
The Impact of Cr(III) and Cr(VI) on Lipid Accumulation in Chlorella pyrenoidosa
by
 ,
,
Tianji Liu
†,
Huawei Guo
†,
Qing Yu
,
Yajun Wang
,
Huan Liu
,
Yanan Zeng
,
Yitong Wang
*,
Chunyu Liu
* and
Junguo Li
College of Metallurgy and Energy, North China University of Science and Technology, 21 Bohai Street, Tangshan 063210, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this manuscript.
Processes 2024, 12(5), 905; https://doi.org/10.3390/pr12050905
Submission received: 3 April 2024
/
Revised: 24 April 2024
/
Accepted: 24 April 2024
/
Published: 29 April 2024
(This article belongs to the Section Biological Processes and Systems)
Abstract
:In this study, Chlorella pyrenoidosa was cultured with different concentrations of Cr(III) and Cr(VI), and the changes in biomass, lipid content, protein content and total sugar content of Chlorella pyrenoidosa under different culture conditions were detected. The effects of Cr(III) and Cr(VI) on the intracellular antioxidant system and related key enzymes of lipid synthesis were analyzed. The results showed that appropriate Cr(III) and Cr(VI) concentrations could make Chlorella pyrenoidosa regulate the contents of SOD, POD, ASA, CAT and GSH in cells, improve the enzyme activity of ACC and promote lipid synthesis. The aim was to investigate the mechanism of lipid synthesis by analyzing the distribution of Cr(III) and Cr(VI) in algal cells. The lipid content of Chlorella pyrenoidosa reached its highest level at 50.43 wt% and 38.00 wt% when both Cr(III) and Cr(VI) concentrations in the culture environment were 0.5 mg/L. These values were 1.37 and 1.03 times higher than the control group, respectively. The addition of appropriate Cr(III) and Cr(VI) concentrations can increase ME enzyme activity and decrease PEPC enzyme activity, shorten the biomass accumulation cycle and prolong the lipid synthesis cycle.
1. Introduction
There are three main forms of Cr in nature: one is the uncharged metallic form (Cr), another is the tri-valent form (Cr(III)) and the last is the six-valent form (Cr(VI)) [1]. In humans, Cr(III) is a micronutrient essential for glucose, lipid and protein metabolism. In contrast, Cr(VI) has no recognized biological function and is a potent carcinogen [2]. In the water, Cr existsas Cr(VI) in the body of fish and reaches the human body through the food chain [3]. The consumption of plants growing on contaminated soil will cause serious harm to human health due to the bioaccumulation of toxic heavy metals [4]. Cr(VI) is 100 times more poisonous than Cr(III), has high solubility, bioavailability, fluidity and biotoxicity and has been listed as one of the 20 most toxic pollutants [5]. Cr(VI) has a strong oxidizing ability and can change the expression of plant genes and proteins, thus disturbing plant metabolism. The U.S. Environmental Protection Agency demands that the amount of total chromium released into surface water should not exceed 2 mg/L and the level of Cr(VI) must not surpass 0.05 mg/L [6]. There are several common methods to address Cr, which primarily encompass physical, chemical and biological approaches [7]. The first two methods are easy to cause secondary pollution and cannot meet the national emission limits, which costs a lot and lacks efficiency. The biological method is a strategy that uses biological organisms such as bacteria, fungi, plants and algae to restore the environment to its original state. The biological processes include biosorption, bioaccumulation and biotransformation mechanisms for pollutant detoxification, which is a natural process [8]. In the case of bacteria and fungi, to promote their growth they need to supplement with additional sources of nutrients [9]. The efficiency of the remediation of plants largely depends on the type of aquatic plants that accumulate heavy metals. The efficacy of plants depends upon a wide range of root systems, as they need a long amount of time to cultivate and produce aquatic plants [10]. Nevertheless, microalgae could potentially serve as an environmentally friendly method of remediation. Microalgae are photosynthetic organisms that can rapidly reproduce, use sunlight, water and carbon dioxide to convert into chemical energy and can produce biomass through the use of the nutrients present in wastewater while producing biomass that can be used to produce different value-added products [11], which can reduce additional nutrient requirements. Meanwhile, microalgae have excellent adsorption performance against Cr, which is an excellent candidate for removing Cr from wastewater. The excessive deposition of chromium in plant cells leads to alterations in enzyme activity, degradation of chlorophyll, reduction in biomass and disruption of the antioxidant defense system [12]. This applies to both Cr(III) and Cr(VI) which have varying levels of bioavailability, mobility and toxicity [13,14]. The removal of chromium by microalgae cells occurs not only through adsorption, but also through biological mechanisms. One example is the enzymatic reduction of Cr(VI) to Cr(III) as a means of alleviating its toxicity. When Chlorella vulgaris is under Cr(VI) stress will release Glutathione (GSH) which can reduce Cr(VI) to Cr(III). Additionally, most Cr(VI) is aggregated in organelles of microalgae cells [15]. Existing research focuses on the tolerance ability and removal ability of microalgae to Cr(VI). Lu et al. have studied Chlorella vulgaris in bioaccumulation of Cr(VI), where they found that the growth, chlorophyll content, oxidative damage and Cr(VI) removal rate under Cr(VI) stress were evaluated [16]. Shen et al. found that in the presence of added Cr (VI) and sodium acetate in Botryocossuss sp. NJD-1 for Cr (VI) removal of 94.2% [6].
According to the above research, microalgae have been found to effectively remove Cr(VI). Some studies have shown that microalgae not only produce enough biomass and accumulate a significant amount of lipids under Cr stress, but also have the ability to bioremediate Cr(VI) within the toxic concentration limits for the human body (1 mg/L) [1]. Kafil et al. showed that Chlorella under Cr(VI) stress can change the content of fatty acid methyl esters in microalgae, thereby improving the quality of biodiesel [17].
Nevertheless, there is a lack of comprehensive research on the relationship between Cr and lipid accumulation in microalgae. Hence, the primary focus of this study is to assess the effects of Cr(III) and Cr(VI) on Chlorella pyrenoidosa biomass, lipid content, antioxidant system and key enzymes associated with lipid synthesis. Cr(III) and Cr(VI) were studied in valence transformation under the action of Chlorella pyrenoidosa. This study guides large-scale energy microalgae cultivation and high value-added utilization of Cr-containing waste.
2. Materials and Methods
The materials used in the experiment are presented in Section 2.1 and information on analytical methods and instrument parameters is shown in Section 2.2 and Section 2.3.
2.1. Materials
The Chlorella pyrenoidosa used in the experiment was purchased from Nanjing Haiersi Biological Co., Ltd. (Nanjing, China). Table 1 lists the chemical reagents used for this study. Deionized water (pH: 5.83; ionization constant: 18.25 MΩ·cm) was from the ultrapure water system (Exceed-Ad-32, Chengdu Eco Water Treatment Equipment Co., Ltd., Chengdu, China). The ultraviolet/visible/near-infrared spectrophotometer used in the experiment came from Hitachi Manufacturing Co., LTD. (UH4150, Hitachi Manufacturing Co., Ltd., Tokyo, Japan).
2.2. Preparation of Cr Source Medium and Cultivation of Chlorella pyrenoidosa
2.2.1. Preparation of Cr Source Medium
Chlorella pyrenoidosa was cultured in BG11 medium. Table 2 lists the compositions of BG11 medium used for this study. The BG11 medium has the following compositions (g/1 L ultrapure water): 1.5 NaNO3; 0.04 K2HPO4; 0.75 MgSO4·7H2O; 0.006 C6H8O7; 0.006 C6H11FeNO7; 0.001 Na2EDTA; 0.036 CaCl2·2H2O; 0.02 Na2CO3; 0.00286 H3BO3; 0.00182 MnCl4·4H2O; 0.00022 ZnSO4·7H2O; 0.0004 Na2MoO4·2H2O; 0.00008 CuSO4·5H2O; 0.0005 Co(NO3)2·6H2O.
Weigh CrCl3·6H2O (molecular weight: 266.44) 0.51 g and dissolve with BG11, then transfer into 1000 mL volumetric bottle and set the volume to 1000 mL, then Cr(III) = 100 mg/L. BG11 was employed for diluting Cr(III) concentrations at 0.5, 1.0, 2.0 and 5.0 mg/L, respectively. The pH was regulated to a range of 6.8 to 7.1 using NaOH and HCl and was subsequently sterilized at 121 °C for 20 min. K2Cr2O7 (molecular weight: 294.18) 0.28 g was weighed and diluted in constant volume according to Cr(III) configuration steps. After dilution, Cr(VI) concentrations were 0.5, 1.0, 2.0 and 5.0 mg/L, respectively.
2.2.2. Cultivation of Chlorella pyrenoidosa
Before inoculation, 1 mol/L of NaOH or HCl was used to adjust the pH of Cr source culture medium to 7.1 and then transferred to 1000 mL conical bottle. (DGL-50B, Shanghai Lichen Bangxi Instrument Technology Co., Ltd., Shanghai, China) at 121 °C for 20 min and cooled to 25 °C naturally. A total of 50 mL of Chlorella pyrenoidosa seed was inoculated into the sterilized medium (1000 mL). All the operations mentioned above were carried out on the clean bench (SW-CJ-1D, Shanghai Lichen Bangxi Instrument Technology Co., Ltd., China). The Cr source medium (1000 mL) was transferred to an illuminating incubator (GXZ-500B, Ningbo Jiangnan Instrument Factory, Ningbo, China) with a culture temperature of 25 °C, light intensity of 4000 Lux and light/darkness of 12/12. For experimental accuracy and to avoid microalgae sinking, three bottles were made for each group of experiments and were shaken three times per day.
2.3. Analysis of Chlorella pyrenoidosa
2.3.1. Analysis of Chlorella pyrenoidosa Lipid Content
We analyzed the biomass and lipid content of Chlorella pyrenoidosa after 25 days of being cultured in light. The algal liquid of 1000 mL was separated into batches and centrifuged at 8000 rpm for 15 min using a high-speed centrifuge. After removing the supernatant, the sediment was then transferred to a freeze dryer −50 °C until a constant weight was achieved [18]. The weight of the sample after it has dried is used to determine the biomass. Dried Chlorella pyrenoidosa powder (0.10–0.30 g) and quartz sand with a mass ratio of 1/1 were mixed and ground, sealed with filter paper (to prevent Chlorella pyrenoidosa powder from entering the distillation bottle during Soxhlet extraction) and transferred to a 250 mL Soxhlet extractor (SZF-06A, Shanghai Lichen Bangxi Instrument Technology Co., Ltd., China). Petroleum ether (200 mL) was added into a distillation flask equipped with a reflux condenser, heated to 90 °C for 12 h and all solvents were removed at 75 °C. Lipid content of Chlorella pyrenoidosa (wt%) was calculated according to Equation (1).
Lipid content of Chlorella pyrenoidosa = (W2/W1) × 100%,
W1: mass of dried Chlorella pyrenoidosa powder, g; W2: mass of crude lipid, g.
2.3.2. Analysis of Chlorella pyrenoidosa Biomass
The absorbance of the cultured algal liquid was measured at 680 nm and the Chlorella pyrenoidosa biomass was determined according to the study of Lu et al. [16]. Equation (2) was used to calculate the biomass of Chlorella pyrenoidosa.
Biomass = 0.25 × A680 + 0.001, (R2 = 0.995)
A680: absorbance value at 680 nm.
2.3.3. Analysis of Chlorella pyrenoidosa Enzyme Activity
Approximately 0.10 g Chlorella pyrenoidosa powder (fresh weight) was mixed with 1 mL extraction solution to be homogenized in an ice bath where they were centrifuged at 4 °C for 10 min with the rate of 8000× g and then kept on ice. The pretreated sample was analyzed using a quantitative detection kit (cominbio, Suzhou, China) to measure the levels of reactive oxygen species (ROS), superoxide dismutase (SOD) activity, malondialdehyde (MDA) content, peroxidase (POD) activity, acetyl-CoA carboxylase (ACC) activity, malic enzyme (ME) activity, phosphoenolpyruvate carboxylase (PEPC) activity, protein content and total sugar content.
2.3.4. Analysis of Cr(III) and Cr(VI)
GB7467-1987 was used to determine the concentration of Cr(VI), weigh 0.2829g of K2Cr2O7 dried at 120 °C for 2 h and prepare it into a Cr(VI) concentration of 100 mg/L solution and dilute it with deionized water to a Cr(VI) concentration of 1.0 mg/L solution. A total of 0.20 g of diphenylcar bazide was dissolved in 50 mL acetone and diluted with water to 100 mL to prepare the Cr(VI) color-developing agent. Cr (VI) solution (concentration of 0, 0.004, 0.01, 0.02, 0.04, 0.08, 0.12, 0.16 and 0.20 mg/L) were prepared by adding 0, 0.2, 0.5, 1, 2, 4, 6, 8 and 10 mL of Cr (VI) solution (concentration of 1.0 mg/L) into a 50 mL volumetric flask and adding ultrapure water to set the volume to 50 mL, then add 1 mL of H3PO4 solution (the volume ratio of H3PO4 to water is 1/1) and 2 mL of color developer respectively and shake well. The absorbance of the solution after color development was measured at 540 nm wavelength 5–10 min after color development. The absorbance of Cr(VI) at 540 nm is depicted in Figure 1. Equation (3) is for calculating the concentration of Cr(VI).
C = 1.8009 × A540 − 0.0002, (R2 = 0.9991)
C: concentration of Cr(VI), mg/L; A540: absorbance value at 540 nm.
The total chromium content was determined by the ICP-OES instrument and the content of Cr(VI) was subtracted from the total chromium content to obtain the content of Cr(III) in the liquid phase.
2.3.5. Statistical Analysis
The obtained biological data were subjected to statistical analysis using SPSS version 22.0. The statistically significant differences in the mean of the applied rates were analyzed with ANOVA followed by Duncan’s test at a level of p < 0.05.
3. Results
Chlorella pyrenoidosa was cultivated in media containing Cr(III) and Cr(VI) at concentrations of 0.0, 0.5, 1.0, 2.0 and 5.0 mg/L. The biomass, total sugar, protein and lipid contents of Chlorella pyrenoidosa were then quantified. The impact of Cr(III) and Cr(VI) on lipid accumulation in Chlorella pyrenoidosa is presented in Figure 2 and Figure 3 to provide clarity. The levels of SOD, MDA, POD, GSH and AsA were assessed following a 10 day culture of Chlorella pyrenoidosa under a controlled environment. The study elucidated the impact of Cr(III) and Cr(VI) on the antioxidant system and lipid peroxidation performance of Chlorella pyrenoidosa, with the findings featured in Figure 4. The removal ability of Chlorella pyrenoidosa to total Cr and the determination results of Cr(III) and Cr(VI) distribution in algal cells are shown in Figure 5. To uncover the regulatory mechanism of Cr(III) and Cr(VI) on the lipid accumulation of Chlorella pyrenoidosa, we investigated the alterations in acetyl-CoA carboxylase (ACC), malic enzyme (ME) and phosphoenolpyruvate carboxylase (PEPC) content after 10 days of Chlorella pyrenoidosa culture in a specific environment (Figure 6 and Figure 7).
3.1. Effects of Cr(III) and Cr(VI) on Biomass
As illustrated in Figure 2, when the concentration of Cr(III) in the culture environment increased from 0.5 mg/L to 5.0 mg/L, the biomass of algal cells decreased from 104 mg/L to 62 mg/L, which was lower than the control group (124 mg/L). Similar results were found in the culture environment with the addition of Cr(VI), where the concentration of Cr(VI) increased from 0.5 mg/L to 2.0 mg/L, the biomass of Chlorella pyrenoidosa decreased from 93 mg/L to 9 mg/L and the growth of Chlorella pyrenoidosa was completely inhibited.
3.2. Cr(III) and Cr(VI) on Effect of Lipid, Protein and Total Sugar Content
Lipid content: when the concentration of both Cr(III) and Cr(VI) in the culture environment was 0.5 mg/L, the lipid content of Chlorella pyrenoidosa reached its maximum at 50.43 wt% and 38.00 wt%, respectively, compared to the control group (36.87 wt%) (Figure 3a). With the increase of Cr(III) or Cr(III) concentration, lipid content tended to decrease (the lipid content corresponding to 5 mg/L Cr(III) decreased to 24.88 wt%; lipid content corresponding to 5 mg/L Cr(VI) decreased to 21.87 wt%.
Protein content: a low concentration of Cr(III) is helpful for the protein generation of Chlorella pyrenoidosa (Figure 3b). When Cr(III) is 0.5 mg/L, the protein content of Chlorella pyrenoidosa is 13.60 mg/g fresh weight (FW), 1.09 times that of the control group. As the concentration of Cr(III) in the medium continued to rise, there was a decrease in protein content within the algal cells. At a concentration of 1.0 mg/L FW, the protein content reached 11.23 mg/g FW, which was lower than the control group. However, the presence of Cr(VI) was unfavorable to the protein synthesis of Chlorella pyrenoidosa. The medium experienced a rise in Cr(VI) concentration from 0.5 to 5.0 mg/L, coinciding with a gradual decline in protein content from 8.42 mg/g FW to 3.45 mg/g FW.
Total sugar content: the total sugar content in Chlorella pyrenoidosa demonstrated an increasing trend followed by a decreasing trend as the concentrations of Cr(III) and Cr(VI) increased (Figure 3c). During the culture process with Cr(III) added, the total sugar content of Chlorella pyrenoidosa was lower than that of the control group at 32.83 mg/g FW and the highest value was achieved at 1.0 mg/L Cr(III), where the total sugar content of Chlorella pyrenoidosa was 30.66 mg/g FW. When 1.0 mg/L Cr(VI) was added to the medium, the total sugar content of Chlorella pyrenoidosa was 37.28 mg/g FW, higher than that of the control group. It reached the maximum value of 48.32 mg/g FW when Cr(VI) concentration was 2.0 mg/L.
3.3. Chlorella pyrenoidosa of Cr(III) and Cr(VI) Stress Response
After 10 days, Chlorella pyrenoidosa was cultured in a medium containing concentrations of 0.5–5.0 mg/L of Cr(III) and Cr(VI) sources. The levels of SOD, CAT, POD as well as non-enzymatic antioxidants (GSH and AsA) were measured in the algae cells to clarify the effects of Cr(III) and Cr(VI) stress on antioxidant and non-antioxidant enzymes of Chlorella pyrenoidosa.
Antioxidant enzymes (SOD, CAT and POD): the SOD and CAT activities of Chlorella pyrenoidosa in the medium supplemented with 0.5–5.0 mg/L Cr(III) exhibited a pattern of initially increasing and subsequently decreasing (Figure 4a,b). In the medium supplemented with 2.0 mg/L Cr(III), the SOD and CAT activities of Chlorella pyrenoidosa showed a trend of decreasing. SOD and CAT activities reached the maximum 136.98 U/mg prot and 30.91 U/mg prot respectively, which were 1.99 and 2.98 times that of the control group. A comparable trend was observed in the medium containing 0.5–5.0 mg/L of Cr(VI) as well. When the concentration of Cr(VI) was 1.0 mg/L and 2.0 mg/L, the activities of SOD and CAT reached the maximum of 101.81 and 24.76 U/mg prot, which were 1.48 and 2.39 times that of the control group. In Figure 4c, POD activity reached its maximum value at 1.0 mg/L Cr(III) and 5.0 mg/L Cr(VI), 34.65 and 49.24 U/mg prot, respectively, 1.43 and 2.03 times that of the control group (POD activity of the control group: 24.26 U/mg prot).
Non-enzymatic antioxidants (GSH and AsA): when Cr(III) and Cr(VI) were added to the culture environment increased from 0.5 mg/L to 1.0 mg/L, the GSH content of Chlorella pyrenoidosa under Cr(III) treatment increased from 216.64 nmol/g in FW (Cr(VI) environment (Figure 4d). The amount of 105.54 nmol/g FW was increased to the maximum value of 290.29 nmol/g FW (GSH content in Cr(VI) environment (283.33 nmol/g FW) in the control group (GSH content: 3.15 and 3.07 times of 92.19 nmol/g FW), Cr(III) and Cr(VI) concentrations continued to increase and GSH content began to decline. The AsA content of Chlorella pyrenoidosa decreased with the increase of Cr(III) and Cr(VI) concentrations in the culture environment (Figure 4e). Compared with the control group, the AsA content was higher only under 0.5 and 1.0 mg/L Cr(III) conditions. To further understand the influence of AsA on Cr(VI) in Chlorella pyrenoidosa, the reduction rate of Cr(VI) at different ratios of Cr(VI)/AsA was explored (Figure 4f). With the increase of ascorbic acid, the reduction rate of Cr(VI) gradually increased. In water containing 5.0 mg/L Cr(VI), the three concentration ratios of Cr(VI)/AsA were 1/1, 1/2 and 1/3, respectively, and the reduction rates were 5.62%, 27.62% and 45.27%.
3.4. Distribution of Cr(III) and Cr(VI) in Algal Cells
A concentration of 0.5 mg/L for both Cr(III) and Cr(VI) was chosen as the specific culture conditions to assess the capability of Chlorella pyrenoidosa in removing total Cr from the culture environment. Figure 5a illustrates that the removal rate for Cr(III) reached 65.00% by the end of the culture period. For Cr(VI), there was no significant decrease in total Cr concentration during cultivation, and the final removal rate was only 11.53%.
The distribution of Cr(III) and Cr(VI) within algae cells was analyzed by selecting a Cr(VI) concentration of 0.5 mg/L as a specific culture condition. According to Figure 5b, the distribution of Cr(VI) and Cr(III) content in algae cells was observed under specific culture conditions with 0.5 mg/L of Cr(VI). The active compounds found in microalgae transform Cr(VI) into Cr(III) partially, thus reducing the toxicity of Cr(VI).
3.5. The Effect of Cr (III) and Cr (VI) on the Activity of ACC, ME and PEPC
Chlorella pyrenoidosa was cultured in Cr(III) and Cr(VI) source medium with concentrations of 0.5–5.0 mg/L. ACC, ME and PEPC were found in algal cells after a period of 10 days. The objective is to investigate the impact of Cr(III) and Cr(VI) stress on the enzymes involved in lipid synthesis in Chlorella pyrenoidosa.
Acetyl-CoA carboxylase (ACC): the ACC activity of Chlorella pyrenoidosa varied with the concentration of Cr (Figure 6a). Compared to the control group (8.05 μmol/h/mg prot), it initially increased and then decreased. ACC activity in Cr(III) and Cr(VI) at 2.0 mg/L reached a maximum of 13.67 and 14.32 μmol/h/mg prot, respectively, which was 1.70 and 1.78 times that of the control group. The results were inconsistent with the results of the lipid content of Chlorella pyrenoidosa (when Cr(III) or Cr(VI) concentrations were both 0.5 mg/L in the culture environment, the lipid content of Chlorella pyrenoidosa reached the maximum 50.43 wt% and 38.00 wt%).
Malic enzyme (ME): the activity of ME was higher than the control group in the concentration range of 0.5–5.0 mg/L for Cr(III) and Cr(VI) (Figure 6b). At 2.0 mg/L Cr(III) and 5.0 mg/L Cr(VI), the maximum prot values were 52.96 and 54.31 nmol/h/mg, respectively, which were 5.03 and 5.16 times that of the control group.
Phosphoenolpyruvate carboxylase (PEPC): at concentrations of 0.5 and 1.0 mg/L Cr(III), the PEPC activity of Chlorella pyrenoidosa was 12.72 nmol/min/mg prot and 10.43 nmol/min/mg prot, respectively, lower than that of the control group (Figure 6c). At 0.5 mg/L and 1.0 mg/L Cr(VI) concentrations, the PEPC activity of Chlorella pyrenoidosa was 11.82 nmol/min/mg prot and 6.31 nmol/min/mg prot, respectively, lower than that of the control group.
4. Discussion
4.1. Effects of Cr(III) and Cr(VI) on Biomass
Cr(VI) can be converted into Cr(V) complexes by physiological reducing agents, including NADPH, various pentoses, glutathione and others. The toxic effects of Cr are associated with the reduction of Cr(VI) to a lower oxidation state, particularly the instantaneous formation of Cr(V). The Cr(V) complex reacts with H2O2 to produce a significant amount of OH− radicals instead of the corresponding O2−, which can directly induce alterations in DNA and the synthesis of other harmful compounds [19]. Therefore, it is particularly important to explore the effects of Cr(III) and Cr(VI) on the physiological and biochemical indexes of Chlorella pyrenoidosav. It has been found that when the concentration of Cr(VI) increases from 5 ppm to 10 ppm, the growth rate of microalgae will be adversely affected, resulting in a decrease in biomass density [17]. Cr(VI) induces oxidative stress in microalgal cells, inhibiting electron transport and the absorption, transfer and utilization of light energy, resulting in this phenomenon [20].
4.2. Cr(III) and Cr(VI) on Effect of Lipid, Protein and Total Sugar Content
Lipid content: microalgae synthesize lipids by using CO2 or glucose under environmental stress (Figure 3a) [21]. Cr(III) and Cr(VI) as stress factors had positive effects on lipid accumulation in microalgae in the culture environment. Obviously, the addition of Cr(III) or Cr(VI) during culture triggered the production of a large number of ROS by microalgal cells, which activated different protective mechanisms and intensified the accumulation of secondary metabolites (such as lipids) [22]. According to the report, it was found that Scenedesmus quadricauda increased the lipid content of Cr(VI) at low concentrations [20]. However, high concentrations of Cr can cause damage to intracellular structures and inhibit cell growth, thereby reducing lipid content.
Protein content: Sharma et al. also reported similar results, diluting concentrations of 75% wastewater to cultivate algae group MAC2 (Chlorella sp., Nannochloropsis sp., Scenedesmus dimorphus, Kirchnella and Microcoleus) (Figure 3b). After 10 days, the protein content was higher than that of the control group, and when the wastewater concentration was increased to 100%, the protein content was lower than that of the control group [23]. The increase in protein content may result from the synthesis of certain stress proteins or metal-chelating proteins. The presence of high concentrations of Cr(III) and Cr(VI) will lead to protein degradation and is used for the growth of Chlorella pyrenoidosa under high Cr stress [24].
Total sugar content: the increase in total sugar content can be attributed to certain changes in the photosynthetic mechanism, enhanced cell division and synthesis of proteins and osmoprotectants (Figure 3c) [25]. It has been shown that the accumulation of total sugars in microalgae can be enhanced under heavy metal stress [26]. Kafil et al. reported that microalgae accumulate total sugars under heavy metal stress, which is a mechanism to combat heavy metal stress [17]. The stress mechanism may be that microalgae produce total sugars in the heavy metal environment and the newly formed total sugars bind to heavy metals in the culture environment on the cell wall of microalgae to reduce the damage of heavy metals to microalgae. The possible reason for the decrease in total sugar is that the degradation of total sugar caused by high concentrations of Cr(III) and Cr(VI) provides nutrients for algal cell growth under high Cr stress.
4.3. Chlorella pyrenoidosa of Cr(III) and Cr(VI) Stress Response
Antioxidant enzymes (SOD, CAT and POD): some external environmental factors of stress such as heavy metals, organic acid and salt, can induce algal cells to form the ROS (Figure 4). Intracellular ROS such as hydroxyl radical (OH−), hydrogen peroxide (H2O2), peroxide(O2−) and singlet oxygen (1O2) have strong oxidation potential and can cause fatal damage to algal cells [27]. SOD can convert O2− to H2O2 and O2, while CAT and POD can convert H2O2 to O2 and H2O. SOD, CAT and POD are frequently utilized as biomarkers to measure cell response to external stress [28]. Therefore, this chapter focuses on the changes in SOD, CAT and POD contents in Chlorella pyrenoidosa cells under Cr(III) and Cr(VI) stress. Relevant studies have confirmed that a low concentration of Cr(VI) will increase the SOD and CAT activities of C. vulgaris, while high concentration of Cr(VI) will decrease the SOD and CAT activities of microalgae [13]. It can be seen from Figure 4 that the addition of Cr(III) and Cr(VI) at different concentrations has significant effects on the antioxidant activity of Chlorella pyrenoidosa. The reason for these results is that when microalgae cells are exposed to low concentrations of Cr, they protect themselves from excessive ROS by increasing the activity of antioxidant enzymes, thereby preventing or reducing oxidative damage. When Cr concentrations exceed the threshold, the antioxidant enzyme system cannot maintain a dynamic balance of free radical production and clearance and the intracellular antioxidant enzyme system produces certain oxidative damage, leading to a decrease in antioxidant capacity.
Non-enzymatic antioxidants (GSH and AsA): to mitigate the toxic effects of ROS, plant cells have developed different defense mechanisms. The initial reaction of oxidative stress is related to the change in the glutathione-ascorbate cycle and catalase activity, which is responsible for the degradation of H2O2 [29]. Branca et al. observed that in the GSH-Cr(VI) system, glutathione reacts with chromate to produce a thioester compound. This thioester substance can be efficiently reduced by free tripeptide molecules. In systems that contain chromates, sugars and glutathione, Cr(VI) interacts with a sugar ligand (or both sugar and GSH) to form an ester that can be easily reduced by GSH. The resulting Cr(V) ion is subsequently stabilized by coordination with the sugar ligand [30]. Therefore, this chapter focuses on the changes in GSH and AsA contents of Chlorella pyrenoidosa in Cr(III) and Cr(VI) medium with different concentrations. As shown in Figure 4d, relevant studies have also confirmed that the GSH content of microalgae cultured in the range of 0–40 mg/L Cr(VI) has similar changes [31]. Schiavon et al. reported that the increased GSH content may be related to the reduction of Cr(VI) to Cr(III), which is less toxic [32]. In this experiment, Cr(III) and Cr(VI) act as environmental stressors, causing a significant increase in the GSH (glutathione) content within algal cells. This increase helps to remove excess reactive oxygen species (ROS) from microalgae. However, when the Cr concentration increases to exceed the threshold of microalgae, it will cause damage to GSH, resulting in a decrease in GSH content. As shown in Figure 4e, during oxidative stress, AsA undergoes oxidation to form dehydroascorbic acid (DHA), resulting in a decrease in AsA content particularly under heavy metal stress. Kováčik et al. also found similar results, AsA was lower than the control group in the presence of Hg and Pb and AsA was protected from destruction of metal toxicity by regulating ROS/NO balance C. subellipsoidea [33]. To further understand the influence of AsA on Cr(VI) in Chlorella pyrenoidosa, the reduction rate of Cr(VI) at different ratios of Cr(VI)/AsA was explored (Figure 4f). The findings indicate that the increase in AsA usage is associated with higher rates of Cr(VI) reduction, as observed by Yen et al. who found that Cr(VI) reduction in microalgae may have two synergistic mechanisms: biological mechanism (enzyme pathway) and abiotic mechanism (GSH release) [34]. Yen et al. also observed the average reduction rate of Cr(VI) in microalgae by three treatment methods: live cells, dead cells and cell-broken super-natant (Table 3) [34].
4.4. Distribution of Cr(III) and Cr(VI) in Algal Cells
In Figure 5a, The removal of Cr(III) is made possible by the binding between cell wall functional groups and metal ions. Moreno et al. discovered that Cr(III) was found when Tetradesmus sp. and Scenedesmus sp. were treated together, and Ascomycota sp., microalgae exhibited irregular and porous cell walls. The interaction between the functional groups of the cell wall and Cr(III) changed the morphological characteristics of microalgae, enhancing the adsorption energy of Cr(III) [35]. For Cr(VI), Lu et al. pointed out that when the concentration of Cr(VI) exceeds a certain limit, the growth of Chlorella vulgaris is significantly inhibited, resulting in the inhibition of Cr removal [16].
According to Figure 5b, The active compounds found in microalgae transform Cr(VI) into Cr(III) partially, thus reducing the toxicity of Cr(VI). Shen et al. pointed out that the decrease in Cr(VI) toxicity is related to the growth media that induce microalgae growth (such as organic carbon, nitrogen and phosphorus) and different types of organic carbon sources can cause different distributions of Cr(VI) and Cr(III) content in microalgae cells [6]. Cr(VI) can react with plant chelating elements and ATP synthase to form hydrogen and ion bonds. Plant chelating elements are rich in cysteine, which can chelate metal ions through thiol (-SH) to reduce the toxicity of Cr(VI). In addition, after entering the cell, Cr(VI) not only binds to macromolecules such asphytochelin, metallothionein and chlorophyll, but also binds to enzymes in the cell, serving as an important detoxification mechanism [36]. The active substance converting Cr(VI) into Cr(III) in algae cells may be ascorbic acid (Figure 4e,f).
4.5. Effect of Cr (III) and Cr (VI) on Activity of ACC, ME and PEPC
In Figure 6a, compared with the control group, the ACC activity of Chlorella pyrenoidosa first increases and then decreases with the concentration of Cr, and is inconsistent results in lipid content of Chlorella pyrenoidosa, this is due to the fact that ACC activity is regulated not only at the transcriptional level, but also after transcription [37,38,39]. However, Cr induces an overproduction of reactive oxygen species, which leads to a decrease in conversion efficiency of light energy and causes damage to lipids, proteins and other macromolecules [40,41,42]. This ultimately results in a decrease in lipid content and an increase in ACC activity.
Ikaran et al. reported that under nitrogen stress, the ME gene of Chlorella vulgaris was significantly upregulated and the total lipid content was increased (Figure 6b) [37]. Ren et al. reported that adding malic acid promotes Schizochytrium sp. to enhance NADPH supply and increase lipid content [43]. Increased ME activity results in a notable rise in the concentration of malate and citrate within the cell, causing an increase in their outflow as well. This increase in concentration enhances the catalytic process of converting malate into pyruvate, which can then be transformed into ACC through the action of pyruvate dehydrogenase [44]. These results suggest that ME may be responsible for both, providing reducing capacities for fatty acid synthesis and ACC under Cr stress, promoting lipid accumulation.
Previous studies have shown that the decrease in PEPC activity is beneficial to fatty acid synthesis (Figure 6c) [45] which is corroborated by the lipid content in Figure 3a. This is because downregulation of the PEPC gene increases the availability of phosphoenolpyruvate and converts it into pyruvate, which is catalyzed by pyruvate dehydrogenase to generate ACC and promote lipid synthesis [37]. Therefore, the low concentration of Cr can promote the synthesis of lipids, the lipid content is 50.43 wt% and 44.95 wt% (0.5 mg/L and 1.0 mg/L Cr(III)), respectively, and Cr (VI) concentrations are 0.5 mg/L and 1.0 mg/L Cr(VI) with lipid content of 38.00 wt% and 37.15 wt%, respectively. The maximum value of PEPC activity in algal cells was achieved at concentrations of 2.0 mg/L Cr(III) and 5.0 mg/L Cr(VI), which were 1.53 and 1.22 times higher than the control group, respectively. This cannot be explained by the demand for carbon in the TCA cycle. Other studies have shown that Chlamydomonas reinhardtii increases PEPC when subjected to nitrogen stress [46,47]. This may be an active molecular mechanism by which algal cells respond to heavy metal stress, resulting in different PEPC activities in Cr(III) and Cr(VI).
4.6. Mechanism of Influence of Cr(III) and Cr(VI) on Lipid Synthesis of Chlorella pyrenoidosa
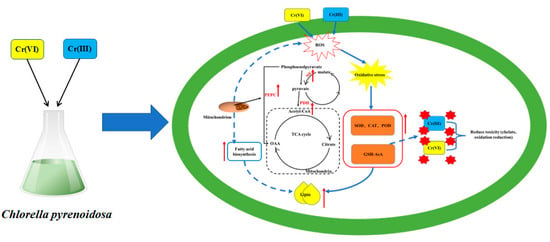
Based on the above results, we can speculate the mechanism of detoxification and lipid production of Cr(III) and Cr(VI) by Chlorella pyrenoidosa (Figure 7). Algal cells are stimulated by Cr(III) and Cr(VI) to produce reactive oxygen species. To mitigate cell damage from oxidative stress and stimulate immune responses, Cr(III) and Cr(VI) engage in a complex chelation reaction with Cr(III) and Cr(VI), promoting the upregulation of antioxidant enzymes (SOD, CAT and POD) as well as non-enzymatic antioxidants (GSH and AsA). Lipid synthesis is stimulated while the TCA cycle in mitochondria is activated to produce secondary metabolites (such as ACC, ME and PEPC) that promote lipid synthesis.
5. Conclusions
Chlorella pyrenoidosa was cultured with Cr(III) and Cr(VI) at 0–5.0 mg/L. The analysis focused on the impact of Cr(III) and Cr(VI) on the lipid content, biomass, protein, total sugars, non-enzymatic antioxidants and enzyme activities (SOD, CAT, POD, ACC, ME and PEPC) of Chlorella pyrenoidosa. The growth and lipid synthesis of Chlorella pyrenoidosa were examined in response to Cr(III) and Cr(VI). The following are the main conclusions.
- (1)
- When Cr(VI) concentration was in the range of 1.0–2.0 mg/L, it was beneficial to accumulate total sugar (37.28–48.33 mg/g FW > 32.83 mg/g FW in the control group). Concentrations of Cr(III) and Cr(VI) ranging from 0.5 to 1.0 mg/L were found to be conducive to lipid accumulation, with a weight percentage of 37.15–50.43 wt%, compared to the control group with 36.87 wt%. At a concentration of 0.5 mg/L of Cr(III), protein accumulation was observed to be higher (13.60 mg/g FW) compared to the control group (12.47 mg/g FW), indicating a beneficial effect.
- (2)
- Cr(VI) is 100 times more toxic than Cr(III), is easy to dissolve and has a strong oxidation capacity, changing the expression of genes and proteins. Microalgae cope with Cr(VI) stress by regulating the activity of antioxidant enzymes, such as increasing the activities of SOD, CAT and POD. Additionally, algal cells actively convert the more toxic Cr(VI) to Cr(III) through substances like GSH and AsA.
- (3)
- Cr(III) and Cr(VI) can help Chlorella pyrenoidosa improve the key enzyme of lipid synthesis in microalgae, but only under the conditions of 0.5–1.0 mg/L Cr(III) and Cr(VI), the lipid level of Chlorella pyrenoidosa is higher than that of the control group. Although the high Cr environment can help the key enzymes of lipid synthesis in microalgae maintain high activity to produce more lipids, the excessive production of reactive oxygen species induced by high Cr will also accelerate the destruction of the synthesized lipids and other macromolecules, which is unfavorable to the final result.
Author Contributions
Methodology, investigation, validation, writing—original draft preparation, T.L. and H.G.; writing—review and editing, Q.Y. and H.L.; supervision, writing—review and editing, funding acquisition, Y.W. (Yajun Wang), C.L., Y.Z. and J.L.; conceptualization, methodology, validation, supervision, writing—review and editing, funding acquisition, Y.W. (Yitong Wang). All authors have read and agreed to the published version of the manuscript.
Funding
The authors wish to acknowledge the financial support from National Natural Science Foundation of China (Nos: 52004095 and 52274333), Outstanding Youth Science Fund Project of Hebei Natural Science Foundation (E2022209019), Innovation Research Group Project of Natural Science Foundation of Hebei Province (E2022209093), Central Guidance Local Science and Technology Development Foundation of Hebei Provincial Department of Science and Technology (236Z3802G) and Key Scientific Research Project of North China University of Science and Technology (ZD-ST-202311-23).
Data Availability Statement
Data are contained within the article.
Acknowledgments
The authors give thanks to the anonymous reviewers and all the editors in the process of manuscript revision.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hedayatkhah, A.; Cretoiu, M.S.; Emtiazi, G.; Stal, L.J.; Bolhuis, H. Bioremediation of chromium contaminated water by diatoms with concomitant lipid accumulation for biofuel production. J. Environ. Manag. 2018, 227, 313–320. [Google Scholar] [CrossRef]
- Monga, A.; Fulke, A.B.; Dasgupta, D. Recent developments in essentiality of trivalent chromium and toxicity of hexavalent chromium: Implications on human health and remediation strategies. J. Hazard. Mater. Adv. 2022, 7, 100113. [Google Scholar] [CrossRef]
- Sarker, S.S.; Akter, T.; Parveen, S.; Uddin, M.T.; Mondal, A.K.; Sujan, S.A. Microalgae-based green approach for effective chromium removal from tannery effluent: A review. Arab. J. Chem. 2023, 16, 105085. [Google Scholar] [CrossRef]
- Naz, A.; Chowdhury, A.; Chandra, R.; Mishra, B.K. Potential human health hazard due to bioavailable heavy metal exposure via consumption of plants with ethnobotanical usage at the largest chromite mine of India. Environ. Geochem. Health 2020, 42, 4213–4231. [Google Scholar] [CrossRef]
- Luo, L.; Yang, C.; Jiang, X.; Guo, W.; Ngo, H.H.; Wang, X.C. Impacts of fulvic acid and Cr (VI) on metabolism and chromium removal pathways of green microalgae. J. Hazard. Mater. 2023, 459, 132171. [Google Scholar] [CrossRef]
- Shen, L.; Saky, S.A.; Yang, Z.; Ho, S.H.; Chen, C.; Qin, L.; Lu, Y. The critical utilization of active heterotrophic microalgae for bioremoval of Cr (VI) in organics co-contaminated wastewater. Chemosphere 2019, 228, 536–544. [Google Scholar] [CrossRef]
- Prasad, S.; Yadav, K.K.; Kumar, S.; Gupta, N.; Cabral-Pinto, M.M.; Rezania, S.; Alam, J. Chromium contamination and effect on environmental health and its remediation: A sustainable approaches. J. Environ. Manag. 2021, 285, 112174. [Google Scholar] [CrossRef]
- de Moura Sousa, L.; Moreira, F.S.; Cardoso, V.L.; Batista, F.R.X. Light intensity effect on the performance of Rhodobacter capsulatus in removal of chromium from effluent. J. Water Process. 2023, 52, 103567. [Google Scholar] [CrossRef]
- Congeevaram, S.; Dhanarani, S.; Park, J.; Dexilin, M.; Thamaraiselvi, K. Biosorption of chromium and nickel by heavy metal resistant fungal and bacterial isolates. J. Hazard. Mater. 2007, 146, 270–277. [Google Scholar] [CrossRef]
- Lanka, S.; Murari, S.G. Aquatic Plants in Phytoextraction of Hexavalent Chromium and Other Metals from Electroplating Effluents. In Advances in Bioremediation and Phytoremediation for Sustainable Soil Management: Principles, Monitoring and Remediation; Springer: Berlin/Heidelberg, Germany, 2022; pp. 129–139. [Google Scholar]
- Devi, A.; Verma, M.; Saratale, G.D.; Saratale, R.G.; Ferreira, L.F.R.; Mulla, S.I.; Bharagava, R.N. Microalgae: A green eco-friendly agents for bioremediation of tannery wastewater with simultaneous production of value-added products. Chemosphere 2023, 336, 139192. [Google Scholar] [CrossRef]
- Sharma, A.; Kapoor, D.; Wang, J.; Shahzad, B.; Kumar, V.; Bali, A.S.; Yan, D. Chromium bioaccumulation and its impacts on plants: An overview. Plants 2020, 9, 100. [Google Scholar] [CrossRef]
- Aharchaou, I.; Rosabal, M.; Liu, F.; Battaglia, E.; Vignati, D.A.; Fortin, C. Bioaccumulation and subcellular partitioning of Cr (III) and Cr (VI) in the freshwater green alga Chlamydomonas reinhardtii. Aquat. Toxicol. 2017, 182, 49–57. [Google Scholar] [CrossRef]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Abbas, F.; Bukhari, S.A.H.; Wu, L. Citric acid assisted phytoextraction of chromium by sunflower; morpho-physiological and biochemical alterations in plants. Ecotoxicol. Environ. Saf. 2017, 145, 90–102. [Google Scholar] [CrossRef]
- Daneshvar, E.; Zarrinmehr, M.J.; Kousha, M.; Hasht, A.M.; Saratale, G.D.; Maiti, A.; Bhatnagar, A. Hexavalent chromium removal from water by microalgal-based materials: Adsorption, desorption and recovery studies. Bioresour. Technol. 2019, 293, 122064. [Google Scholar] [CrossRef]
- Lu, M.M.; Gao, F.; Li, C.; Yang, H.L. Response of microalgae Chlorella vulgaris to Cr stress and continuous Cr removal in a membrane photobioreactor. Chemosphere 2021, 262, 128422. [Google Scholar] [CrossRef]
- Kafil, M.; Berninger, F.; Koutra, E.; Kornaros, M. Utilization of the microalga Scenedesmus quadricauda for hexavalent chromium bioremediation and biodiesel production. Bioresour. Technol. 2022, 346, 126665. [Google Scholar] [CrossRef]
- Fozer, D.; Kiss, B.; Lorincz, L.; Szekely, E.; Mizsey, P.; Nemeth, A. Improvement of microalgae biomass productivity and subsequent biogas yield of hydrothermal gasification via optimization of illumination. Renew. Energy 2019, 138, 1262–1272. [Google Scholar] [CrossRef]
- Cervantes, C.; Campos-García, J.; Devars, S.; Gutiérrez-Corona, F.; Loza-Tavera, H.; Torres-Guzmán, J.C.; Moreno-Sánchez, R. Interactions of chromium with microorganisms and plants. FEMS Microbiol. Rev. 2001, 25, 335–347. [Google Scholar] [CrossRef]
- Ji, X.; Cheng, J.; Gong, D.; Zhao, X.; Qi, Y.; Su, Y.; Ma, W. The effect of NaCl stress on photosynthetic efficiency and lipid production in freshwater microalga—Scenedesmus obliquus XJ002. Sci. Total Environ. 2018, 633, 593–599. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; Maurya, R.; Trivedi, K.; Patidar, S.K.; Ghosh, A.; Mishra, S. Salinity induced oxidative stress enhanced biofuel production potential of microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 189, 341–348. [Google Scholar] [CrossRef]
- Sharma, J.; Kumar, V.; Kumar, S.S.; Malyan, S.K.; Mathimani, T.; Bishnoi, N.R.; Pugazhendhi, A. Microalgal consortia for municipal wastewater treatment–Lipid augmentation and fatty acid profiling for biodiesel production. J. Photochem. 2020, 202, 111638. [Google Scholar] [CrossRef]
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: A review. Energies 2013, 6, 4607–4638. [Google Scholar] [CrossRef]
- Sun, X.; Zhong, Y.; Huang, Z.; Yang, Y. Selenium accumulation in unicellular green alga Chlorella vulgaris and its effects on antioxidant enzymes and content of photosynthetic pigments. PLoS ONE 2014, 9, e112270. [Google Scholar] [CrossRef]
- Markou, G.; Angelidaki, I.; Georgakakis, D. Microalgal carbohydrates: An overview of the factors influencing carbohydrates production, and of main bioconversion technologies for production of biofuels. Appl. Microbiol. Biotechnol. 2012, 96, 631–645. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive oxygen species: Response of algal cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Cheng, J.; Ye, Q.; Yang, Z.; Yang, W.; Zhou, J.; Cen, K. Microstructure and antioxidative capacity of the microalgae mutant Chlorella PY-ZU1 during tilmicosin removal from wastewater under 15% CO2. J. Hazard. Mater. 2017, 324, 414–419. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Branca, M.; Micera, G.; Kozlowski, H.; Swiatek, J. Reduction of chromate ions by glutathione tripeptide in the presence of sugar ligands. J. Inorg. 1990, 39, 217–226. [Google Scholar] [CrossRef]
- Song, X.; Liu, B.F.; Kong, F.; Song, Q.; Ren, N.Q.; Ren, H.Y. Simultaneous chromium removal and lipid accumulation by microalgae under acidic and low temperature conditions for promising biodiesel production. Bioresour. Technol. 2023, 370, 128515. [Google Scholar] [CrossRef]
- Schiavon, M.; Galla, G.; Wirtz, M.; Pilon-Smits, E.A.; Telatin, V.; Quaggiotti, S.; Malagoli, M. Transcriptome profiling of genes differentially modulated by sulfur and chromium identifies potential targets for phytoremediation and reveals a complex S–Cr interplay on sulfate transport regulation in B. juncea. J. Hazard. Mater. 2012, 239, 192–205. [Google Scholar] [CrossRef]
- Kováčik, J.; Rotková, G.; Bujdoš, M.; Babula, P.; Peterková, V.; Matúš, P. Ascorbic acid protects Coccomyxa subellipsoidea against metal toxicity through modulation of ROS/NO balance and metal uptake. J. Hazard. Mater. 2017, 339, 200–207. [Google Scholar] [CrossRef]
- Yen, H.W.; Chen, P.W.; Hsu, C.Y.; Lee, L. The use of autotrophic Chlorella vulgaris in chromium (VI) reduction under different reduction conditions. J. Taiwan Inst. Chem. Eng. 2017, 74, 1–6. [Google Scholar] [CrossRef]
- Moreno-García, A.F.; Neri-Torres, E.E.; Mena-Cervantes, V.Y.; Altamirano, R.H.; Pineda-Flores, G.; Luna-Sánchez, R.; Suastes-Rivas, J.K. Sustainable biorefinery associated with wastewater treatment of Cr (III) using a native microalgae consortium. Fuel 2021, 290, 119040. [Google Scholar] [CrossRef]
- Li, N.; Qin, L.; Zhang, L.; Geng, W. Extracellular adsorption, intracellular accumulation and tolerance mechanisms of Cyclotella sp. to Cr (VI) stress. Chemosphere 2021, 270, 128662. [Google Scholar] [CrossRef]
- Ikaran, Z.; Suárez-Alvarez, S.; Urreta, I.; Castañón, S. The effect of nitrogen limitation on the physiology and metabolism of Chlorella vulgaris var L3. Algal Res. 2015, 10, 134–144. [Google Scholar] [CrossRef]
- Sasaki, Y.; Nagano, Y. Plant acetyl-CoA carboxylase: Structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Biotechnol. Biochem. 2004, 68, 1175–1184. [Google Scholar] [CrossRef]
- Ke, J.; Wen, T.N.; Nikolau, B.J.; Wurtele, E.S. Coordinate regulation of the nuclear and plastidic genes coding for the subunits of the heteromeric acetyl-coenzyme A carboxylase. Plant Physiol. 2000, 122, 1057–1072. [Google Scholar] [CrossRef]
- Zhao, Y.; Qiao, T.; Han, B.; Dong, X.; Geng, S.; Ning, D.; Yu, X. Simultaneous enhancement of biomass and lipid production of Monoraphidium sp. QLZ-3 in a photobioreactor by using walnut shell extracts. Energy Convers. 2020, 204, 112326. [Google Scholar] [CrossRef]
- Yang, J.; Cao, J.; Yuan, H. Lipid production combined with biosorption and bioaccumulation of cadmium, copper, manganese and zinc by oleaginous microalgae Chlorella minutissima UTEX2341. Bioresour. Technol. 2015, 175, 537–544. [Google Scholar] [CrossRef]
- Han, W.; Ding, W.; Lu, S.; Song, K.; Chen, C.; Zhou, X. Effects of nutrient composition, lighting conditions, and metal ions on the growth and lipid yield of the high-lipid-yielding microalgae (Chlorella pyrenoidosa) cultivated in municipal wastewater. J. Environ. 2021, 9, 106491. [Google Scholar] [CrossRef]
- Ren, L.J.; Huang, H.; Lian, M.; Ji, X.J. Enhanced docosahexaenoic acid production by reinforcing acetyl-CoA and NADPH supply in Schizochytrium sp. HX-308. Bioprocess. Biosyst. Eng. 2009, 32, 837–843. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, B.; Ding, H.; Zhang, J.; Li, S. The role of NADP-malic enzyme in plants under stress. Plant Sci. 2019, 281, 206–212. [Google Scholar] [CrossRef]
- Seo, S.; Jeon, H.; Chang, K.S. Enhanced biomass production by Phaeodactylum tricornutum overexpressing phosphoenolpyruvate carboxylase. Algal Res. 2018, 31, 489–496. [Google Scholar] [CrossRef]
- Mamedov, T.G.; Moellering, E.R.; Chollet, R. Identification and expression analysis of two inorganic C-and N-responsive genes encoding novel and distinct molecular forms of eukaryotic phosphoenolpyruvate carboxylase in the green microalga Chlamydomonas reinhardtii. Plant J. 2005, 42, 832–843. [Google Scholar] [CrossRef]
- Moellering, E.R.; Ouyang, Y.; Mamedov, T.G.; Chollet, R. The two divergent PEP-carboxylase catalytic subunits in the green microalga Chlamydomonas reinhardtii respond reversibly to inorganic-N supply and co-exist in the high-molecular-mass, hetero-oligomeric Class-2 PEPC complex. FEBS Lett. 2007, 581, 4871–4876. [Google Scholar] [CrossRef]
Figure 1.
Absorbance of Cr(VI) at 540 nm.

Figure 2.
Effects of Cr(III) and Cr(VI) on biomass of Chlorella pyrenoidosa.

Figure 3.
Effects of Cr(III) and Cr(VI) on (a) lipid content, (b) protein content and (c) total sugar content of Chlorella pyrenoidosa. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).
Figure 3.
Effects of Cr(III) and Cr(VI) on (a) lipid content, (b) protein content and (c) total sugar content of Chlorella pyrenoidosa. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).

Figure 4.
Response of Chlorella pyrenoidosa to Cr(III) and Cr(VI) stress and effect of AsA on Cr(VI) reduction rate. (a) SOD, (b) CAT, (c) POD, (d) GSH, (e) ASA and (f) Cr(VI) reduction rate. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).
Figure 4.
Response of Chlorella pyrenoidosa to Cr(III) and Cr(VI) stress and effect of AsA on Cr(VI) reduction rate. (a) SOD, (b) CAT, (c) POD, (d) GSH, (e) ASA and (f) Cr(VI) reduction rate. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).

Figure 5.
Removal of (a) total Cr and distribution of (b) Cr(III) and Cr(VI) contents in algal cells.
Figure 5.
Removal of (a) total Cr and distribution of (b) Cr(III) and Cr(VI) contents in algal cells.

Figure 6.
Effects of Cr(III) and Cr(VI) on (a) ACC, (b) ME and (c) PEPC activities. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).
Figure 6.
Effects of Cr(III) and Cr(VI) on (a) ACC, (b) ME and (c) PEPC activities. The different letters are used to indicate the difference results after significant difference analysis (p < 0.05).

Figure 7.
Mechanism of Cr(III) and Cr(VI) on lipid synthesis of Chlorella pyrenoidosa.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Chemical reagents used in study.
| Reagents | Chemical Formula | Purity | Manufacturer | Location |
|---|---|---|---|---|
| Chromium trichloride hexahydrate | CrCl3·6H2O | ≥98.00% | Aladdin Reagent Co., Ltd. | Shanghai, China |
| Copper sulfate pentahydrate | CuSO4·5H2O | ≥99.00% | ||
| Sodium carbonate | Na2CO3 | ≥99.50% | ||
| Dipotassium phosphate | K2HPO4 | ≥99.00% | ||
| Ammonium ferric citrate | C6H11FeNO7 | ≥99.00%, Fe: 20.50–22.50% | ||
| Disodium dihydrate ethylenediamine tetraacetate | C10H14N2Na2O8·2H2O | ≥98.00% | ||
| Boric acid | H3BO3 | ≥99.00% | ||
| Zinc sulfate heptahydrate | ZnSO4·7H2O | ≥99.00% | ||
| Magnesium sulfate heptahydrate | MgSO4·7H2O | ≥99.00% | ||
| Sodium molybdate dihydrate | Na2MoO4·2H2O | ≥99.00% | ||
| Calcium chloride dihydrate | CaCl2·2H2O | ≥99.90% | Macklin Biochemical Technology Co., Ltd. | Shanghai, China |
| Manganese chloride tetrahydrate | MnCl2·4H2O | ≥99.00% | ||
| Cobalt nitrate hexahydrate | Co(NO3)2·6H2O | ≥99.00% | ||
| Sodium nitrate | NaNO3 | ≥99.00% | SHENTAI Chemical Industry Co., Ltd. | Tianjin, China |
| Potassium dichromate | K2Cr2O7 | ≥99.80% |
Table 2.
Compositions of BG11 medium used in study, (g/1 L ultrapure water).
| Compositions | NaNO3 | K2HPO4 | MgSO4 7H2O | C6H8O7 | C6H11FeNO7 | Na2EDTA | CaCl2 2H2O | Na2CO3 | H3BO3 | MnCl4 4H2O | ZnSO4 7H2O | Na2MoO4 2H2O | CuSO4 5H2O | Co(NO3)2 6H2O |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Content | 1.5 | 0.04 | 0.75 | 0.006 | 0.006 | 0.001 | 0.036 | 0.02 | 0.00286 | 0.00182 | 0.00022 | 0.0004 | 0.00008 | 0.0005 |
Table 3.
Average reduction rate of Cr(VI) by different treatment methods [34].
Table 3.
Average reduction rate of Cr(VI) by different treatment methods [34].
| Living Cell | Dead Cell | Cell Crushing Supernatant | |
|---|---|---|---|
| Decrement rate (mg/L/day) | 3.73 | 1.52 | 0.38 |
| Specific reduction rate (mg/g/day) | 2.07 | 0.95 | - |
Note: Performed at an initial concentration of 10 mg/L Cr(VI).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, T.; Guo, H.; Yu, Q.; Wang, Y.; Liu, H.; Zeng, Y.; Wang, Y.; Liu, C.; Li, J. The Impact of Cr(III) and Cr(VI) on Lipid Accumulation in Chlorella pyrenoidosa. Processes 2024, 12, 905. https://doi.org/10.3390/pr12050905
AMA Style
Liu T, Guo H, Yu Q, Wang Y, Liu H, Zeng Y, Wang Y, Liu C, Li J. The Impact of Cr(III) and Cr(VI) on Lipid Accumulation in Chlorella pyrenoidosa. Processes. 2024; 12(5):905. https://doi.org/10.3390/pr12050905
Chicago/Turabian StyleLiu, Tianji, Huawei Guo, Qing Yu, Yajun Wang, Huan Liu, Yanan Zeng, Yitong Wang, Chunyu Liu, and Junguo Li. 2024. "The Impact of Cr(III) and Cr(VI) on Lipid Accumulation in Chlorella pyrenoidosa" Processes 12, no. 5: 905. https://doi.org/10.3390/pr12050905
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.