, S.; Lu, D.; Xie, J.; Ho, T.V.; Tsark, W.M.; et al. Identification of Oct4-activating compounds that enhance reprogramming efficiency. Proc. Natl. Acad. Sci. USA 2012, 109, 20853–20858. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhou, A.; Huang, Y.; Zhang, Y.; Xu, Y.; Shao, K.; Ning, X. N-Cadherin Nanoantagonist Driven Mesenchymal-to-Epithelial Transition in Fibroblasts for Improving Reprogramming Efficiency. Nano Lett. 2021, 21, 5540–5546. [Google Scholar] [CrossRef] [PubMed]
- Pei, D.; Shu, X.; Gassama-Diagne, A.; Thiery, J.P. Mesenchymal-epithelial transition in development and reprogramming. Nat. Cell Biol. 2019, 21, 44–53. [Google Scholar] [CrossRef]
- Li, R.; Liang, J.; Ni, S.; Zhou, T.; Qing, X.; Li, H.; He, W.; Chen, J.; Li, F.; Zhuang, Q.; et al. A mesenchymal-to-epithelial transition initiates and is required for the nuclear reprogramming of mouse fibroblasts. Cell Stem Cell 2010, 7, 51–63. [Google Scholar] [CrossRef]
- Lv, M.; Zhong, Z.; Huang, M.; Tian, Q.; Jiang, R.; Chen, J. lncRNA H19 regulates epithelial-mesenchymal transition and metastasis of bladder cancer by miR-29b-3p as competing endogenous RNA. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1887–1899. [Google Scholar] [CrossRef]
- Peperstraete, E.; Lecerf, C.; Collette, J.; Vennin, C.; Raby, L.; Volkel, P.; Angrand, P.O.; Winter, M.; Bertucci, F.; Finetti, P.; et al. Enhancement of Breast Cancer Cell Aggressiveness by lncRNA H19 and its Mir-675 Derivative: Insight into Shared and Different Actions. Cancers 2020, 12, 1730. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; Wu, X.C.; Zhang, M.D.; Weng, M.Z.; Zhou, D.; Quan, Z.W. Upregulation of H19 indicates a poor prognosis in gallbladder carcinoma and promotes epithelial-mesenchymal transition. Am. J. Cancer Res. 2016, 6, 15–26. [Google Scholar] [PubMed]
- Wu, Y.; Zhou, Y.; He, J.; Sun, H.; Jin, Z. Long non-coding RNA H19 mediates ovarian cancer cell cisplatin-resistance and migration during EMT. Int. J. Clin. Exp. Pathol. 2019, 12, 2506–2515. [Google Scholar] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, R.; Zhang, X.; Gong, T.; Zhang, Y.; Wang, Q.; He, C.; Ju, J.; Jin, C.; Ding, W.; Gao, J.; et al. Knockdown H19 Accelerated iPSCs Reprogramming through Epigenetic Modifications and Mesenchymal-to-Epithelial Transition. Biomolecules 2024, 14, 509. https://doi.org/10.3390/biom14050509
Sun R, Zhang X, Gong T, Zhang Y, Wang Q, He C, Ju J, Jin C, Ding W, Gao J, et al. Knockdown H19 Accelerated iPSCs Reprogramming through Epigenetic Modifications and Mesenchymal-to-Epithelial Transition. Biomolecules. 2024; 14(5):509. https://doi.org/10.3390/biom14050509
Chicago/Turabian StyleSun, Ruizhen, Ximei Zhang, Tiantian Gong, Yue Zhang, Qi Wang, Chenyao He, Jielan Ju, Chunmiao Jin, Wenxin Ding, Jingnan Gao, and et al. 2024. "Knockdown H19 Accelerated iPSCs Reprogramming through Epigenetic Modifications and Mesenchymal-to-Epithelial Transition" Biomolecules 14, no. 5: 509. https://doi.org/10.3390/biom14050509