1. Introduction
Improvements in food production that both utilize wastes efficiently and support local economies to improve circularity are needed to feed a growing population [
1]. This is especially true given the current shift in population density towards urban centers [
2] that provides opportunities and challenges for food production. Urban centers that are surrounded by agricultural production are in a unique situation where small growing operations have the opportunity to use composted local municipal food and/or local manure wastes to produce food more sustainably.
Between 2010 and 2020, the population within the United States increased by 7.3%, but in Florida the population increased almost twice as fast, at a rate of 14.5% [
3]. The number of counties that were reclassified as urban from rural has increased by 15.8% between 2013 and 2021 in the state, with 65.7% of Florida counties now classified as urban [
4]. Furthermore, over 1.68 million head of beef and dairy cattle were raised in Florida in 2021 [
5], generating large quantities of manures that are often composted prior to utilization. As a state that experiences both an increase in municipal food waste generation due to rapid urbanization and the continued production of large quantities of livestock waste, Florida is an ideal location to compare how different compost feedstocks affect soil fertility and crop nutrition.
Compost can provide multiple benefits in agroecosystems, such as increasing soil organic matter (SOM), water holding capacity, and concentration of soil nutrients [
6]. Using composted instead of raw organic materials has additional benefits, including the reduction of viable weed seeds in the field after application, increased nutrient concentrations, and lower transport costs due to reduced moisture and carbon content in the finished product [
7,
8]. Compost can be created from a variety of feedstocks, with manure-based compost primarily used in rural commercial operations and food- or manure-based compost used in urban and small-scale operations. Large farms typically include fertilizers with compost to increase yields [
9], but smaller operations are more likely to use composts as a sole source of nutrients due to availability and cost-effectiveness [
10]. The utilization of food-based compost has the additional benefit of diverting organic waste from landfills, which can help reduce greenhouse gas emissions (e.g., methane) and conserve landfill space.
Most producers that use compost as their sole nutrient source apply composts based on plant nitrogen (N) requirements, although satisfying crop N requirements with composts typically requires a high rate of application due to low N concentrations in most composts [
11]. Synchronizing the mineralization of compost N with crop N needs is necessary to achieve maximum yields and avoid N losses, but this can be challenging as microbial activity controls the quantity and timing of inorganic plant-available nitrogen (PAN) released from compost after application. Composts that have a feedstock with a carbon to N (C:N) ratio above 20:1 can result in N immobilization and a reduction in PAN [
12,
13], although the release or immobilization of PAN can be variable for a given C:N value. Furthermore, as nitrate is prone to losses via leaching and denitrification, the effects of different composts on the nitrification rate of the ammonium produced during mineralization will also affect N cycling and availability.
At increased compost application rates, phosphorus (P) is typically provided in excess of crop P requirement, especially for manure-based composts [
6]. While P is typically lost from most agroecosystems via erosion or runoff, coarse-textured soils and/or soils that have saturated their P sorption capacity can also leach excess P, including several agricultural soils of Florida [
14]. Although P applications can reduce uptake of toxic elements such as cadmium in some crops such as spinach (
Spinacia oleracea L.) [
15], excessive P losses can impair waterways, reduce root length, and inhibit micronutrient uptake (e.g., Fe, Zn) [
16,
17]. Other essential nutrients are also found in compost at varying rates, depending on the feedstock, including potassium (K). Composts with a high level of K may provide fertility benefits in soils with low K concentrations, especially coarse-textured soils [
18]. However, these coarse-textured soils, including many of Florida’s agricultural soils [
14], are also favorable to leaching of monovalent cations such as K. Ultimately, while N is the primary target guiding compost application rates, different composts will affect the availability and cycling of several other nutrients, and these must be taken into consideration to maximize crop productivity and minimize environmental impacts.
A previous laboratory incubation using compost products available in Florida demonstrated that manure-based composts immobilized and decreased soil PAN whereas food-based composts increased PAN relative to unamended soils [
19]. As that previous study did not include crop growth, the objective of this study was to further the research conducted by Kelley et al. [
19] in a greenhouse study designed to evaluate the effects of different compost amendments on spinach growth, nutrient concentration and accumulation, and soil nutrient concentrations. We selected spinach as a model crop as it is a member of an important family (Amaranthaceae, which includes beets and chard) for Florida vegetable growers that operate diversified small-scale operations. Spinach’s fast growth and relatively modest size also make it a good model crop to grow potted in a greenhouse environment, to investigate the effects of different amendments on multiple essential nutrients. Given the past results of Kelley et al. [
19], we hypothesized that food-based composts would result in higher spinach biomass production, nutrient accumulation, and soil nutrient concentrations relative to manure-based composts.
2. Materials and Methods
2.1. Experimental Location and Design
This study was conducted between 3 April and 29 May in 2019 in a temperature-controlled greenhouse located at the University of Florida (UF; Gainesville, FL, USA). Soil amendments were selected to compare manure- and food-based composts of different origins. The experiment was arranged as a randomized complete block design with five blocks and two factors (
Table 1). The first factor included two soils and the second factor included four compost amendments: Black Kow
TM (Black Kow, Oxford, FL, USA), Ecoscraps
TM (EcoScraps Organic Compost, Marysville, OH, USA), composted dairy manure consisting of fibrous solids mechanically separated from flushed dairy manure [
20] (hereafter referred to as ‘Dairy’), and food waste compost from the University of Florida-Institute of Food and Agricultural Sciences (UF-IFAS) Compost Cooperative (hereafter referred to as ‘SCC’). Black Kow is a commercial product derived from cow manure whereas Ecoscraps is a commercially produced compost consisting of a mixture of plant materials, food waste, and processed forest products. The Dairy and SCC composts are produced locally.
All compost amendments were air dried and passed through a 6 mm screen and stored at 4 °C in polyethylene bags for less than a month before mixing with the soil. An unamended negative control of each soil type and a positive control with commercially purchased potting soil (Sta-Green Potting Mix, The Mulch and Soil Company, Ft. Myers, FL, USA) were included in each block. The experiment included a total of 55 pots (diameter: 15.5 cm; height: 16 cm): four treatments and a negative control for each soil in addition to the positive control in each block, replicated in five blocks.
Despite environmental conditions being better controlled in a greenhouse environment relative to field conditions, we decided to set up this experiment using a blocked design, as pots were located on two tables that may not have experienced the same direct light exposure, temperature, and humidity. The inclusion of a block factor allowed us to take into consideration any potential heterogeneity in growing conditions.
2.2. Soil and Compost Processing
Surface soils (0–15 cm) were collected from two sites on the UF campus, as previously described by Kelley et al. [
19]. The first soil was mapped as a Lake sand (Hyperthermic, coated Typic Quartzipsamments) with high C and N for the area, which may be due to prior land use consisting of a grassland with bahiagrass (
Paspalum notatum) and wildflowers for 20 years before cultivation (
Table 2). The second soil was mapped as a Millhopper sand (Loamy, siliceous, semiactive, hyperthermic Grossarenic Paleudults) that had been under intensive agricultural operation for several years, including low organic inputs and frequent tillage. This site is used as a teaching garden for the Horticultural Sciences Department at UF, and a broad variety of vegetables are grown at this location, although the exact vegetable crop grown immediately before soil collection is not known. Soils were collected from multiple areas at each site, homogenized thoroughly, air-dried, and sieved (2 mm) before analysis. All analyses were conducted on a sample that was pooled from several subsamples collected carefully within the whole mass of soil available.
Compost pH (in water), total macronutrient concentration (by digestion and inductively coupled plasma spectrometry) and estimated N availability in the first year after application were analyzed by an external laboratory (Agrolabs, Harrington, DE, USA). Nitrogen availability in the first year after application was estimated by Agrolabs based on ammonium and nitrate concentrations measured (assumed to be all available) and the fraction of organic N that becomes mineralized during the growing season, estimated using equations specific to different compost types that are based on past research and lab experience (William Rohrer, certified professional agronomist and certified crop advisor, Agrolabs; personal communication, 28 June 2022). We measured total C (TC) and total N (TN) by combustion using a Thermo Flash EA, resin-extractable P by colorimetry [
21], N-NH
4+ [
22] and N-NO
3− [
23] by colorimetry, and moisture within labs at UF (
Table 3); additional details of N and P extractions are provided in
Section 2.3. All analyses were conducted on a sample that was pooled from several subsamples collected carefully within each compost bag.
Compost input rates were selected to provide 101 kg PAN ha
−1, the recommended N rate for spinach in Florida [
24], using 50% of the estimated N availability measured by Agrolabs, given the shorter growing season (less than two months) relative to a full year (
Table 4). Amendments were thoroughly mixed with each soil using a shovel and pots were filled with 2 kg of soil plus amendments in a quantity that provided 47 mg N kg
−1 of soil, equivalent to 101 kg PAN ha
−1 for a soil depth of 15 cm at a bulk density of 1.43 g cm
−3. All pots were adjusted to 50% water-filled pore space after planting and drip irrigated until harvest.
2.3. Soil Nutrient Analyses
Resin-extractable P was measured before planting and at harvest whereas PAN was measured before planting and every two weeks after planting, until harvest. For resin-extractable P, fresh soils (2.5 g) were extracted with 35 mL of double-distilled water and a resin strip that was previously charged with NaHCO
3 on a reciprocal shaker for 16 h, then eluted with 0.5 M HCl. For soil PAN, fresh soils (5 g) were extracted using 25 mL of 2M KCl for 30 min on a reciprocal shaker, followed by centrifugation and filtration. Extracted samples were analyzed by colorimetry for resin-extractable P [
21] and PAN [
22,
23] on an Epoch 2 microplate reader (Biotek, Winooski, VT, USA).
2.4. Plant Tissue Analyses
Spinach (cv. ‘Arrowhead’) was seeded at four seeds per pot and thinned to one seed per pot. After eight weeks, all aboveground biomass was separated from roots by clipping the biomass at the soil level; this was performed for each individual pot. Roots were thoroughly washed with deionized water to remove adhering soil. Fresh biomass weight was recorded at harvest and dry weight was recorded after tissue samples were dried to a constant weight in a forced air dryer (65 °C).
Dried spinach aboveground biomass was ground through a 2 mm sieve and analyzed for total Kjeldahl N (TKN), and total P and K via digestion followed by quantification using inductively coupled plasma spectrometry (UF/IFAS Analytical Services Laboratories, Analytical Research Laboratory, Gainesville, FL, USA). Nutrient accumulation in aboveground biomass was computed by multiplying dried biomass with N, P, and K concentrations. Because greenhouse conditions are often not equivalent to field conditions, we did not quantify quality indicators for spinach (e.g., vitamin C, oxalate), and restricted the variables measured to biomass and nutrient concentrations.
2.5. Data Analysis
Soil type and amendment were treated as fixed factors for analyses. Data for PAN and soil P were analyzed using a repeated measures analysis, resulting in an additional fixed factor (time) in the model. All data were tested for normality and homogeneity of variance and log transformed before analysis if needed; only tissue nutrients and soil PAN required a log transformation. All data satisfied analysis of variance (ANOVA) assumptions after transformations. If two- or three-way interactions were observed between soil, treatment, or time, each soil was evaluated separately. Due to poor growth in some treatments of the Millhopper soil, a complete dataset for tissue nutrients was only available for the Lake soil. Thus, each soil was analyzed separately from the onset for nutrient concentrations and accumulation in spinach. Less than 0.02% of the data had residuals ≥ 4 standard deviations, and these were omitted before re-running final analyses. When significant at a p-value ≤ 0.05, means were separated using the Tukey HSD method.
Regressions were computed to evaluate the relationship between spinach shoot dry weight biomass and soil nutrients measured at planting or amendment inputs of C or nutrients. Regressions were also performed between concentrations or accumulation of N, P, and K in harvested spinach tissue and soil N concentrations or amendment N, P or K inputs measured at planting.
All analyses were conducted in SAS using the GLIMMIX or REG procedure (v 9.4; SAS Institute, Cary, NC, USA).
3. Results
3.1. Soil Nutrients
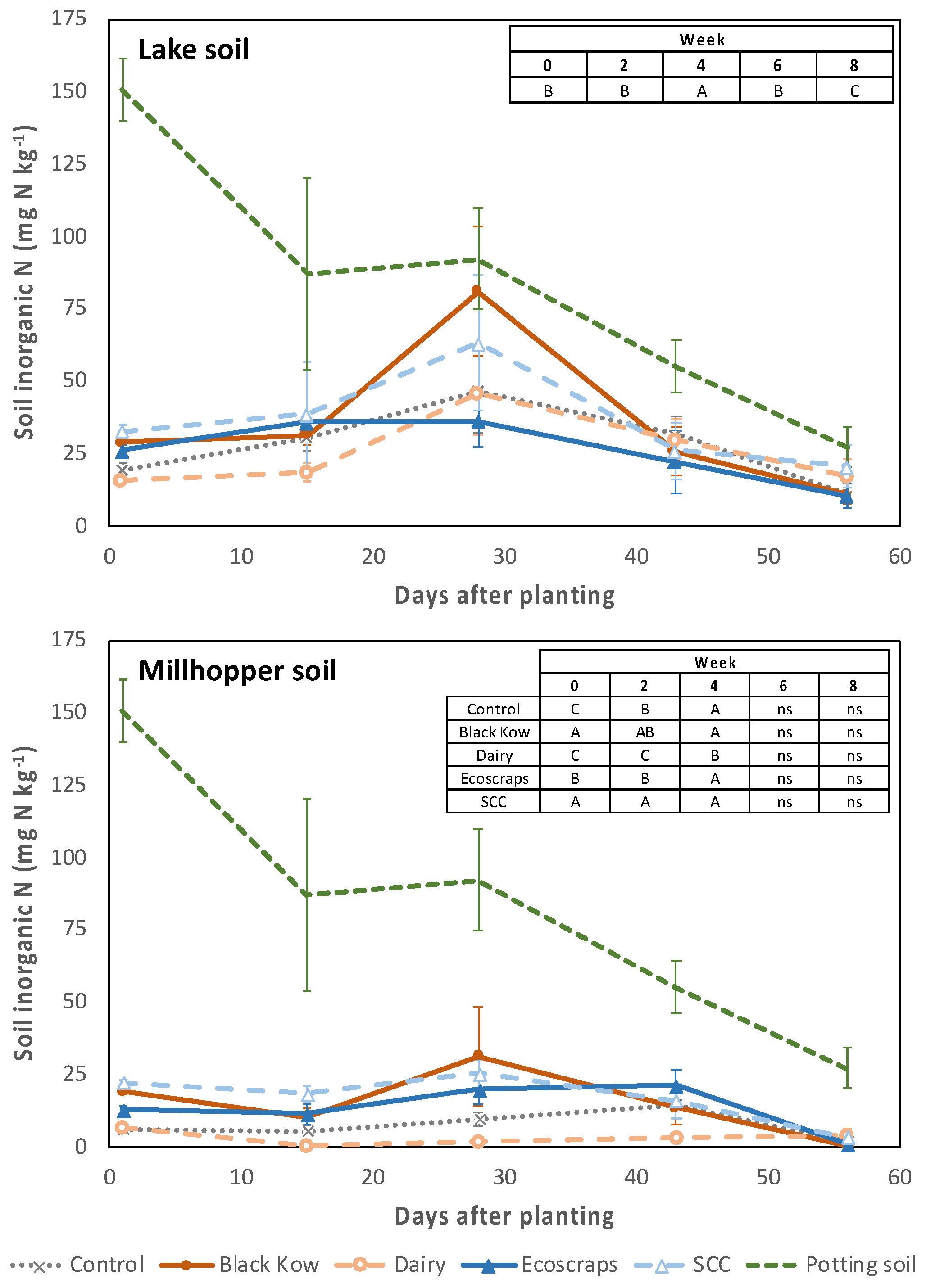
There was a time x treatment interaction for soil PAN in the Millhopper soil (
p < 0.001), but not for the Lake soil (
p = 0.81). During the first week of the study in the Millhopper soil, PAN was highest with SCC and Black Kow (19–22 mg N kg
−1), intermediate with Ecoscraps (13 mg N kg
−1), and lowest in the control and Dairy treatments (6–7 mg N kg
−1;
Figure 1). The concentration of PAN increased in the Ecoscraps and control treatments until week 4, when all treatments were higher (10–31 mg N kg
−1) than Dairy (2 mg N kg
−1). After week 4, PAN decreased for the remainder of the experiment, except for Ecoscraps and the control, and was similar among treatments from week 6 through harvest (0–21 mg N kg
−1). Soil PAN in the Lake soil followed a similar temporal trend (
p < 0.001) with a peak at week 4 (37–81 mg N kg
−1) and the lowest values at harvest (11–21 mg N kg
−1); treatments did not affect soil PAN however (
p = 0.74). In all cases, soil PAN was numerically lower in amended soils than in the potting soil.
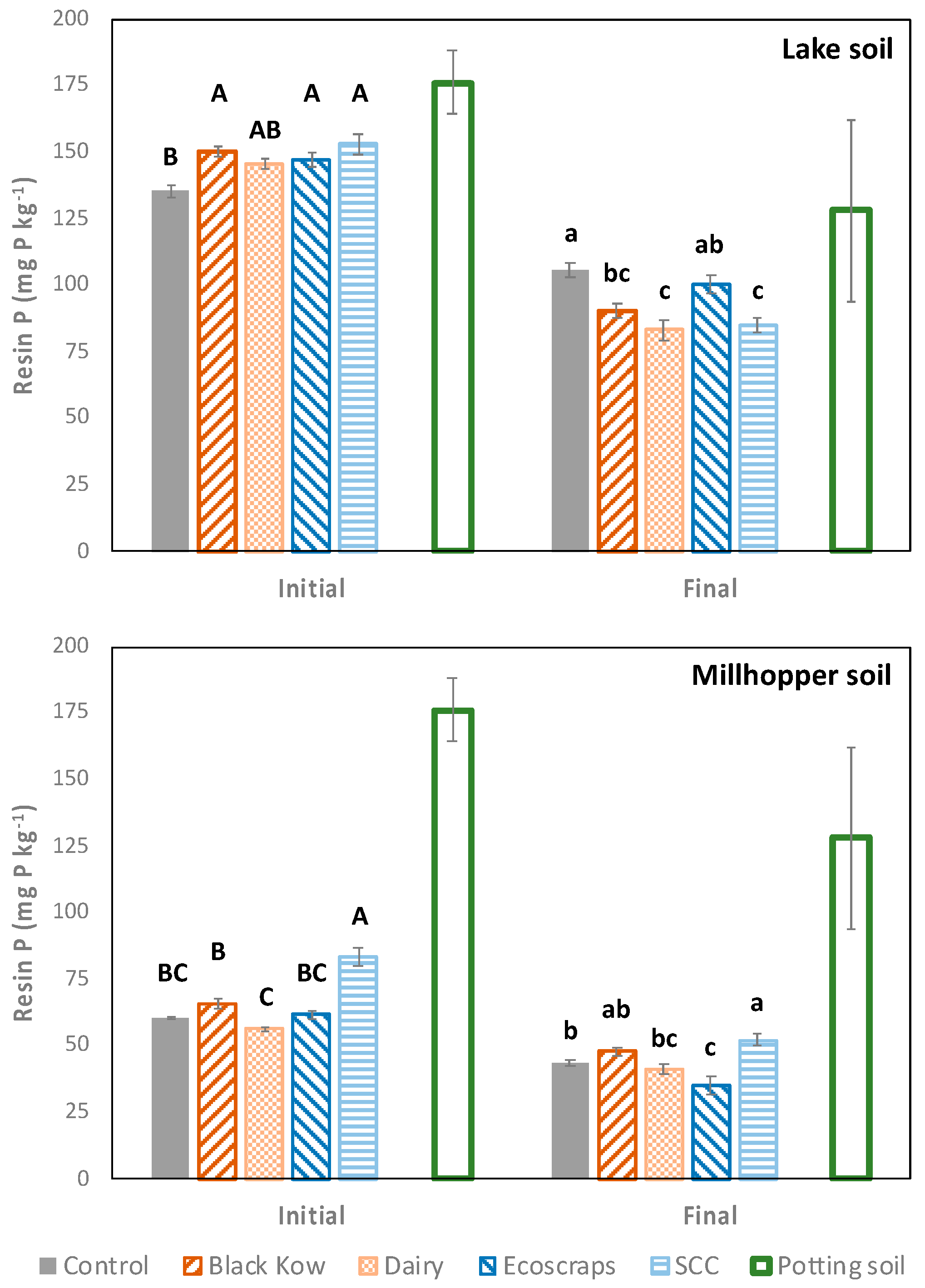
There was a soil x treatment x time interaction (
p < 0.001), in addition to treatment x time and soil x time interactions for all resin P analyses (
p < 0.001 for all combinations). In the Lake soil, soil P at planting was greater in SCC, Ecoscraps, and Black Kow (147–153 mg P kg
−1) compared to the control (135 mg P kg
−1,
Figure 2,
p = 0.002). Soil P decreased for all treatments during the experiment, and the control had a greater concentration of soil P (106 mg P kg
−1) at harvest compared to Black Kow, SCC, and Dairy (83–91 mg P kg
−1;
p < 0.001). For the Millhopper soil, soil P was greater in the SCC treatment (84 mg P kg
−1) relative to the control (61 mg P kg
−1) at planting (
p < 0.001), and all soil P levels were lower at harvest, with SCC (52 mg P kg
−1) and Ecoscraps (35 mg P kg
−1) being higher and lower than the control (43 mg P kg
−1), respectively (
p < 0.001). The potting soil had the highest soil P concentrations for both sampling events.
3.2. Plant Responses
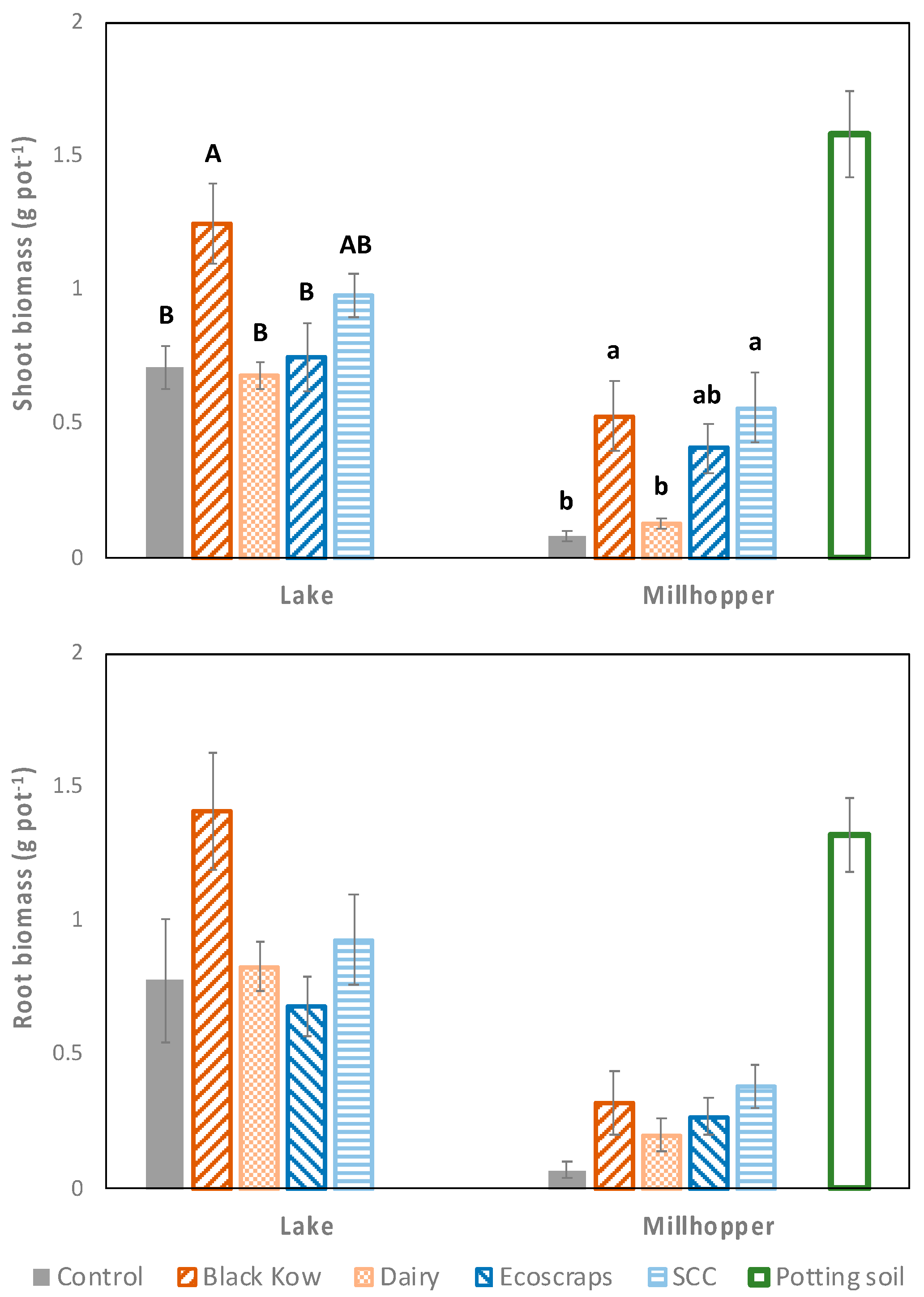
There was no soil x treatment interaction for root biomass (
p = 0.13) or shoot biomass (
p = 0.16). Overall, root and shoot biomass were 375% and 262% greater in the Lake soil compared to the Millhopper soil, respectively (data not shown;
p < 0.003). Shoot biomass was greatest for Black Kow (0.53–1.25 g pot
−1) compared to the control (0.08–0.71 g pot
−1) and Dairy (0.13–0.68 g pot
−1) in both soils (
Figure 3;
p = 0.02). SCC (0.98 g pot
−1) had similar shoot biomass to Black Kow (1.25 g pot
−1) and the control (0.71 g pot
−1) in the Lake soil, but greater shoot biomass (0.56 g pot
−1) than the control (0.08 g pot
−1) in the Millhopper soil. Ecoscraps (0.41 g pot
−1) was similar to Black Kow (0.53 g pot
−1) and SCC (0.56 g pot
−1) in the Millhopper soil only. Root biomass was not affected by treatment in either soil (
p > 0.17 for all analyses). The potting soil had greater shoot biomass than other treatments, and similar root biomass to Black Kow in the Lake soil.
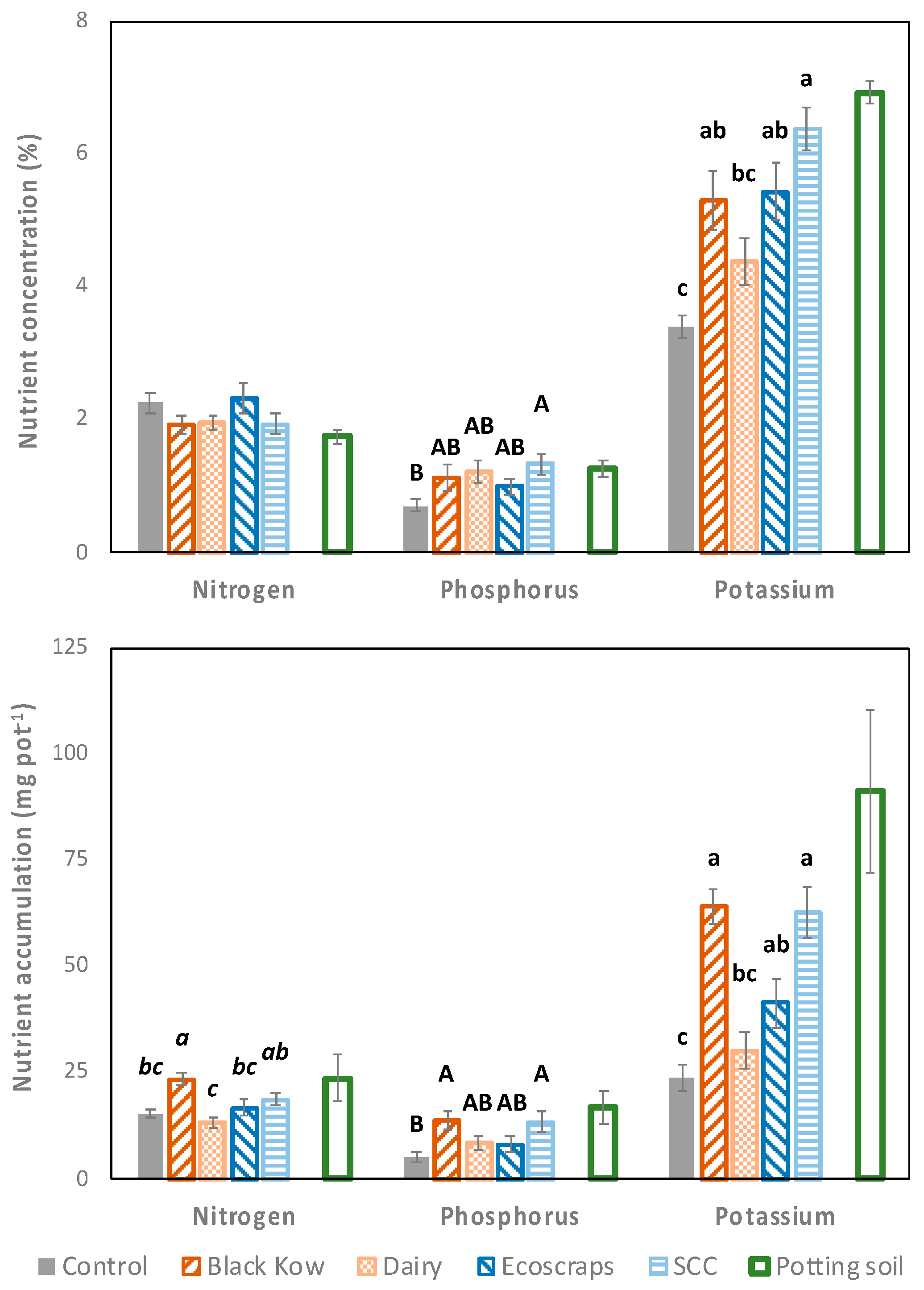
Plant nutrients were measured for all treatments in the Lake soil, but only for Black Kow, SCC, and Ecoscraps in the Millhopper soil; we only report data for the Lake soil in
Figure 4 given the lack of data for the control in the Millhopper soil. There were differences among treatments in the Lake soil for all variables measured (
Figure 4), except for N concentration (
p = 0.23). Tissue K concentration (
p < 0.001) and accumulation (
p < 0.001) were greater in the SCC (6.4% K, 63 mg K pot
−1), Ecoscraps (5.4% K, 41 mg K pot
−1), and Black Kow (5.3% K, 64 mg K pot
−1) treatments compared to the control (3.4% K, 24 mg K pot
−1). Tissue P concentration (
p = 0.04) and accumulation (
p = 0.01) were higher with SCC (1.3% P, 14 mg P pot
−1) relative to the control (0.7% P, 5 mg P pot
−1), and P accumulation was also greater with Black Kow (14 mg P pot
−1) relative to the control. Tissue N accumulation (
p < 0.001) was greater with Black Kow (23 mg N pot
−1) compared to the control (15 mg N pot
−1).
When including both soils for the Black Kow, SCC, and Ecoscraps treatments, tissue K concentrations were higher with SCC (7.0% K) than Black Kow and Ecoscraps (5.4–5.7% K; p = 0.001). In addition, spinach P concentration was higher in the Millhopper soil (1.5% P) than in the Lake soil (1.2% P; p = 0.003), but N concentration was higher in the Lake soil (2.1% N) relative to the Millhopper soil (1.3% N; p < 0.0001).
3.3. Regression Analyses
There were several positive albeit weak relationships (R2 ≤ 0.50) between biomass and soil or amendment properties. In the Lake soil, biomass increases were related to compost inputs of nitrate (R2 = 0.50, p < 0.001), PAN (R2 = 0.46, p < 0.001), S (R2 = 0.45, p < 0.001), Ca (R2 = 0.40, p < 0.001), Mg (R2 = 0.40, p = 0.001), resin P (R2 = 0.31, p = 0.005), and K (R2 = 0.18, p = 0.04) added at planting. Similar results were observed in the Millhopper soil where compost inputs of Ca (R2 = 0.46, p < 0.001), Mg (R2 = 0.42, p < 0.001), nitrate (R2 = 0.40, p = 0.001), PAN (R2 = 0.38, p = 0.002), resin P (R2 = 0.35, p = 0.003), K (R2 = 0.35, p = 0.003), TN (R2 = 0.34, p = 0.004), and S (R2 = 0.26, p = 0.01) added at planting were related to increases in biomass. All other regressions had p > 0.05.
Tissue N, P, and K concentrations and accumulation were positively related to several soil or amendment properties. In the Lake soil, N accumulation was directly associated with nitrate inputs (R2 = 0.53, p < 0.001); P concentration was weakly associated with compost inputs of resin-extractable P (R2 = 0.20, p = 0.03) and total P (R2 = 0.25, p = 0.01); P accumulation was associated with compost inputs of resin-extractable P (R2 = 0.41, p < 0.001) and total P (R2 = 0.40, p < 0.001); and tissue K concentration (R2 = 0.52, p < 0.0001) and accumulation (R2 = 0.55, p < 0.0001) were directly associated with compost K inputs. In the Millhopper soil, only K concentration was directly associated with K inputs (R2 = 0.75, p = 0.001). All other regressions had p > 0.05.
4. Discussion
4.1. Effects of Compost Feedstock and Soil Type on N Cycling
Black Kow (large-batch manure-based compost) and SCC (small-batch food-based compost) performed better than their respective counterparts. The four composts were added to provide 101 kg PAN ha
−1 based on half of the estimate of annual N availability provided by a commercial laboratory, with twice as much Black Kow added on a mass basis compared to the other treatments to achieve the desired application of N. Soil PAN differed among treatments, suggesting that this estimate may not have been adequate for this type of soil and/or study duration (two months). Soil PAN peaked around 4 weeks in both soils, similar to other studies [
25,
26], although peak soil PAN was almost 3-fold greater in the Lake soil (81 mg PAN kg
−1) relative to the Millhopper soil (31 mg PAN kg
−1). This could be due to the greater initial soil PAN and background N mineralization in the Lake soil, which could mask the effects of different composts. Amendments did not affect soil PAN significantly in the Lake soil whereas three composts (SCC, Black Kow, and Ecoscraps) had higher PAN relative to the unamended control and Dairy compost in the early stages of the experiment with the Millhopper soil. This indicates a greater effect of amendments in the less fertile Millhopper soil and/or a greater capacity to detect a signal from amendments in the soil with the lower background N concentration.
We observed an inverse relationship between compost C:N ratios and PAN at planting, where amendment PAN inputs and soil PAN increased in the order Dairy (C:N = 24) < Ecoscraps (C:N = 18) < Black Kow (C:N = 16) < SCC (C:N = 12). As composts with high C:N ratios can reduce N mineralization rates or even immobilize N [
27,
28], the immobilization of N observed with the Dairy compost in both soils is consistent with both its C:N ratio being above 20:1 and previous studies [
19,
29,
30]. Excluding the Dairy compost, all composts had similar soil PAN from week 6 through harvest, although high initial concentrations of soil PAN (specifically NO
3−) likely contributed to the higher biomass production observed with Black Kow and SCC. This is consistent with Parwada et al. [
31] who reported higher spinach yields with low C:N composts, and with Conesa et al. [
32] who reported greater spinach yield when nitrate concentrations were three-fold higher than ammonium concentration in the nutrient solution of hydroponic systems, compared to pure ammonium or pure nitrate.
The Ecoscraps compost was less homogenous than the other composts and had visible wood pieces, resulting in a higher C:N ratio than both Black Kow and SCC. The overall performance of Ecoscraps was intermediate compared to the other composts, suggesting that the mixture of plant materials, food waste, and processed forest products likely reduced N availability relative to a more homogeneous food compost like SCC. Given that Ecoscraps was added at half the rate of Black Kow based on estimated PAN release, it is possible that a higher Ecoscraps application rate, similar to what was used for Black Kow, could result in comparable results between the two amendments.
4.2. Effects of Compost Feedstock and Soil Type on P Cycling
Soil resin P decreased from planting to harvest for all treatments in both soils and in the potting mix, suggesting that leaching driven by uniform irrigation among treatments was the likely cause of these P losses. Phosphorus leaching is typically not the main loss pathway for P in most soils, which is often dominated by erosion and runoff, but P leaching is not uncommon in several Florida coarse-textured soils, including those used in this study [
14]. The larger decrease in soil P observed for the SCC and Black Kow amendments as compared to the control in the Lake soil is consistent with the greater yield and plant P accumulation in these treatments, where additional P accumulation was needed to support normal physiological growth in higher yielding treatments.
Unlike previous studies in field conditions [
33] or in urban gardens [
17], manure-based composts did not increase soil P at harvest in this study, which can become an issue if higher soil P results in P leaching. The only treatment that increased soil P at harvest was SCC in the Millhopper soil, consistent with the incubation study of Kelley et al. [
19] that found a larger P increase with a similar food-based compost relative to manure-based compost. The lack of change in soil P with manure-based compost could be driven by leaching losses in the coarse-textured soils used in this study, masking a potential effect of manure-based compost on soil P, as leachate was not collected. Small et al. [
17] found that raised-bed vegetable gardens that use manure-based compost as an amendment leached 4-fold or more P than food-based composts or unamended control, suggesting that this could be an important loss to be monitored in future studies, especially in coarse-textured soils that are potentially susceptible to P leaching [
14].
4.3. Effects of Compost Feedstock and Soil Type on K Cycling
In this study, the Lake soil had almost 5 times the amount of Mehlich-extractable K relative to the Millhopper soil. Potassium is easily leached from coarse-textured soils and deficiencies are observed in regularly cultivated and irrigated soils [
18], which could explain the lower concentrations found in the Millhopper soil. Based on current interpretations of Mehlich soil K concentrations, no additional K would be recommended for the Lake soil, although an input of 67 kg K ha
−1 would be recommended for the Millhopper soil [
24]. According to these recommendations, the only compost that supplied enough K was SCC at 72 kg ha
−1, with the next highest compost being Black Kow at 31 kg ha
−1. In the Millhopper soil, both the SCC compost and Black Kow produced a higher yield than the control. This is consistent with previous research reporting lower yields when spinach is grown in K-deficient soil [
34], although others have reported no impact of K deficiency on spinach [
35,
36].
In both soils, the amount of K applied in each amendment was directly related to tissue K concentration and biomass, although the relationship was stronger in the less fertile Millhopper soil. These results suggest that the application of the SCC compost, and to a lesser extent Black Kow, may have provided enough K to overcome any deficiencies inherent in the Millhopper soil. However, as the univariate regression analysis used here is a reductive approach, the cumulative effects of all compost properties are possibly more influential on plant growth and nutrient assimilation than any single property taken individually.
4.4. Implications
The best compost to use as a sole source of nutrients to improve plant growth depends on compost availability and soil type. Black Kow improved yield in both soils, provided PAN during the early growth stages and did not increase soil P at harvest relative to the control. The SCC compost similarly increased PAN during the early growth stages, improved yield, but increased soil P relative to the control at harvest, although these effects were observed only in the Millhopper soil. Overall, SCC performed similarly as Black Kow, but did so at half the mass application rate, indicating that SCC is a superior nutrient source, which confirms our main hypothesis.
Potassium inputs provided by the SCC compost in the Millhopper soil suggest that the yield increase may be due to satisfying plant K demand, at least partially. However, the yield benefits observed with Black Kow despite providing lower K inputs suggest that K availability was only one of the reasons why some composts increased yields whereas others did not.
Separated dairy solids did not improve yield and have been shown to immobilize N in this study and others [
19]. Thus, they will probably need to be coupled with N inputs from other sources to satisfy crop needs [
9], and target soil conditioning and/or increases in SOM rather than providing short-term fertility. Similarly, Ecoscraps did not improve yield, which was likely due to the high C:N resulting from the presence of processed forest products, suggesting that it should be used in a similar fashion as separated dairy solids, i.e., primarily as a soil conditioner.
Ultimately, characteristics of the compost that will improve plant growth need to be catered to the soil type. The Lake soil was recently converted to cropland and had high TC, TN, and K with an optimum pH of 6.6 (optimum range for spinach is between 6.0 and 6.8 [
24]). The Millhopper soil had a higher pH of 7.3, above the optimum for spinach, and lower concentrations of soil N, TC and K. All composts had a pH above the optimum range, hence selecting an amendment that will limit the pH increase while providing deficient nutrients would be recommended in these soils, especially for the Millhopper soil. In contrast, in more acidic soils with a pH below the optimum for spinach, the pH above 7 measured for all amendments could be advantageous and help increase the pH to a more favorable range for this crop.
5. Conclusions
Black Kow and SCC were the two composts that performed best in this experiment, increasing spinach biomass and nutrient availability relative to an unamended control. In contrast, Ecoscraps and Dairy did not contribute as much PAN and did not increase yields relative to an unamended control, suggesting that they will be most useful as soil conditioners rather than fertility sources. Overall, the combination of high PAN (especially nitrate) and K at planting seemed to have been the most important factors determining the appropriateness of different composts as a nutrient source in this experiment.
As Black Kow was added at twice the amount of SCC to achieve the desired N application rate, we conclude that SCC was a superior nutrient source, consistent with our previous incubation study [
19] and our hypothesis. However, the increase in soil P observed with SCC could be a concern in sandy soils prone to P leaching, although P supply will be advantageous in P-deficient soils. Furthermore, as food-based composts such as SCC divert organic waste from landfills, they help improve the circularity and sustainability of food systems.








{kind=link}
{kind=link}
{kind=link}
{kind=link}