
Is the A-1 Pigment in Photosystem I Part of P700? A (P700+–P700) FTIR Difference Spectroscopy Study of A-1 Mutants

 ,
,
Abstract
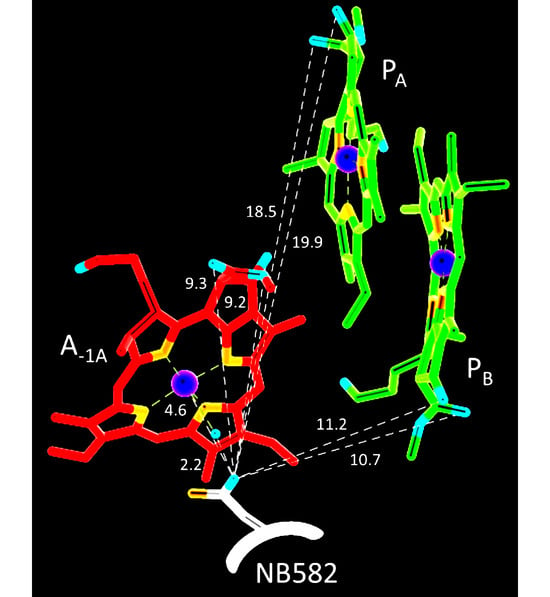
:
1. Introduction
2. Results and Discussion
2.1. Infrared Band Assignments
2.2. Bands Associated with A-1B
2.2.1. A C=O Group of Asn
2.2.2. Amide (Protein) Vibrations
2.2.3. The 131-Keto C=O Group of A-1B
2.3. Bands Associated with A-1A
2.4. 133-Ester C=O Bands of A-1
- In WT PSI, the 131-keto C=O group of A-1B is found at 1697 cm−1 and upshifts 13 cm−1 to 1710 cm−1 upon cation formation. When Asn at A600 is changed to Met, the 1697(−)/1710(+) cm−1 difference band downshifts 16/19 cm−1 to 1683(−)/1691(+) cm−1, respectively.
- In WT PSI, the 131-keto C=O group of A-1A is found at 1692 cm−1 and upshifts by 7 cm−1 to 1699 cm−1 upon cation formation. When Asn at B582 is changed to Met, the 1692(−)/1699(+) cm−1 difference band downshifts 13/7 cm−1 to 1679(−)/1692(+) cm−1.
2.5. Additivity of Mutation-Induced Spectral Changes
3. Materials and Methods
3.1. PSI Particle Preparation
3.2. FTIR Sample Preparation
3.3. FTIR Spectroscopy
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Walker, D. Energy, Plants and Man and Like Clockwork, 2nd ed.; Oxygraphics, Brighton, East Susse: Mill Valley, CA, USA, 1993. [Google Scholar]
- Vassiliev, I.R.; Jung, Y.S.; Mamedov, M.D.; Semenov, A.Y.; Golbeck, J.H. Near-IR absorbance changes and electrogenic reactions in the microsecond-to-second time domain in photosystem I. Biophys. J. 1997, 72, 301–315. [Google Scholar] [CrossRef]
- Golbeck, J.; Bryant, D. Photosystem I. In Current Topics in Bioenergetics; Academic Press: New York, NY, USA, 1991; Volume 16, pp. 83–175. [Google Scholar]
- Brettel, K. Electron transfer and arrangement of the redox cofactors in photosystem I. Biochim. Et Biophys. Acta 1997, 1318, 322–373. [Google Scholar] [CrossRef]
- Webber, A.N.; Lubitz, W. P700: The primary electron donor of photosystem I. Biochim. Biophys. Acta 2001, 1507, 61–79. [Google Scholar] [CrossRef]
- Holzwarth, A.R.; Muller, M.G.; Niklas, J.; Lubitz, W. Ultrafast transient absorption studies on photosystem I reaction centers from Chlamydomonas reinhardtii. 2: Mutations near the P700 reaction center chlorophylls provide new insight into the nature of the primary electron donor. Biophys J. 2006, 90, 552–565. [Google Scholar] [CrossRef]
- Zamzam, N.; van Thor, J.J. Excited State Frequencies of Chlorophyll f and Chlorophyll a and Evaluation of Displacement through Franck-Condon Progression Calculations. Molecules 2019, 24, 1326. [Google Scholar] [CrossRef]
- Di Donato, M.; Stahl, A.D.; van Stokkum, I.H.M.; van Grondelle, R.; Groot, M.-L. Cofactors Involved in Light-Driven Charge Separation in Photosystem I Identified by Subpicosecond Infrared Spectroscopy. Biochemistry 2011, 50, 480–490. [Google Scholar] [CrossRef]
- Di Donato, M.; Groot, M.L. Ultrafast infrared spectroscopy in photosynthesis. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2015, 1847, 2–11. [Google Scholar] [CrossRef]
- Cherepanov, D.A.; Shelaev, I.V.; Gostev, F.E.; Nadtochenko, V.A.; Xu, W.; Golbeck, J.H.; Semenov, A.Y. Symmetry breaking in photosystem I: Ultrafast optical studies of variants near the accessory chlorophylls in the A- and B-branches of electron transfer cofactors. Photochem. Photobiol. Sci. 2021, 20, 1209–1227. [Google Scholar] [CrossRef]
- Cherepanov, D.A.; Shelaev, I.V.; Gostev, F.E.; Petrova, A.; Aybush, A.V.; Nadtochenko, V.A.; Xu, W.; Golbeck, J.H.; Semenov, A.Y. Primary charge separation within the structurally symmetric tetrameric Chl2APAPBChl2B chlorophyll exciplex in photosystem I. J. Photochem. Photobiol. B-Biol. 2021, 217, 13. [Google Scholar] [CrossRef]
- Savikhin, S. Ultrafast optical spectroscopy of Photosystem I. In Photosystem I The Light-Driven Plastocyanin: Ferredoxin Oxidoreductase; Golbeck, J., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 155–175. [Google Scholar]
- Croce, R.; van Amerongen, H. Light-harvesting in photosystem I. Photosynth. Res. 2013, 116, 153–166. [Google Scholar] [CrossRef]
- Breton, J.; Nabedryk, E.; Leibl, W. FTIR study of the primary electron donor of photosystem I (P700) revealing delocalization of the charge in P700(+) and localization of the triplet character in (3)P700. Biochemistry 1999, 38, 11585–11592. [Google Scholar] [CrossRef]
- Jordan, P.; Fromme, P.; Witt, H.T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 angstrom resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef]
- Brettel, K. Electron Transfer from Acceptor A1 to The Iron Sulfur Clusters in Photosystem I Measured with A Time Resolution of 2 ns. In Photosynthesis: Mechanisms and Effects; Garab, G., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA, 1998; Volume I, pp. 611–614. [Google Scholar]
- Brettel, K.; Leibl, W. Electron transfer in photosystem I. Biochim. Biophys. Acta Bioenerg. 2001, 1507, 100–114. [Google Scholar] [CrossRef]
- Badshah, S.L.; Sun, J.L.; Mula, S.; Gorka, M.; Baker, P.; Luthra, R.; Lin, S.; van der Est, A.; Golbeck, J.H.; Redding, K.E. Mutations in algal and cyanobacterial Photosystem I that independently affect the yield of initial charge separation in the two electron transfer cofactor branches. Biochim. Biophys. Acta Bioenerg. 2018, 1859, 42–55. [Google Scholar] [CrossRef]
- Santabarbara, S.; Kuprov, I.; Poluektov, O.; Casal, A.; Russell, C.A.; Purton, S.; Evans, M.C.W. Directionality of Electron-Transfer Reactions in Photosystem I of Prokaryotes: Universality of the Bidirectional Electron-Transfer Model. J. Phys. Chem. B 2010, 114, 15158–15171. [Google Scholar] [CrossRef]
- Cherepanov, D.A.; Milanovsky, G.E.; Gopta, O.A.; Balasubramanian, R.; Bryant, D.A.; Semenov, A.Y.; Golbeck, J.H. Electron–Phonon Coupling in Cyanobacterial Photosystem I. J. Phys. Chem. B 2018, 122, 7943–7955. [Google Scholar] [CrossRef]
- Kurashov, V.; Gorka, M.; Milanovsky, G.E.; Johnson, T.W.; Cherepanov, D.A.; Semenov, A.Y.; Golbeck, J.H. Critical evaluation of electron transfer kinetics in P700–FA/FB, P700–FX, and P700–A1 Photosystem I core complexes in liquid and in trehalose glass. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2018, 1859, 1288–1301. [Google Scholar] [CrossRef]
- Guergova-Kuras, M.; Boudreaux, B.; Joliot, A.; Joliot, P.; Redding, K. Evidence for two active branches for electron transfer in photosystem I. Proc. Natl. Acad. Sci. USA 2001, 98, 4437–4442. [Google Scholar] [CrossRef]
- Li, Y.J.; van der Est, A.; Lucas, M.G.; Ramesh, V.M.; Gu, F.F.; Petrenko, A.; Lin, S.; Webber, A.N.; Rappaport, F.; Redding, K. Directing electron transfer within photosystem I by breaking H-bonds in the cofactor branches. Proc. Natl. Acad. Sci. USA 2006, 103, 2144–2149. [Google Scholar] [CrossRef]
- Suga, M.; Ozawa, S.-I.; Yoshida-Motomura, K.; Akita, F.; Miyazaki, N.; Takahashi, Y. Structure of the green algal photosystem I supercomplex with a decameric light-harvesting complex I. Nat. Plants 2019, 5, 626–636. [Google Scholar] [CrossRef]
- Malavath, T.; Caspy, I.; Netzer-El, S.Y.; Klaiman, D.; Nelson, N. Structure and function of wild-type and subunit-depleted photosystem I in Synechocystis. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2018, 1859, 645–654. [Google Scholar] [CrossRef]
- Martin, A. Light Reactions of Photosynthesis: Exploring Early Energy and Electron Transfers in Cyanobacterial Photosystem I via Optical Spectroscopy. Ph.D. Thesis, Purdue University, West Lafayette, India, 2020. [Google Scholar]
- Katz, J.J.; Closs, G.L.; Pennington, F.C.; Thomas, M.R.; Strain, H.H. Infrared Spectra, Molecular Weights, and Molecular Association of Chlorophylls a and b, Methyl Chlorophyllides, and Pheophytins in Various Solvents. J. Am. Chem. Soc. 1963, 85, 3801–3809. [Google Scholar] [CrossRef]
- Breton, J. FTIR Studies of the Primary Electron Donor, P700. In Photosystem I: The Light-Driven Plastocyanin:Ferredoxin Oxidoreductase; Golbeck, J.H., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 271–289. [Google Scholar] [CrossRef]
- Breton, J.; Nabedryk, E.; Clerici, A. Light-induced FTIR difference spectroscopy of photosynthetic charge separation between 9000 and 250 cm−1). Vib. Spectrosc. 1999, 19, 71–75. [Google Scholar] [CrossRef]
- Nabedryk, E.; Leonhard, M.; Maentele, W.; Breton, J. Fourier transform infrared difference spectroscopy shows no evidence for an enolization of chlorophyll a upon cation formation either in vitro or during P700 photooxidation. Biochemistry 1990, 29, 3242–3247. [Google Scholar] [CrossRef]
- Hastings, G.; Ramesh, V.M.; Wang, R.; Sivakumar, V.; Webber, A. Primary donor photo-oxidation in photosystem I: A re-evaluation of (P700+- P700) Fourier transform infrared difference spectra. Biochemistry 2001, 40, 12943–12949. [Google Scholar] [CrossRef]
- Hastings, G.; Badshah, S.L.; Zhao, N.; Redding, K. The A–1 pigment in photosystem I contributes to (P700+–P700) FTIR difference spectra. In Photosynthesis; Hou, H.J.M., Allakhverdiev, S.I., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 67–82. [Google Scholar] [CrossRef]
- Barth, A. The infrared absorption of amino acid side chains. Prog. Biophys. Mol. Biol. 2000, 74, 141–173. [Google Scholar] [CrossRef]
- Susi, H.; Byler, D.M. Resolution-enhanced Fourier transform infrared spectroscopy of enzymes. Methods Enzym. 1986, 130, 290–311. [Google Scholar] [CrossRef]
- Parameswaran, S.; Wang, R.L.; Hastings, G. Calculation of the Vibrational Properties of Chlorophyll a in Solution. J. Phys. Chem. B 2008, 112, 14056–14062. [Google Scholar] [CrossRef]
- Xu, W.; Chitnis, P.; Valieva, A.; van der Est, A.; Pushkar, Y.N.; Krzystyniak, M.; Teutloff, C.; Zech, S.G.; Bittl, R.; Stehlik, D.; et al. Electron Transfer in Cyanobacterial Photosystem I: I. Physiological and Spectroscopic Characterization of Site-Directed Mutants in a Putative Electron Transfer Pathway from A0 Through A1 to FX*. J. Biol. Chem. 2003, 278, 27864–27875. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NB582 C=O | NA600 C=O | |
|---|---|---|
| H2O oxygen atom | 2.2 | 2.2 |
| Mg atom | 4.6 | 4.9 |
| 131-keto C=O oxygen atom of PA | 19.9 | 11.6 |
| 133-ester C=O oxygen atom of PA | 18.5 | 10.4 |
| 131-keto C=O oxygen atom of PB | 11.2 | 20.9 |
| 133-ester C=O oxygen atom of PB | 10.7 | 19.7 |
| 131-keto C=O oxygen atom of A-1A | 9.4 | 15.2 (not in Figure 1) |
| 133-ester C=O oxygen atom of A-1A | 9.3 | 18.7 (not in Figure 1) |
| 131-keto C=O oxygen atom of A-1B | 14.3 (not in Figure 1) | 10.1 |
| 133-ester C=O oxygen atom of A-1B | 17.4 (not in Figure 1) | 8.0 |
| S6803 | ||||||
|---|---|---|---|---|---|---|
| WT | Δ | N-to-M Mutant | Δ | |||
| 131-keto C=O A-1B | 1697(−)/1710(+) | 13 | 1683(−)/1691(+) | 8 | ||
| 131-keto C=O A-1A | 1692(−)/1699(+) | 7 | 1679(−)/1692(+) | 13 | ||
| C. reinhardtii | ||||||
| WT | Δ | N-to-K Mutant | Δ | N-to-D Mutant | Δ | |
| 131-keto C=O A-1B | 1692(−)/1702(+) 1691(−)/1700(+) | 10 9 | 1674(−)/1688(+) | 14 | 1673(−)/1688(+) | 15 |
| 131-keto C=O A-1A | 1698(−)/1706(+) 1697(−)/1704(+) | 8 7 | 1690(−)/1698(+) | 8 | 1670(−)/1682(+) | 12 |
| S6803 | |||||
|---|---|---|---|---|---|
| WT | N-to-M Mutant | Δ | |||
| 131-keto C=O A-1B | 1697(−)/1710(+) | 1683(−)/1691(+) | 14/19 | ||
| 131-keto C=O A–1A | 1692(−)/1699(+) | 1679(−)/1692(+) | 13/7 | ||
| C. reinhardtii | |||||
| WT | N-to-K Mutant | Δ | N-to-D Mutant | Δ | |
| 131-keto C=O A-1B | 1692(−)/1702(+) 1691(−)/1700(+) | 1674(−)/1688(+) | 18/14 17/12 | 1673(−)/1688(+) | 19/14 18/12 |
| 131-keto C=O A-1A | 1698(−)/1706(+) 1697(−)/1704(+) | 1690(−)/1698(+) | 8/8 7/6 | 1670(−)/1682(+) | 28/24 17/22 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirpich, J.S.; Luo, L.; Nelson, M.R.; Agarwala, N.; Xu, W.; Hastings, G. Is the A-1 Pigment in Photosystem I Part of P700? A (P700+–P700) FTIR Difference Spectroscopy Study of A-1 Mutants. Int. J. Mol. Sci. 2024, 25, 4839. https://doi.org/10.3390/ijms25094839
Kirpich JS, Luo L, Nelson MR, Agarwala N, Xu W, Hastings G. Is the A-1 Pigment in Photosystem I Part of P700? A (P700+–P700) FTIR Difference Spectroscopy Study of A-1 Mutants. International Journal of Molecular Sciences. 2024; 25(9):4839. https://doi.org/10.3390/ijms25094839
Chicago/Turabian StyleKirpich, Julia S., Lujun Luo, Michael R. Nelson, Neva Agarwala, Wu Xu, and Gary Hastings. 2024. "Is the A-1 Pigment in Photosystem I Part of P700? A (P700+–P700) FTIR Difference Spectroscopy Study of A-1 Mutants" International Journal of Molecular Sciences 25, no. 9: 4839. https://doi.org/10.3390/ijms25094839