
Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides


Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Synthesis α-L-Fucose Fluoresceinated Glycopolymer
2.3. Glycopolymer Specificity Verification
2.4. Bacteria
2.4.1. Clinical Isolates and Laboratory Strains Sources, Characteristics, and Cultures
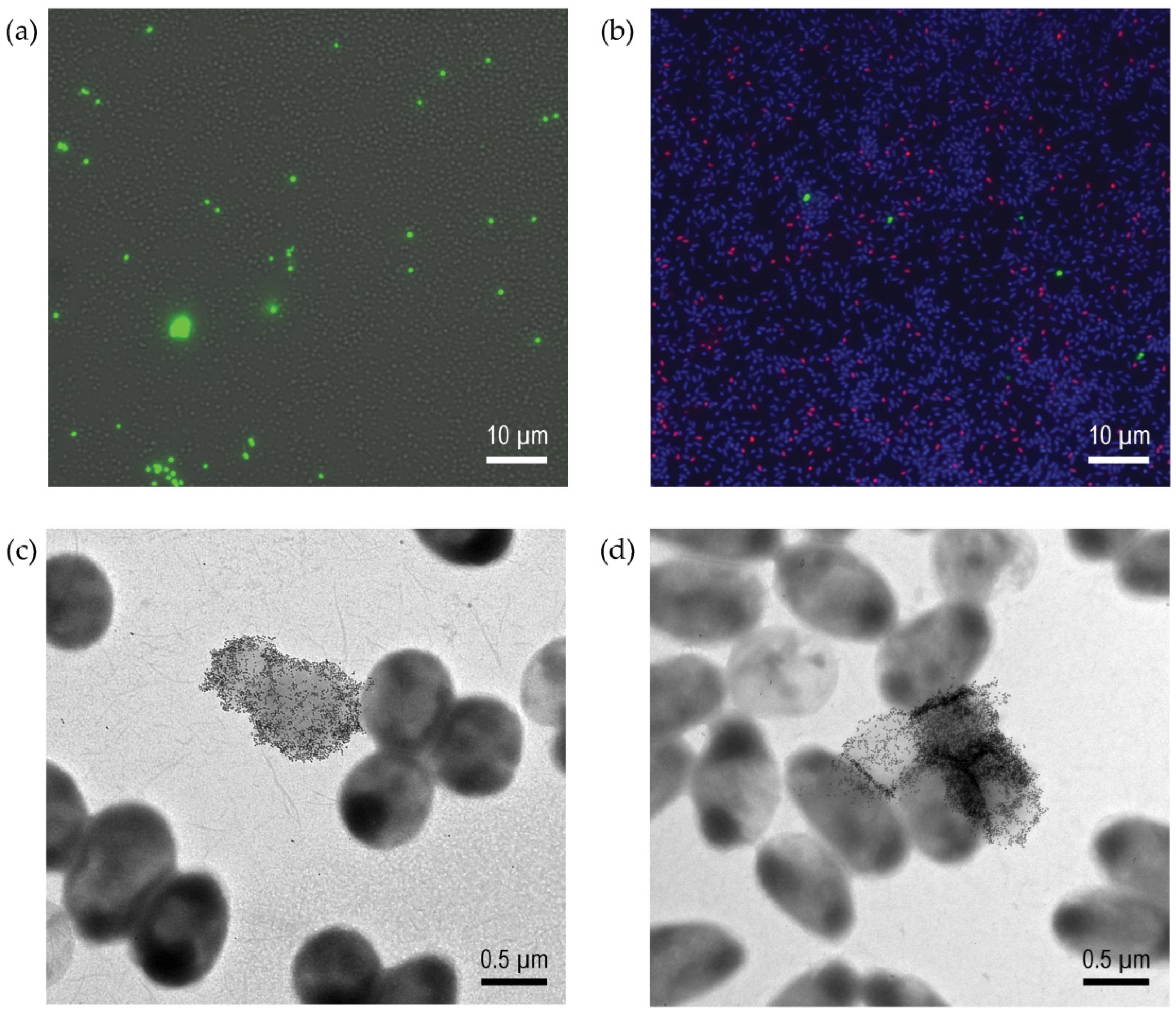
2.4.2. P. aeruginosa Structural Features Assessment by Transmission Electron Microscopy
2.4.3. Preparation of Bacteria for Binding Experiments
2.5. P. aeruginosa Carbohydrate-Binding Assays Using Fluorescent Glycopolymers
2.5.1. Solution-Phase Binding Reactions
2.5.2. Detection of Bacterial-Glycopolymer Binding
2.6. Localization and Population Distribution of Bacterial-Bound Glycopolymer with TEM and Flow Cytometry
2.7. Isolation and Identification of Soluble Lectins LecA and LecB from P. aeruginosa
2.7.1. Preparations of Presumptive LecA and LecB Fractions from P. aeruginosa Cultures
2.7.2. Identification of LecA and LecB Proteins with Mass Spectrometry
3. Results
3.1. P. aeruginosa Collection Heterogeneity in Source, Physiology, and Phenotypes
3.1.1. P. aeruginosa Collection Character Overview
3.1.2. Physiological Diversity in This Collection of CF and Non-CF P. aeruginosa
3.1.3. Phenotypic Diversity of This P. aeruginosa Collection
3.2. PAA-Fluor Glycopolymers Were Amenable to Solution-Phase Bacterial Binding Studies
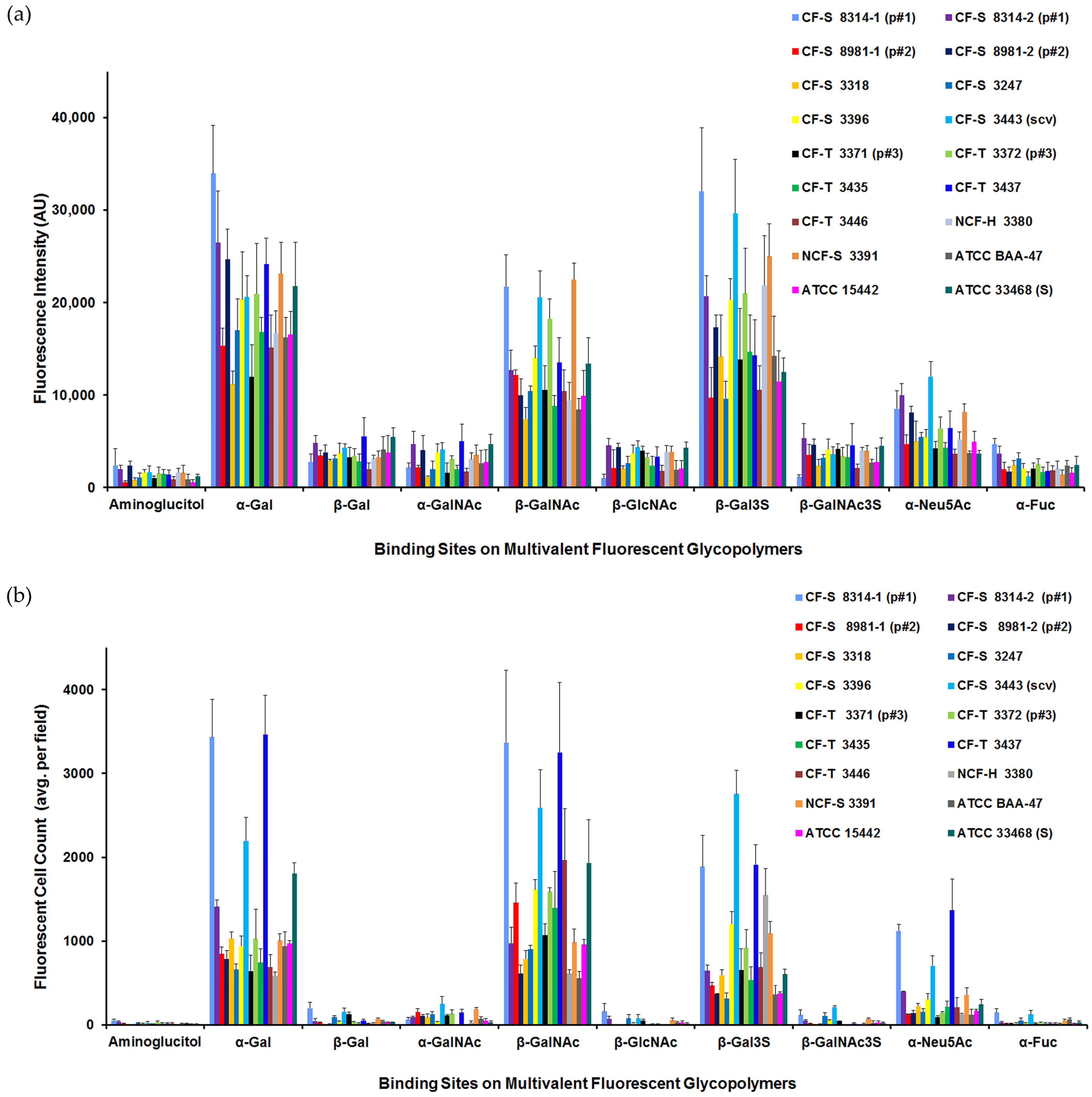
3.3. P. aeruginosa Carbohydrate-Binding Profiles
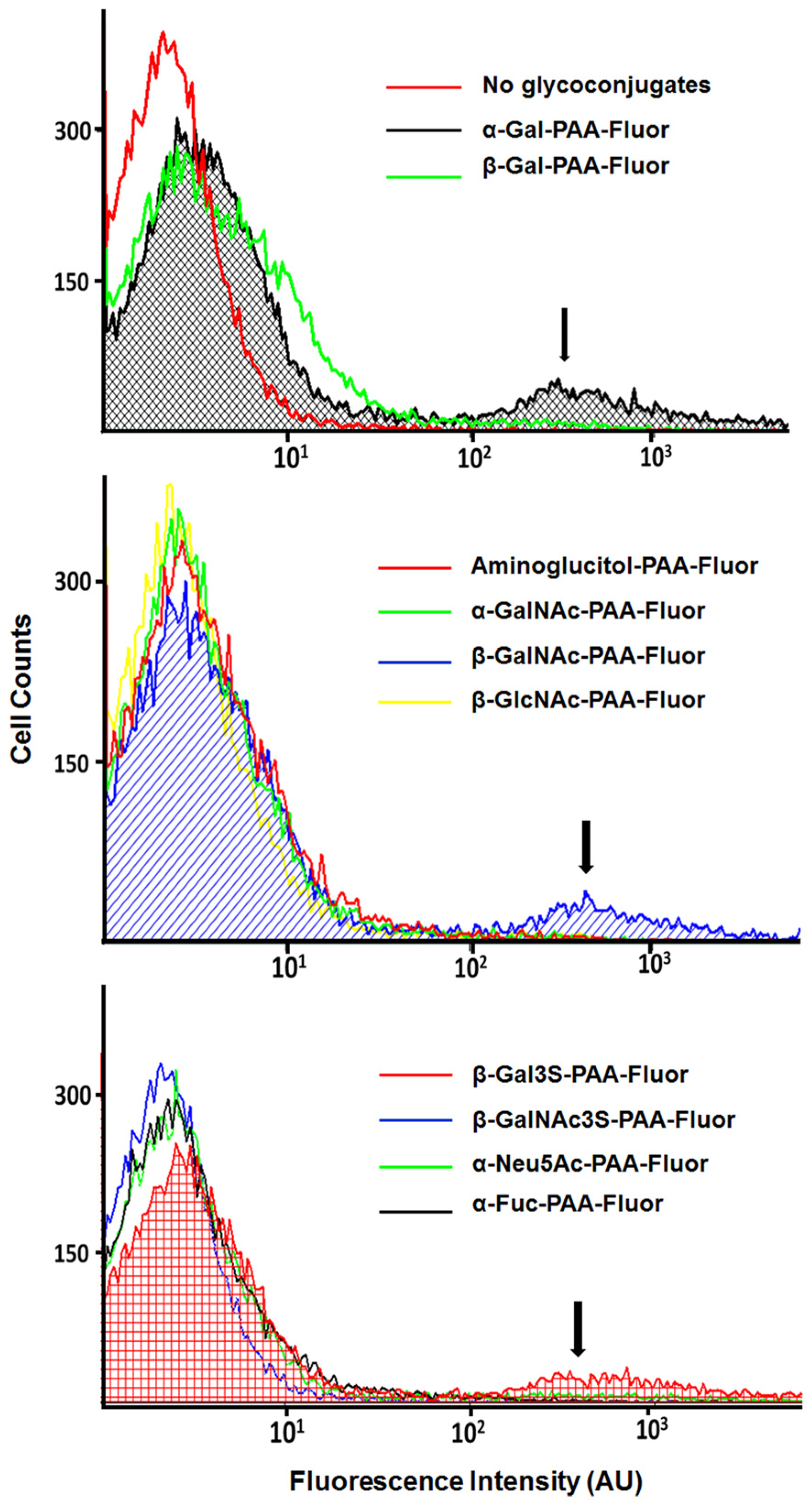
3.4. Fluorescence Microscopy, TEM, and Flow Cytometry Reveal Subset of Population Was Responsible for the Positive Binding Characteristic
3.4.1. Glycopolymer-Bound Populations Investigated via Microscopy
3.4.2. Glycopolymer-Bound Populations Evaluated by Flow Cytometry
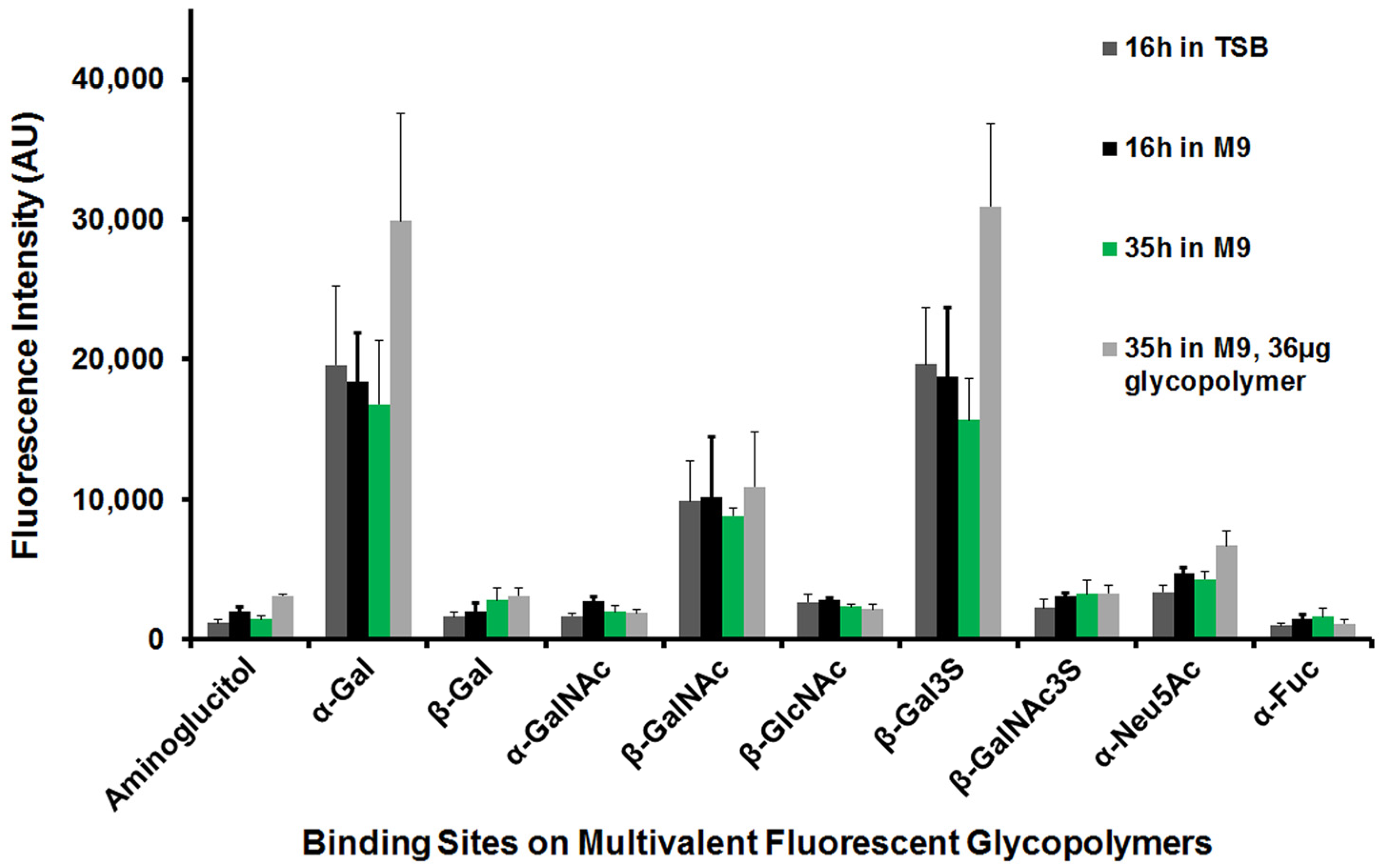
3.5. Varied Assay Conditions Did Not Change the P. aeruginosa Preferred Pendant Monosaccharide PAA-Fluor Glycopolymer-Binding Profile
3.6. Lectins LecA (PA-IL) and LecB (PA-IIL) Were Present in Binding Assay Cell Preparations of CF Sputum-Derived P. aeruginosa
3.7. P. aeruginosa Source, Phenotype, and Structural Features Did Not Clearly Predict Enhanced Glycopolymer-Binding Profiles
3.7.1. Overview of P. aeruginosa “Higher Binding” Glycopolymers Status
3.7.2. Considerations of Source, Phenotype, and Structural Features of P. aeruginosa for Contribution to Glycopolymer “High Binding” Status
3.7.3. Summary of P. aeruginosa Attributes and Enhanced Carbohydrate-Binding Profiles
4. Discussion
4.1. Importance of Investigating the Characteristics Diversity of P. aeruginosa in Collection for Representative Data to Inform Clinically Relevant Carbohydrate-Binding-Based Therapeutic Strategies
4.2. Additional Factors Potentially Influencing P. aeruginosa Carbohydrate Binding In Vivo
4.3. Fluorescent Polyacrylamide-Based Glycopolymers with Pendant Monosaccharides as Convenient Tools for Surveying Whole-Cell P. aeruginosa Carbohydrate Binding
4.4. Insights Gained from Imaging and Quantitation of P. aeruginosa Carbohydrate-Binding Profiles via Multiple Modalities
4.5. Implications of Carbohydrate-Binding Profile Observations across This P. aeruginosa Collection on Potential Carbohydrate-Based Therapeutics Development and Application to Cystic Fibrosis Patients
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malhotra, S.; Hayes, D., Jr.; Wozniak, D.J. Cystic fibrosis and Pseudomonas aeruginosa: The host-microbe interface. Clin. Microbiol. Rev. 2019, 32, e00138-18. [Google Scholar] [CrossRef] [PubMed]
- Morrison, C.B.; Markovetz, M.R.; Ehre, C. Mucus, mucins, and cystic fibrosis. Pediatr. Pulmonol. 2019, 54, S84–S96. [Google Scholar] [CrossRef]
- Nichols, D.P.; Durmowicz, A.G.; Field, A.; Flume, P.A.; VanDevanter, D.R.; Mayer-Hamblett, N. Developing inhaled antibiotics in cystic fibrosis: Current challenges and opportunities. Ann. Am. Thorac. Soc. 2019, 16, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Somayaji, R.; Nichols, D.P.; Bell, S.C. Cystic fibrosis—Ten promising therapeutic approaches in the current era of care. Expert. Opin. Investig. Drugs 2020, 29, 1107–1124. [Google Scholar] [CrossRef] [PubMed]
- Girón Moreno, R.M.; García-Clemente, M.; Diab-Cáceres, L.; Martínez-Vergara, A.; Martínez-García, M.Á.; Gómez-Punter, R.M. Treatment of pulmonary disease of cystic fibrosis: A comprehensive review. Antibiotics 2021, 10, 486. [Google Scholar] [CrossRef] [PubMed]
- Manos, J. Current and emerging therapies to combat cystic fibrosis lung infections. Microorganisms 2021, 9, 1874. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.; Waters, V.; Grasemann, H. Approaches to targeting bacterial biofilms in cystic fibrosis airways. Int. J. Mol. Sci. 2021, 22, 2155. [Google Scholar] [CrossRef]
- Reece, E.; Bettio, P.H.A.; Renwick, J. Polymicrobial interactions in the cystic fibrosis airway microbiome impact the antimicrobial susceptibility of Pseudomonas aeruginosa. Antibiotics 2021, 10, 827. [Google Scholar] [CrossRef]
- Venkatakrishnan, V.; Packer, N.H.; Thaysen-Andersen, M. Host mucin glycosylation plays a role in bacterial adhesion in lungs of individuals with cystic fibrosis. Expert. Rev. Respir. Med. 2013, 7, 553–576. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Martin, I.; Sainz-Mejias, M.; McClean, S. Pseudomonas aeruginosa: An audacious pathogen with an adaptable arsenal of virulence factors. Int. J. Mol. Sci 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.; Fothergill, J.L. The role of multispecies social interactions in shaping Pseudomonas aeruginosa pathogenicity in the cystic fibrosis lung. FEMS Microbiol. Lett. 2017, 364, fnx128. [Google Scholar] [CrossRef]
- Wagner, S.; Sommer, R.; Hinsberger, S.; Lu, C.; Hartmann, R.W.; Empting, M.; Titz, A. Novel strategies for the treatment of Pseudomonas aeruginosa infections. J. Med. Chem. 2016, 59, 5929–5969. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Singh, P.K. Evolving stealth: Genetic adaptation of Pseudomonas aeruginosa during cystic fibrosis infections. Proc. Natl. Acad. Sci. USA 2006, 103, 8305–8306. [Google Scholar] [CrossRef] [PubMed]
- Starkey, M.; Hickman, J.H.; Ma, L.; Zhang, N.; De, L.S.; Hinz, A.; Palacios, S.; Manoil, C.; Kirisits, M.J.; Starner, T.D.; et al. Pseudomonas aeruginosa rugose small-colony variants have adaptations that likely promote persistence in the cystic fibrosis lung. J. Bacteriol. 2009, 191, 3492–3503. [Google Scholar] [CrossRef] [PubMed]
- Folkesson, A.; Jelsbak, L.; Yang, L.; Johansen, H.K.; Ciofu, O.; Hoiby, N.; Molin, S. Adaptation of Pseudomonas aeruginosa to the cystic fibrosis airway: An evolutionary perspective. Nat. Rev. Microbiol. 2012, 10, 841–851. [Google Scholar] [CrossRef] [PubMed]
- Workentine, M.L.; Sibley, C.D.; Glezerson, B.; Purighalla, S.; Norgaard-Gron, J.C.; Parkins, M.D.; Rabin, H.R.; Surette, M.G. Phenotypic heterogeneity of Pseudomonas aeruginosa populations in a cystic fibrosis patient. PLoS ONE 2013, 8, e60225. [Google Scholar] [CrossRef]
- Jorth, P.; Staudinger, B.J.; Wu, X.; Hisert, K.B.; Hayden, H.; Garudathri, J.; Harding, C.L.; Radey, M.C.; Rezayat, A.; Bautista, G.; et al. Regional isolation drives bacterial diversification within cystic fibrosis lungs. Cell Host Microbe 2015, 18, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Malone, J.G. Role of small colony variants in persistence of Pseudomonas aeruginosa infections in cystic fibrosis lungs. Infect. Drug Resist. 2015, 8, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa Evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; La, R.R.; Bartell, J.A.; Marvig, R.L.; Haagensen, J.A.J.; Sommer, L.M.; Molin, S.; Johansen, H.K. Pseudomonas aeruginosa adaptation and evolution in patients with cystic fibrosis. Nat. Rev. Microbiol. 2021, 19, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Tolker-Nielsen, T. Tolerance and resistance of Pseudomonas aeruginosa biofilms to antimicrobial agents-how P. aeruginosa can escape antibiotics. Front. Microbiol. 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed]
- Limqueco, E.; Passos Da, S.D.; Reichhardt, C.; Su, F.Y.; Das, D.; Chen, J.; Srinivasan, S.; Convertine, A.; Skerrett, S.J.; Parsek, M.R.; et al. Mannose conjugated polymer targeting P. aeruginosa biofilms. ACS Infect. Dis. 2020, 6, 2866–2871. [Google Scholar] [CrossRef] [PubMed]
- Trostrup, H.; Laulund, A.S.; Moser, C. Insights into host-pathogen interactions in biofilm-infected wounds reveal possibilities for new treatment strategies. Antibiotics 2020, 9, 396. [Google Scholar] [CrossRef] [PubMed]
- Jennings, L.K.; Dreifus, J.E.; Reichhardt, C.; Storek, K.M.; Secor, P.R.; Wozniak, D.J.; Hisert, K.B.; Parsek, M.R. Pseudomonas aeruginosa aggregates in cystic fibrosis sputum produce exopolysaccharides that likely impede current therapies. Cell Rep. 2021, 34, 108782. [Google Scholar] [CrossRef] [PubMed]
- Metelkina, O.; Huck, B.; O’Connor, J.S.; Koch, M.; Manz, A.; Lehr, C.M.; Titz, A. Targeting extracellular lectins of Pseudomonas aeruginosa with glycomimetic liposomes. J. Mater. Chem. B 2022, 10, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Tuon, F.F.; Dantas, L.R.; Suss, P.H.; Tasca Ribeiro, V.S. Pathogenesis of the Pseudomonas aeruginosa Biofilm: A Review. Pathogens 2022, 11, 300. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Eisha, S.; Park, S.; Morris, A.J.; Martin, I. How three self-secreted biofilm exopolysaccharides of Pseudomonas aeruginosa, Psl, Pel, and alginate, can each be exploited for antibiotic adjuvant effects in cystic fibrosis lung infection. Int. J. Mol. Sci 2023, 24, 8709. [Google Scholar] [CrossRef] [PubMed]
- Mayer-Hamblett, N.; Rosenfeld, M.; Gibson, R.L.; Ramsey, B.W.; Kulasekara, H.D.; Retsch-Bogart, G.Z.; Morgan, W.; Wolter, D.J.; Pope, C.E.; Houston, L.S.; et al. Pseudomonas aeruginosa in vitro phenotypes distinguish cystic fibrosis infection stages and outcomes. Am. J. Respir. Crit Care Med. 2014, 190, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Kamath, K.S.; Pascovici, D.; Penesyan, A.; Goel, A.; Venkatakrishnan, V.; Paulsen, I.T.; Packer, N.H.; Molloy, M.P. Pseudomonas aeruginosa cell membrane protein expression from phenotypically diverse cystic fibrosis isolates demonstrates host-specific adaptations. J. Proteome Res. 2016, 15, 2152–2163. [Google Scholar] [CrossRef] [PubMed]
- Bacci, G.; Mengoni, A.; Fiscarelli, E.; Segata, N.; Taccetti, G.; Dolce, D.; Paganin, P.; Morelli, P.; Tuccio, V.; De Alessandri, A.; et al. A different microbiome gene repertoire in the airways of cystic fibrosis patients with severe lung disease. Int. J. Mol. Sci. 2017, 18, 1654. [Google Scholar] [CrossRef]
- Fernandez-Barat, L.; Ciofu, O.; Kragh, K.N.; Pressler, T.; Johansen, U.; Motos, A.; Torres, A.; Hoiby, N. Phenotypic shift in Pseudomonas aeruginosa populations from cystic fibrosis lungs after 2-week anti-Pseudomonal treatment. J. Cyst. Fibros. 2017, 16, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.K.; Rau, M.H.; Johansen, H.K.; Ciofu, O.; Jelsbak, L.; Yang, L.; Folkesson, A.; Jarmer, H.O.; Aanaes, K.; von Buchwald, C.; et al. Evolution and diversification of Pseudomonas aeruginosa in the paranasal sinuses of cystic fibrosis children have implications for chronic lung infection. ISME J. 2012, 6, 31–45. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.T.; Diaz Caballero, J.; Cheang, M.; Coburn, B.; Wang, P.W.; Donaldson, S.L.; Zhang, Y.; Liu, M.; Keshavjee, S.; Yau, Y.C.W.; et al. Phenotypic diversity within a Pseudomonas aeruginosa population infecting an adult with cystic fibrosis. Sci. Rep. 2015, 5, 10932. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, J.; Cramer, N.; Fischer, S.; Wiehlmann, L.; Tummler, B. Long-term microevolution of Pseudomonas aeruginosa differs between mildly and severely affected cystic fibrosis lungs. Am. J. Respir. Cell Mol. Biol. 2018, 59, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Fleming, D.; Rumbaugh, K.P. Approaches to dispersing medical biofilms. Microorganisms 2017, 5, 15. [Google Scholar] [CrossRef]
- Curran, C.S.; Bolig, T.; Torabi-Parizi, P. Mechanisms and targeted therapies for Pseudomonas aeruginosa lung infection. Am. J. Respir. Crit Care Med. 2018, 197, 708–727. [Google Scholar] [CrossRef] [PubMed]
- Loo, C.Y.; Lee, W.H.; Lauretani, G.; Scalia, S.; Cipolla, D.; Traini, D.; Young, P.; Ong, H.X. Sweetening inhaled antibiotic treatment for eradication of chronic respiratory biofilm infection. Pharm. Res. 2018, 35, 50. [Google Scholar] [CrossRef] [PubMed]
- Rezzoagli, C.; Archetti, M.; Mignot, I.; Baumgartner, M.; Kummerli, R. Combining antibiotics with antivirulence compounds can have synergistic effects and reverse selection for antibiotic resistance in Pseudomonas aeruginosa. PLoS Biol. 2020, 18, e3000805. [Google Scholar] [CrossRef] [PubMed]
- Cystic Fibrosis Foundation Website, Drug Development Pipeline—Clinical Trials Tool. Available online: https://apps.cff.org/trials/pipeline (accessed on 19 February 2024).
- Choi, V.; Rohn, J.L.; Stoodley, P.; Carugo, D.; Stride, E. Drug delivery strategies for antibiofilm therapy. Nat. Rev. Microbiol. 2023, 21, 555–572. [Google Scholar] [CrossRef] [PubMed]
- Mowat, E.; Paterson, S.; Fothergill, J.L.; Wright, E.A.; Ledson, M.J.; Walshaw, M.J.; Brockhurst, M.A.; Winstanley, C. Pseudomonas aeruginosa population diversity and turnover in cystic fibrosis chronic infections. Am. J. Respir. Crit Care Med. 2011, 183, 1674–1679. [Google Scholar] [CrossRef]
- Jiang, Y.; Geng, M.; Bai, L. Targeting biofilms therapy: Current research strategies and development hurdles. Microorganisms 2020, 8, 1222. [Google Scholar] [CrossRef] [PubMed]
- Chance, D.L.; Mawhinney, T.P. Observations of, and insights into, cystic fibrosis mucus heterogeneity in the pre-modulator era: Sputum characteristics, DNA and glycoprotein content, and solubilization time. J. Respir. 2021, 1, 8–29. [Google Scholar] [CrossRef]
- Patil, P.D.; Zheng, H.; Burns, F.N.; Ibanez, A.C.S.; Jin, Y.; Luk, Y.Y. Chimeric ligands of pili and lectin a inhibit tolerance, persistence, and virulence factors of Pseudomonas aeruginosa over a wide range of phenotypes. ACS Infect. Dis. 2022, 8, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Rouillard, K.R.; Markovetz, M.R.; Kissner, W.J.; Boone, W.L.; Plott, L.M.; Hill, D.B. Altering the viscoelastic properties of mucus-grown Pseudomonas aeruginosa biofilms affects antibiotic susceptibility. Biofilm 2023, 5, 100104. [Google Scholar] [CrossRef]
- Bucior, I.; Abbott, J.; Song, Y.; Matthay, M.A.; Engel, J.N. Sugar administration is an effective adjunctive therapy in the treatment of Pseudomonas aeruginosa pneumonia. Am. J. Physiol. Lung Cell Mol. Physiol. 2013, 305, L352–L363. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Krivozubov, M.S.; Karyagina, A.S.; Gintsburg, A.L. Pseudomonas aeruginosa lectins as targets for novel antibacterials. Acta Nat. 2015, 7, 29–41. [Google Scholar] [CrossRef]
- Taccetti, G.; Francalanci, M.; Pizzamiglio, G.; Messore, B.; Carnovale, V.; Cimino, G.; Cipolli, M. Cystic fibrosis: Recent insights into inhaled antibiotic treatment and future perspectives. Antibiotics 2021, 10, 338. [Google Scholar] [CrossRef] [PubMed]
- Mawhinney, T.P.; Chance, D.L.; Waters, J.K.; Mossine, V.V.; He, S.; Cassity, N.A. Characterization of blood group antigen-containing oligosaccharides isolated from human respiratory mucous glycoproteins. In Recent Developments in Carbohydrate Research; Pandalai, S.G., Ed.; Transworld Research Network: Kerala, India, 2003; Volume 1, pp. 1–25. [Google Scholar]
- Ehre, C.; Rushton, Z.L.; Wang, B.; Hothem, L.N.; Morrison, C.B.; Fontana, N.C.; Markovetz, M.R.; Delion, M.F.; Kato, T.; Villalon, D.; et al. An improved inhaled mucolytic to treat airway muco-obstructive diseases. Am. J. Respir. Crit Care Med. 2019, 199, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Flume, P.A.; Amelina, E.; Daines, C.L.; Charlton, B.; Leadbetter, J.; Guasconi, A.; Aitken, M.L. Efficacy and safety of inhaled dry-powder mannitol in adults with cystic fibrosis: An international, randomized controlled study. J. Cyst. Fibros. 2021, 20, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Michaud, G.; Visini, R.; Bergmann, M.; Salerno, G.; Bosco, R.; Gillon, E.; Richichi, B.; Nativi, C.; Imberty, A.; Stocker, A. Overcoming antibiotic resistance in Pseudomonas aeruginosa biofilms using glycopeptide dendrimers. Chem. Sci. 2016, 7, 166–182. [Google Scholar] [CrossRef] [PubMed]
- Musken, M.; Pawar, V.; Schwebs, T.; Bahre, H.; Felgner, S.; Weiss, S.; Haussler, S. Breaking the vicious cycle of antibiotic killing and regrowth of biofilm-residing Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e01635-18. [Google Scholar] [CrossRef] [PubMed]
- Boffoli, D.; Bellato, F.; Avancini, G.; Gurnani, P.; Yilmaz, G.; Romero, M.; Robertson, S.; Moret, F.; Sandrelli, F.; Caliceti, P.; et al. Tobramycin-loaded complexes to prevent and disrupt Pseudomonas aeruginosa biofilms. Drug Deliv. Transl. Res. 2021, 12, 1788–1810. [Google Scholar] [CrossRef] [PubMed]
- Hauck, D.; Joachim, I.; Frommeyer, B.; Varrot, A.; Philipp, B.; Moller, H.M.; Imberty, A.; Exner, T.E.; Titz, A. Discovery of two classes of potent glycomimetic inhibitors of Pseudomonas aeruginosa LecB with distinct binding modes. ACS Chem. Biol. 2013, 8, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Maura, D.; Ballok, A.E.; Rahme, L.G. Considerations and caveats in anti-virulence drug development. Curr. Opin. Microbiol. 2016, 33, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Zahorska, E.; Rosato, F.; Stober, K.; Kuhaudomlarp, S.; Meiers, J.; Hauck, D.; Reith, D.; Gillon, E.; Rox, K.; Imberty, A.; et al. Neutralizing the impact of the virulence factor LecA from Pseudomonas aeruginosa on human cells with new glycomimetic inhibitors. Angew. Chem. Int. Ed. Engl. 2023, 62, e202215535. [Google Scholar] [CrossRef] [PubMed]
- Winzer, K.; Falconer, C.; Garber, N.C.; Diggle, S.P.; Camara, M.; Williams, P. The Pseudomonas aeruginosa lectins PA-IL and PA-IIL are controlled by quorum sensing and by RpoS. J. Bacteriol. 2000, 182, 6401–6411. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Kulkarni, S.S. Small carbohydrate derivatives as potent antibiofilm agents. J. Med. Chem. 2022, 65, 8525–8549. [Google Scholar] [CrossRef]
- Imberty, A.; Wimmerova, M.; Mitchell, E.P.; Gilboa-Garber, N. Structures of the lectins from Pseudomonas aeruginosa: Insights into the molecular basis for host glycan recognition. Microbes Infect. 2004, 6, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Sommer, R.; Rox, K.; Wagner, S.; Hauck, D.; Henrikus, S.S.; Newsad, S.; Arnold, T.; Ryckmans, T.; Bronstrup, M.; Imberty, A.; et al. Anti-biofilm agents against Pseudomonas aeruginosa: A structure-activity relationship study of C-glycosidic LecB inhibitors. J. Med. Chem. 2019, 62, 9201–9216. [Google Scholar] [CrossRef] [PubMed]
- Meiers, J.; Zahorska, E.; Rohrig, T.; Hauck, D.; Wagner, S.; Titz, A. Directing drugs to bugs: Antibiotic-carbohydrate conjugates targeting biofilm-associated lectins of Pseudomonas aeruginosa. J. Med. Chem. 2020, 63, 11707–11724. [Google Scholar] [CrossRef] [PubMed]
- Meiers, J.; Rox, K.; Titz, A. Lectin-targeted prodrugs activated by Pseudomonas aeruginosa for self-destructive antibiotic release. J. Med. Chem. 2022, 65, 13988–14014. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, C.; Eichwald, C.; Eberl, L. Multicellularity in bacteria: From division of labor to biofilm formation. In Evolutionary Transitions to Multicellular Life; Springer: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Scharfman, A.; Degroote, S.; Beau, J.; Lamblin, G.; Roussel, P.; Mazurier, J. Pseudomonas aeruginosa binds to neoglycoconjugates bearing mucin carbohydrate determinants and predominantly to sialyl-Lewis x conjugates. Glycobiology 1999, 9, 757–764. [Google Scholar] [CrossRef] [PubMed]
- Ramphal, R.; Arora, S.K. Recognition of mucin components by Pseudomonas aeruginosa. Glycoconj. J. 2001, 18, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Jeffries, J.L.; Jia, J.; Choi, W.; Choe, S.; Miao, J.; Xu, Y.; Powell, R.; Lin, J.; Kuang, Z.; Gaskins, H.R.; et al. Pseudomonas aeruginosa pyocyanin modulates mucin glycosylation with sialyl-Lewis(x) to increase binding to airway epithelial cells. Mucosal. Immunol. 2016, 9, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Bucior, I.; Pielage, J.F.; Engel, J.N. Pseudomonas aeruginosa pili and flagella mediate distinct binding and signaling events at the apical and basolateral surface of airway epithelium. PLoS Pathog. 2012, 8, e1002616. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, E.P.; Kim, B.T.; Kim, K.C. Identification of Pseudomonas aeruginosa flagellin as an adhesin for Muc1 mucin. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 282, L751–L756. [Google Scholar] [CrossRef]
- Chemani, C.; Imberty, A.; de Bentzmann, S.; Pierre, M.; Wimmerova, M.; Guery, B.P.; Faure, K. Role of LecA and LecB lectins in Pseudomonas aeruginosa-induced lung injury and effect of carbohydrate ligands. Infect. Immun. 2009, 77, 2065–2075. [Google Scholar] [CrossRef] [PubMed]
- Sheth, H.B.; Lee, K.K.; Wong, W.Y.; Srivastava, G.; Hindsgaul, O.; Hodges, R.S.; Paranchych, W.; Irvin, R.T. The pili of Pseudomonas aeruginosa strains PAK and PAO bind specifically to the carbohydrate sequence βGalNAc(1-4)βGal found in glycosphingolipids asialo-GM1 and asialo-GM2. Mol. Microbiol. 1994, 11, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Ramphal, R.; Carnoy, C.; Fievre, S.; Michalski, J.C.; Houdret, N.; Lamblin, G.; Strecker, G.; Roussel, P. Pseudomonas aeruginosa recognizes carbohydrate chains containing type 1 (Gal β1-3GlcNAc) or type 2 (Gal β1-4GlcNAc) disaccharide units. Infect. Immun. 1991, 59, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Hahn, H.P. The type-4 pilus is the major virulence-associated adhesin of Pseudomonas aeruginosa—A review. Gene 1997, 192, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Lamblin, G.; Degroote, S.; Perini, J.M.; Delmotte, P.; Scharfman, A.; Davril, M.; Lo-Guidice, J.M.; Houdret, N.; Dumur, V.; Klein, A.; et al. Human airway mucin glycosylation: A combinatory of carbohydrate determinants which vary in cystic fibrosis. Glycoconj. J. 2001, 18, 661–684. [Google Scholar] [CrossRef] [PubMed]
- Scharfman, A.; Arora, S.K.; Delmotte, P.; Van, B.E.; Mazurier, J.; Ramphal, R.; Roussel, P. Recognition of Lewis x derivatives present on mucins by flagellar components of Pseudomonas aeruginosa. Infect. Immun. 2001, 69, 5243–5248. [Google Scholar] [CrossRef] [PubMed]
- Mewe, M.; Tielker, D.; Schonberg, R.; Schachner, M.; Jaeger, K.E.; Schumacher, U. Pseudomonas aeruginosa lectins I and II and their interaction with human airway cilia. J. Laryngol. Otol. 2005, 119, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Krivan, H.C.; Roberts, D.D.; Ginsburg, V. Many pulmonary pathogenic bacteria bind specifically to the carbohydrate sequence GalNAc beta 1-4Gal found in some glycolipids. Proc. Natl. Acad. Sci. USA 1988, 85, 6157–6161. [Google Scholar] [CrossRef] [PubMed]
- Imundo, L.; Barasch, J.; Prince, A.; Al-Awqati, Q. Cystic fibrosis epithelial cells have a receptor for pathogenic bacteria on their apical surface. Proc. Natl. Acad. Sci. USA 1995, 92, 3019–3023. [Google Scholar] [CrossRef] [PubMed]
- Saiman, L.; Prince, A. Pseudomonas aeruginosa pili bind to asialoGM1 which is increased on the surface of cystic fibrosis epithelial cells. J. Clin. Investig. 1993, 92, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.K.; Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. The Pseudomonas aeruginosa flagellar cap protein, FliD, is responsible for mucin adhesion. Infect. Immun. 1998, 66, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Scharfman, A.; Delmotte, P.; Beau, J.; Lamblin, G.; Roussel, P.; Mazurier, J. Sialyl-Le(x) and sulfo-sialyl-Le(x) determinants are receptors for P. aeruginosa. Glycoconj. J. 2000, 17, 735–740. [Google Scholar] [CrossRef] [PubMed]
- Davril, M.; Degroote, S.; Humbert, P.; Galabert, C.; Dumur, V.; Lafitte, J.J.; Lamblin, G.; Roussel, P. The sialylation of bronchial mucins secreted by patients suffering from cystic fibrosis or from chronic bronchitis is related to the severity of airway infection. Glycobiology 1999, 9, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Degroote, S.; Maes, E.; Humbert, P.; Delmotte, P.; Lamblin, G.; Roussel, P. Sulfated oligosaccharides isolated from the respiratory mucins of a secretor patient suffering from chronic bronchitis. Biochimie 2003, 85, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Royall, J.A.; Damera, G.; Sachdev, G.P.; Cummings, R.D. Altered O-glycosylation and sulfation of airway mucins associated with cystic fibrosis. Glycobiology 2005, 15, 747–775. [Google Scholar] [CrossRef] [PubMed]
- Zanin, M.; Baviskar, P.; Webster, R.; Webby, R. The interaction between respiratory pathogens and mucus. Cell Host Microbe 2016, 19, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Adam, E.C.; Mitchell, B.S.; Schumacher, D.U.; Grant, G.; Schumacher, U. Pseudomonas aeruginosa II lectin stops human ciliary beating: Therapeutic implications of fucose. Am. J. Respir. Crit Care Med. 1997, 155, 2102–2104. [Google Scholar] [CrossRef] [PubMed]
- Adam, E.C.; Schumacher, D.U.; Schumacher, U. Cilia from a cystic fibrosis patient react to the ciliotoxic Pseudomonas aeruginosa II lectin in a similar manner to normal control cilia—A case report. J. Laryngol. Otol. 1997, 111, 760–762. [Google Scholar] [CrossRef] [PubMed]
- Garber, N.C.; Zinger-Yosovich, K.D.; Sudakevitz, D.; Axelrad, I.; Gilboa-Garber, N. Regulation of lectin production by the human pathogens Pseudomonas aeruginosa and Chromobacterium violaceum: Effects of choline, trehalose, and ethanol. Adv. Exp. Med. Biol. 2011, 705, 229–256. [Google Scholar] [CrossRef] [PubMed]
- Passos da Silva, D.; Matwichuk, M.L.; Townsend, D.O.; Reichhardt, C.; Lamba, D.; Wozniak, D.J.; Parsek, M.R. The Pseudomonas aeruginosa lectin LecB binds to the exopolysaccharide Psl and stabilizes the biofilm matrix. Nat. Commun. 2019, 10, 2183. [Google Scholar] [CrossRef] [PubMed]
- Tielker, D.; Hacker, S.; Loris, R.; Strathmann, M.; Wingender, J.; Wilhelm, S.; Rosenau, F.; Jaeger, K.E. Pseudomonas aeruginosa lectin LecB is located in the outer membrane and is involved in biofilm formation. Microbiology 2005, 151 Pt 5, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.M.; Wu, J.H.; Singh, T.; Liu, J.H.; Tsai, M.S.; Gilboa-Garber, N. Interactions of the fucose-specific Pseudomonas aeruginosa lectin, PA-IIL, with mammalian glycoconjugates bearing polyvalent Lewis(a) and ABH blood group glycotopes. Biochimie 2006, 88, 1479–1492. [Google Scholar] [CrossRef] [PubMed]
- Funken, H.; Bartels, K.M.; Wilhelm, S.; Brocker, M.; Bott, M.; Bains, M.; Hancock, R.E.W.; Rosenau, F.; Jaeger, K.E. Specific association of lectin LecB with the surface of Pseudomonas aeruginosa: Role of outer membrane protein OprF. PLoS ONE 2012, 7, e46857. [Google Scholar] [CrossRef] [PubMed]
- Gilboa-Garber, N. Pseudomonas aeruginosa lectins. Methods Enzymol. 1982, 83, 378–385. [Google Scholar] [PubMed]
- Chen, C.P.; Song, S.C.; Gilboa-Garber, N.; Chang, K.S.; Wu, A.M. Studies on the binding site of the galactose-specific agglutinin PA-IL from Pseudomonas aeruginosa. Glycobiology 1998, 8, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Gilboa-Garber, N.; Katcoff, D.J.; Garber, N.C. Identification and characterization of Pseudomonas aeruginosa PA-IIL lectin gene and protein compared to PA-IL. FEMS Immunol. Med. Microbiol. 2000, 29, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Cioci, G.; Mitchell, E.P.; Gautier, C.; Wimmerova, M.; Sudakevitz, D.; Perez, S.; Gilboa-Garber, N.; Imberty, A. Structural basis of calcium and galactose recognition by the lectin PA-IL of Pseudomonas aeruginosa. FEBS Lett. 2003, 555, 297–301. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, E.; Houles, C.; Sudakevitz, D.; Wimmerova, M.; Gautier, C.; Perez, S.; Wu, A.M.; Gilboa-Garber, N.; Imberty, A. Structural basis for oligosaccharide-mediated adhesion of Pseudomonas aeruginosa in the lungs of cystic fibrosis patients. Nat. Struct. Biol. 2002, 9, 918–921. [Google Scholar] [CrossRef] [PubMed]
- Boukerb, A.M.; Rousset, A.; Galanos, N.; Mear, J.B.; Thepaut, M.; Grandjean, T.; Gillon, E.; Cecioni, S.; Abderrahmen, C.; Faure, K.; et al. Antiadhesive properties of glycoclusters against Pseudomonas aeruginosa lung infection. J. Med. Chem. 2014, 57, 10275–10289. [Google Scholar] [CrossRef] [PubMed]
- Gustke, H.; Kleene, R.; Loers, G.; Nehmann, N.; Jaehne, M.; Bartels, K.M.; Jaeger, K.E.; Schachner, M.; Schumacher, U. Inhibition of the bacterial lectins of Pseudomonas aeruginosa with monosaccharides and peptides. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Von Bismarck, P.; Schneppenheim, R.; Schumacher, U. Successful treatment of Pseudomonas aeruginosa respiratory tract infection with a sugar solution-a case report on a lectin based therapeutic principle. Klin. Pädiatrie 2001, 213, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Hauber, H.P.; Schulz, M.; Pforte, A.; Mack, D.; Zabel, P.; Schumacher, U. Inhalation with fucose and galactose for treatment of Pseudomonas aeruginosa in cystic fibrosis patients. Int. J. Med. Sci. 2008, 5, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Cassin, E.K.; Tseng, B.S. Pushing beyond the envelope: The potential roles of OprF in Pseudomonas aeruginosa biofilm formation and pathogenicity. J. Bacteriol. 2019, 201, e00050-19. [Google Scholar] [CrossRef] [PubMed]
- Kolomiets, E.; Swiderska, M.A.; Kadam, R.U.; Johansson, E.M.; Jaeger, K.E.; Darbre, T.; Reymond, J.L. Glycopeptide dendrimers with high affinity for the fucose-binding lectin LecB from Pseudomonas aeruginosa. Chem. Med. Chem. 2009, 4, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.M.; Crusz, S.A.; Kolomiets, E.; Buts, L.; Kadam, R.U.; Cacciarini, M.; Bartels, K.M.; Diggle, S.P.; Camara, M.; Williams, P.; et al. Inhibition and dispersion of Pseudomonas aeruginosa biofilms by glycopeptide dendrimers targeting the fucose-specific lectin LecB. Chem. Biol. 2008, 15, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, A.; Jimenez-Barbero, J.; Casnati, A.; De Castro, C.; Darbre, T.; Fieschi, F.; Finne, J.; Funken, H.; Jaeger, K.E.; Lahmann, M.; et al. Multivalent glycoconjugates as anti-pathogenic agents. Chem. Soc. Rev. 2013, 42, 4709–4727. [Google Scholar] [CrossRef] [PubMed]
- Reymond, J.L.; Bergmann, M.; Darbre, T. Glycopeptide dendrimers as Pseudomonas aeruginosa biofilm inhibitors. Chem. Soc. Rev. 2013, 42, 4814–4822. [Google Scholar] [CrossRef] [PubMed]
- Kadam, R.U.; Bergmann, M.; Hurley, M.; Garg, D.; Cacciarini, M.; Swiderska, M.A.; Nativi, C.; Sattler, M.; Smyth, A.R.; Williams, P.; et al. A glycopeptide dendrimer inhibitor of the galactose-specific lectin LecA and of Pseudomonas aeruginosa biofilms. Angew. Chem. Int. Ed. Engl. 2011, 50, 10631–10635. [Google Scholar] [CrossRef] [PubMed]
- Cecioni, S.; Imberty, A.; Vidal, S. Glycomimetics versus multivalent glycoconjugates for the design of high affinity lectin ligands. Chem. Rev. 2015, 115, 525–561. [Google Scholar] [CrossRef] [PubMed]
- Goyard, D.; Thomas, B.; Gillon, E.; Imberty, A.; Renaudet, O. Heteroglycoclusters with dual nanomolar affinities for the lectins LecA and LecB from Pseudomonas aeruginosa. Front. Chem. 2019, 7, 666. [Google Scholar] [CrossRef] [PubMed]
- Malinovska, L.; Thai, L.S.; Herczeg, M.; Vaskova, M.; Houser, J.; Fujdiarova, E.; Komarek, J.; Hodek, P.; Borbas, A.; Wimmerova, M.; et al. Synthesis of β-D-galactopyranoside-presenting glycoclusters, investigation of their interactions with Pseudomonas aeruginosa Lectin A (PA-IL) and evaluation of their anti-adhesion potential. Biomolecules 2019, 9, 686. [Google Scholar] [CrossRef] [PubMed]
- Leusmann, S.; Menova, P.; Shanin, E.; Titz, A.; Rademacher, C. Glycomimetics for the inhibition and modulation of lectins. Chem. Soc. Rev. 2023, 52, 3663–3740. [Google Scholar] [CrossRef] [PubMed]
- Rodrigue, J.; Ganne, G.; Blanchard, B.; Saucier, C.; Giguere, D.; Shiao, T.C.; Varrot, A.; Imberty, A.; Roy, R. Aromatic thioglycoside inhibitors against the virulence factor LecA from Pseudomonas aeruginosa. Org. Biomol. Chem. 2013, 11, 6906–6918. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, C.; Yu, Y.; Wei, X.; Duan, X.; Dai, X.; Zhang, X. Bioinspired heteromultivalent ligand-decorated nanotherapeutic for enhanced photothermal and photodynamic therapy of antibiotic-resistant bacterial pneumonia. ACS Appl. Mater. Interfaces 2019, 11, 39648–39661. [Google Scholar] [CrossRef] [PubMed]
- Sommer, R.; Wagner, S.; Rox, K.; Varrot, A.; Hauck, D.; Wamhoff, E.C.; Schreiber, J.; Ryckmans, T.; Brunner, T.; Rademacher, C.; et al. Glycomimetic, Orally bioavailable LecB inhibitors block biofilm formation of Pseudomonas aeruginosa. J. Am. Chem. Soc. 2018, 140, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Bartels, K.M.; Funken, H.; Knapp, A.; Brocker, M.; Bott, M.; Wilhelm, S.; Jaeger, K.E.; Rosenau, F. Glycosylation is required for outer membrane localization of the lectin LecB in Pseudomonas aeruginosa. J. Bacteriol. 2011, 193, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Kautto, L.; Nguyen-Khuong, T.; Everest-Dass, A.; Leong, A.; Zhao, Z.; Willcox, M.D.P.; Packer, N.H.; Peterson, R. Glycan involvement in the adhesion of Pseudomonas aeruginosa to tears. Exp. Eye Res. 2016, 145, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Boukerb, A.M.; Decor, A.; Ribun, S.; Tabaroni, R.; Rousset, A.; Commin, L.; Buff, S.; Doleans-Jordheim, A.; Vidal, S.; Varrot, A.; et al. Genomic rearrangements and functional diversification of leca and lecb lectin-coding regions impacting the efficacy of glycomimetics directed against Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 811. [Google Scholar] [CrossRef] [PubMed]
- Sommer, R.; Wagner, S.; Varrot, A.; Nycholat, C.M.; Khaledi, A.; Haussler, S.; Paulson, J.C.; Imberty, A.; Titz, A. The virulence factor LecB varies in clinical isolates: Consequences for ligand binding and drug discovery. Chem. Sci. 2016, 7, 4990–5001. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Chance, D.L.; Mossine, V.V.; Mawhinney, T.P. RAFT-based tri-component fluorescent glycopolymers: Synthesis, characterization and application in lectin-mediated bacterial binding study. Glycoconj. J. 2014, 31, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.D.; Evans, W.L. β-D-glucose-1,2,3,4-tetraacetate. Org. Synth. 1942, 22, 56–58. [Google Scholar] [CrossRef]
- Uchiyama, T.; Hindsgaul, O. Per-O-trimethylsilyl-a-l-fucopyranosyl iodide: A novel glycosylating agent for terminal α-L-fucosylation. Synlett 1996, 1996, 499–501. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Holloway, B.W. Genetic recombination in Pseudomonas aeruginosa. J. Gen. Microbiol. 1955, 13, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Hampton, K.D.; Wasilauskas, B.L. Isolation of oxidase-negative Pseudomonas aeruginosa from sputum culture. J. Clin. Microbiol. 1979, 9, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Chance, D.L.; Mawhinney, T.P. Carbohydrate sulfation effects on growth of Pseudomonas aeruginosa. Microbiology 2000, 146 Pt 7, 1717–1725. [Google Scholar] [CrossRef] [PubMed]
- Klockgether, J.; Munder, A.; Neugebauer, J.; Davenport, C.F.; Stanke, F.; Larbig, K.D.; Heeb, S.; Schock, U.; Pohl, T.M.; Wiehlmann, L.; et al. Genome diversity of Pseudomonas aeruginosa PAO1 laboratory strains. J. Bacteriol. 2010, 192, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Chance, D.L.; Mawhinney, T.P. Using negative staining TEM to study structure/function relationships of cystic fibrosis host-adapted opportunistic pathogen Pseudomonas aeruginosa. Microsc. Microanal. 2017, 23, 1354–1355. [Google Scholar] [CrossRef]
- Chance, D.; Wang, W.; Mawhinney, T. Probing cystic fibrosis Pseudomonas aeruginosa binding data for features that contribute to enhanced associations with α-Gal and β-GalNAc pendant monosaccharides of fluorescent glycopolymers. J. Cyst. Fibros. 2021, 20, S244. [Google Scholar] [CrossRef]
- Morris, A.J.; Jackson, L.; Yau, Y.C.W.Y.; Reichhardt, C.; Beaudoin, T.; Uwumarenogie, S.; Guttman, K.M.; Lynne, H.P.; Parsek, M.R.; Hoffman, L.R.; et al. The role of Psl in the failure to eradicate Pseudomonas aeruginosa biofilms in children with cystic fibrosis. NPJ Biofilms Microbiomes 2021, 7, 63. [Google Scholar] [CrossRef] [PubMed]
- Deligianni, E.; Pattison, S.; Berrar, D.; Ternan, N.G.; Haylock, R.W.; Moore, J.E.; Elborn, S.J.; Dooley, J.S. Pseudomonas aeruginosa cystic fibrosis isolates of similar RAPD genotype exhibit diversity in biofilm forming ability in vitro. BMC Microbiol. 2010, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Nolan, C.; Behrends, V. Sub-inhibitory antibiotic exposure and virulence in Pseudomonas aeruginosa. Antibiotics 2021, 10, 1393. [Google Scholar] [CrossRef] [PubMed]
- Bjarnsholt, T.; Whiteley, M.; Rumbaugh, K.P.; Stewart, P.S.; Jensen, P.O.; Frimodt-Moller, N. The importance of understanding the infectious microenvironment. Lancet Infect. Dis. 2022, 22, e88–e92. [Google Scholar] [CrossRef] [PubMed]
- Cullen, L.; Weiser, R.; Olszak, T.; Maldonado, R.F.; Moreira, A.S.; Slachmuylders, L.; Brackman, G.; Paunova-Krasteva, T.S.; Zarnowiec, P.; Czerwonka, G.; et al. Phenotypic characterization of an international Pseudomonas aeruginosa reference panel: Strains of cystic fibrosis (CF) origin show less in vivo virulence than non-CF strains. Microbiology 2015, 161, 1961–1977. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.E.; Buckley, D.G.; Wu, Z.; Saenphimmachak, C.; Hoffman, L.R.; D’Argenio, D.A.; Miller, S.I.; Ramsey, B.W.; Speert, D.P.; Moskowitz, S.M.; et al. Genetic adaptation by Pseudomonas aeruginosa to the airways of cystic fibrosis patients. Proc. Natl. Acad. Sci. USA 2006, 103, 8487–8492. [Google Scholar] [CrossRef] [PubMed]
- Reichhardt, C.; Jacobs, H.M.; Matwichuk, M.; Wong, C.; Wozniak, D.J.; Parsek, M.R. The versatile Pseudomonas aeruginosa biofilm matrix protein CdrA promotes aggregation through different extracellular exopolysaccharide interactions. J. Bacteriol. 2020, 202, e00216-20. [Google Scholar] [CrossRef] [PubMed]
- Vishwanath, S.; Guay, C.M.; Ramphal, R. Effects of subminimal inhibitory concentrations of antibiotics on the adherence of Pseudomonas aeruginosa to tracheobronchial mucin. J. Antimicrob. Chemother. 1987, 19, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Carfartan, G.; Gerardin, P.; Turck, D.; Husson, M.O. Effect of subinhibitory concentrations of azithromycin on adherence of Pseudomonas aeruginosa to bronchial mucins collected from cystic fibrosis patients. J. Antimicrob. Chemother. 2004, 53, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Mojsoska, B.; Ghoul, M.; Perron, G.G.; Jenssen, H.; Alatraktchi, F.A. Changes in toxin production of environmental Pseudomonas aeruginosa isolates exposed to sub-inhibitory concentrations of three common antibiotics. PLoS ONE 2021, 16, e0248014. [Google Scholar] [CrossRef] [PubMed]
- Mossine, V.V.; Chance, D.L.; Waters, J.K.; Mawhinney, T.P. Interaction of bacterial phenazines with colistimethate in bronchial epithelial cells. Antimicrob. Agents Chemother. 2018, 62, e02349-17. [Google Scholar] [CrossRef] [PubMed]
- Chance, D.; Wang, W.; Mawhinney, T. Targeting cystic fibrosis Pseudomonas aeruginosa with galactose related glycomimics: α- and ß-anomeric configurations, N-acetylation, and sulfation in whole-cell binding. J. Cyst. Fibros. 2023, 22, S2. [Google Scholar] [CrossRef]
- Lowery, N.V.; McNally, L.; Ratcliff, W.C.; Brown, S.P. Division of labor, bet hedging, and the evolution of mixed biofilm investment strategies. mBio 2017, 8, e00672-17. [Google Scholar] [CrossRef] [PubMed]
- El Meouche, I.; Siu, Y.; Dunlop, M.J. Stochastic expression of a multiple antibiotic resistance activator confers transient resistance in single cells. Sci. Rep. 2016, 6, 19538. [Google Scholar] [CrossRef] [PubMed]
- Papa, R.; Imperlini, E.; Trecca, M.; Paris, I.; Vrenna, G.; Artini, M.; Selan, L. Virulence of Pseudomonas aeruginosa in cystic fibrosis: Relationships between normoxia and anoxia lifestyle. Antibiotics 2023, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Zopf, D.; Roth, S. Oligosaccharide anti-infective agents. Lancet 1996, 347, 1017–1021. [Google Scholar] [CrossRef] [PubMed]
- Loris, R.; Tielker, D.; Jaeger, K.E.; Wyns, L. Structural basis of carbohydrate recognition by the lectin LecB from Pseudomonas aeruginosa. J. Mol. Biol. 2003, 331, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Heggelund, J.E.; Varrot, A.; Imberty, A.; Krengel, U. Histo-blood group antigens as mediators of infections. Curr. Opin. Struct. Biol. 2017, 44, 190–200. [Google Scholar] [CrossRef] [PubMed]
- de Bentzmann, S.; Roger, P.; Dupuit, F.; Bajolet-Laudinat, O.; Fuchey, C.; Plotkowski, M.C.; Puchelle, E. Asialo GM1 is a receptor for Pseudomonas aeruginosa adherence to regenerating respiratory epithelial cells. Infect. Immun. 1996, 64, 1582–1588. [Google Scholar] [CrossRef] [PubMed]
- Chance, D.L.; Mawhinney, T.P. Disulfated oligosaccharides derived from tracheobronchial mucous glycoproteins of a patient suffering from cystic fibrosis. Carbohydr. Res. 1996, 295, 157–177. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| General Scheme 1 | Glycopolymer Abbreviation | Pendant Monosaccharide |
|---|---|---|
 | α-Gal-PAA-Fluor | α-D-Galactose |
| β-Gal-PAA-Fluor | β-D-Galactose | |
| α-GalNAc-PAA-Fluor | α-D-N-Acetylgalactosamine | |
| β-GalNAc-PAA-Fluor | β-D-N-Acetylgalactosamine | |
| β-GlcNAc-PAA-Fluor | β-D-N-Acetylglucosamine | |
| β-Gal3S-PAA-Fluor | β-D-Galactose-3-sulfate | |
| β-GalNAc3S-PAA-Fluor | β-D-N-Acetylgalactosamine-3-sulfate | |
| α-Neu5Ac-PAA-Fluor | α-D-N-Acetylneuraminic acid | |
| α-L-Fuc-PAA-Fluor | α-L-Fucose |
| Culture ID | Plot Order | Isolate or Strain Source 1 (Sputum, Throat, Other) | Characteristics 2,3 |
|---|---|---|---|
| CF-S 8314-1 | 1 | CF sputum, pair #1 | Nonmotile, nonmucoid, blue-green diffusible pigment |
| CF-S 8314-2 | 2 | CF sputum, pair #1 | Mucoid |
| CF-S 8981-1 | 3 | CF sputum, pair #2 | Nonmotile, nonmucoid; M9 media restricted growth, TSB cultured |
| CF-S 8981-2 | 4 | CF sputum, pair #2 | Mucoid, brown diffusible pigment on TSA |
| CF-S 3318 | 5 | CF sputum | Motile; M9 media restricted growth, TSB cultured |
| CF-S 3247 | 6 | CF sputum | Motile |
| CF-S 3396 | 7 | CF sputum | Mucoid |
| CF-S 3443 | 8 | CF sputum | Small colony variant (SCV), nonmotile, nonmucoid |
| CF-T 3371 | 9 | CF throat, pair #3 | Nonmotile, nonmucoid; aggregated in M9 media, TSB cultured |
| CF-T 3372 | 10 | CF throat, pair #3 | Mucoid |
| CF-T 3435 | 11 | CF throat | Motile |
| CF-T 3437 | 12 | CF throat | Motile; M9 media restricted growth, TSB cultured |
| CF-T 3446 | 13 | CF throat | Motile |
| NCF-H 3380 | 14 | non-CF, hip infection | Motile |
| NCF-S 3391 | 15 | non-CF, sputum | Motile |
| ATCC BAA-47 | 16 | other—wound | Motile, green diffusible pigment |
| ATCC 15442 | 17 | animal room water bottle | Motile |
| ATCC 33468 | 18 | sputum; not CF tested | Mucoid |
Examples of P. aeruginosa phenotypes on MacConkey Agar (a) and Pseudomonas P Agar (b) 4. | |||
| Source and Culture ID | (#) | Colony Phenotype | Structural Features Confirmed | Preferred Glycopolymer Binding Observed Microscopically Fluorescent Avg # Cells/Field > 1000 Cells | Preferred Glycopolymer Binding Measured Spectroflurometrically Fluorescence Intensity > 6000 AU |
|---|---|---|---|---|---|
| CF sputum CF-S 8314-1 | 1 | nonmucoid | flagella, pili | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF sputum CF-S 8314-2 | 2 | mucoid | flagella | α-Gal | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF sputum CF-S 8981-1 | 3 | nonmucoid | flagella | β-GalNAc | α-Gal, β-GalNAc, β-Gal3S |
| CF sputum CF-S 8981-2 | 4 | mucoid | flagella, pili | - | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF sputum CF-S 3318 | 5 | motile | flagella, pili | α-Gal | α-Gal, β-GalNAc, β-Gal3S |
| CF sputum CF-S 3247 | 6 | motile | flagella, pili | - | α-Gal, β-GalNAc, β-Gal3S |
| CF sputum CF-S 3396 | 7 | mucoid | flagella, pili | β-GalNAc, β-Gal3S | α-Gal, β-GalNAc, β-Gal3S |
| CF sputum CF-S 3443 | 8 | nonmucoid, (SCV) | flagella, pili | α-Gal, β-GalNAc, β-Gal3S | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF throat CF-T 3371 | 9 | nonmucoid | flagella | β-GalNAc | α-Gal, β-GalNAc, β-Gal3S |
| CF throat CF-T 3372 | 10 | mucoid | flagella, pili | α-Gal, β-GalNAc | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF throat CF-T 3435 | 11 | motile | flagella, pili | β-GalNAc | α-Gal, β-GalNAc, β-Gal3S |
| CF throat CF-T 3437 | 12 | motile | flagella, pili | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| CF throat CF-T 3446 | 13 | motile | flagella, pili | β-GalNAc | α-Gal, β-GalNAc, β-Gal3S |
| Non-CF sputum NCF-S 3391 | 15 | motile | pili | α-Gal, β-Gal3S | α-Gal, β-GalNAc, β-Gal3S, α-Neu5Ac |
| Non-CF sputum ATCC 33468 | 18 | mucoid | pili | α-Gal, β-GalNAc | α-Gal, β-GalNAc, β-Gal3S |
| Other NCF-H 3380 | 14 | motile | flagella, pili | β-Gal3S | α-Gal, β-GalNAc, β-Gal3S |
| Other ATCC BAA-47 | 16 | motile | flagella | - | α-Gal, β-GalNAc, β-Gal3S |
| Other ATCC 15442 | 17 | motile | flagella, pili | - | α-Gal, β-GalNAc, β-Gal3S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chance, D.L.; Wang, W.; Waters, J.K.; Mawhinney, T.P. Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides. Microorganisms 2024, 12, 801. https://doi.org/10.3390/microorganisms12040801
Chance DL, Wang W, Waters JK, Mawhinney TP. Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides. Microorganisms. 2024; 12(4):801. https://doi.org/10.3390/microorganisms12040801
Chicago/Turabian StyleChance, Deborah L., Wei Wang, James K. Waters, and Thomas P. Mawhinney. 2024. "Insights on Pseudomonas aeruginosa Carbohydrate Binding from Profiles of Cystic Fibrosis Isolates Using Multivalent Fluorescent Glycopolymers Bearing Pendant Monosaccharides" Microorganisms 12, no. 4: 801. https://doi.org/10.3390/microorganisms12040801