Humic Acid Promotes the Growth of Switchgrass under Salt Stress by Improving Photosynthetic Function
by
Jiaxing Zhang
1,2, 
Qiuxia Meng
1,2,
Zhiping Yang
1,2,*,
Qiang Zhang
1,2,
Min Yan
1,2,*,
Xiaochan Hou
1,2 and
Xunzhong Zhang
3 1
Key Laboratory for Soil Environment and Nutrient Resources of Shanxi Province, Shanxi Agricultural University, Taiyuan 030031, China
2
Institute of Eco-Environment and Industrial Technology, Shanxi Agricultural University, Taiyuan 030801, China
3
School of Plant and Environmental Sciences, Virginia Polytechnic Institute and State University, Blacksburg, VA 24061, USA
*
Authors to whom correspondence should be addressed.
Agronomy 2024, 14(5), 1079; https://doi.org/10.3390/agronomy14051079
Submission received: 24 April 2024
/
Revised: 15 May 2024
/
Accepted: 16 May 2024
/
Published: 19 May 2024
(This article belongs to the Section Soil and Plant Nutrition)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:As a potential crop in saline-alkali land, the growth of switchgrass could also be threatened by salt stress. Promoting the growth of switchgrass under salt stress by humic acid has great significance in the utilization of saline-alkali land. In this study, a pot experiment was arranged to investigate the responses of photosynthetic and physicochemical characteristics of switchgrass to HA under salt stress. Results showed that humic acid increased the photosynthetic function of switchgrass and enhanced plant height by 41.1% and dry weight by 26.9% under salt stress. Correlation analysis showed that the membrane aquaporin gene PvPIP1, malondialdehyde, ascorbate peroxidase, abscisic acid, polyamine, and jasmonic acid were important factors affecting the photosynthetic function of switchgrass in this study. Meanwhile, HA reduced the content of malondialdehyde, indicating the alleviation of the membrane damage caused by salt stress. On the other hand, HA upregulated the relative expression of the PvPIP1 gene and activated ascorbate peroxidase, abscisic acid, polyamine, and jasmonic acid in switchgrass to resist salt stress. These improved the membrane stability and promoted the photosynthetic activity of switchgrass to enhance the plant’s tolerance against salt stress and growth. Results from this study are helpful to the efficient growing of switchgrass and the sustainable development of saline-alkali land.
1. Introduction
According to statistics, the area of saline-alkali land has reached approximately 11 million hectares worldwide [1]. Furthermore, a considerable proportion of the saline-alkali land has been left uncultivated due to the negative impact of salt on plant growth and crop production [2]. Switchgrass (Panicum virgatum L., Gramineae, Panicum), a gramineous plant grown as a forage crop or cellulosic biofuel feedstock, is salt tolerant, high-yield and can well adapt to barren soils [3,4]. This makes switchgrass a potential crop that can be used in the reclamation of saline-alkali land [5,6]. However, related reports have found that the growth of switchgrass is suppressed under high salt stress [7,8]. Therefore, enhancing the plant growth of switchgrass under salt stress is a challenge in promoting the utilization and benefit of saline-alkali land nowadays.
As abiotic stress, salinity in soil inevitably causes negative impacts on the photosynthetic efficiency of plants and contributes to the inhibition of plant growth [9]. In detail, the reduced soil water potential caused by high salinity can disrupt plant–water relationships and reduce cell turgor to lead to osmotic stress [10]. The blade structures are also affected by salinity stress, which reduces the stomatal conductance, transpiration, and gas exchange in leaves, thus weakening the photosynthesis of plants [11]. Moreover, plants also absorb salt via transporters and undergo increases of reactive oxygen species (ROS) to create oxidative stress. This contributes to lipid peroxidation, and membrane deterioration, as well as DNA and protein damage in leaves, which cause the disturbance of photosynthetic metabolism and inhibition of plant growth [12]. Therefore, saline-tolerant plants always deal with the detrimental effects of salinity during their growth through certain physiological responses such as upregulating related genes, transcription factors, proteins, and metabolites to maintain the essential processes of photosynthesis and growth [13,14]. For example, the osmoprotectants such as proline (Pro), glycine betaine (GB), sugars, and sugar alcohols that can sustain the cell turgor are synthesized in saline-alkali tolerant plants to maintain plant growth and yield [9]. Antioxidative substances including antioxidant enzymatic and non-enzymatic antioxidants are also increased to efficiently quench and scavenge excessive ROS to resist stress and maintain the cell membrane integrity and normal metabolism in the leaf [15,16]. Moreover, the plant also upregulates the level of endogenous phytohormones, such as abscisic acid (ABA), salicylic acid (SA), brassinosteroids (BRs), cytokinins (CTK), jasmonic acid (JA), gibberellins (GAs), polyamines, and auxins to mitigate salt stress [17]. These phytohormones play important roles in stabilizing the cellular pH environment, activating Na+/H+ antiporters, increasing cell water status, or enhancing antioxidant capacity under salt stress [18]. It was noteworthy that studies have shown that exogenous biostimulants could affect the release of antioxidant enzymes and phytohormones in plants to promote their adaptation to salt stress physiologically [19]. This suggested that adding biostimulants might be the key to enhancing the photosynthetic function of switchgrass by mobilizing physiological and biochemical responses, thus ensuring the efficient growth of switchgrass in salt conditions.
Humic acid (HA), a biostimulant contains one or more active substances that can stimulate nutrient uptake and the physiological metabolism of plants to resist abiotic stress, possibly promoting the growth of switchgrass in high-salt environments [20]. In nonsaline-alkali soil, Mora et al. observed that HA could effectively promote the root H+-ATPase activity and increase the content of ABA and other hormones in cucumber to improve shoot dry weight by 29.2–57.2% [21]. In the specific salt environment, Shukry et al. found an improved plant height (10.18%) of rice by 40 mg/L HA under salt stress, which was associated with the enhancement of the activities of antioxidant enzymes such as catalase and peroxidase [22]. Hatami et al. found HA enhanced the salt tolerance of almond rootstocks by increasing the protein and enzyme synthesis and decreasing the electrolyte leakage in leaves [23]. Saidimoradi et al. also found that HA mitigated the reduction in relative water content, membrane stability index, chlorophyll content, total biomass, and yield of strawberry to resist salt stress [24]. However, as a pasture plant with certain salt tolerance, previous studies have confirmed that HA promotes the germination, rooting and lignocellulose quality of switchgrass [25,26]. The response of physiological and biochemical indexes of switchgrass such as photosynthetic metabolism and plant growth to HA under salt stress remain unclear.
To explore the effects of HA on the growth of switchgrass under salt stress and clarify the related physicochemical mechanisms, HA was applied to switchgrass under salt stress in this study. The growth performance, basic photosynthetic indexes, membrane stability, antioxidant enzyme activity, phytohormones, and gene expression of switchgrass were measured, and then further evaluated the growth-enhancing effects of HA on switchgrass under salt stress by correlation analysis and redundancy analysis. The results of this study may provide a valuable strategy for the cultivation of switchgrass in saline-alkali soil and be of great significance to accelerate the utilization and reclamation of saline-alkali land.
2. Materials and Methods
2.1. Materials and Culture
BoMaster, a switchgrass variety with relatively higher salt tolerance at the five-leaf stage (Table S1), was selected for the test in this study. Full-grained, undamaged, and uniform-sized switchgrass seeds were taken for cultivation in climate boxes with a 14/10 h light/dark cycle, and 28/20 °C day/night temperature. Fifteen seeds were sown in each pot until the trefoil stage. When the switchgrass grew to the trefoil stage, it was transplanted into a plastic pot with a diameter of 32 cm and ensured 5 switchgrass in each pot. The soil in the pot was taken from the top soil of the experimental base in Shanxi Academy of Agricultural Sciences with the physiochemical parameters of pH value 6.92, electricity conductivity 140.27 μs cm−1, organic matter 15.13 g kg−1, total nitrogen 1.32 g kg−1, available potassium (K2O) 194.06 mg kg−1, and available phosphorus 42.2 mg kg−1. The soil was air-dried and screened (1 cm) to mix with the substrate at 2:1 (mass ratio) and filled 5 kg in each pot in total. While switchgrass grew to the 5-leaf stage (this stage is in the rapid growth period of switchgrass and is greatly affected by the external environment), salt stress and humic acid tests are started.
The humic acid (HA) extracted and purified from the weathered coal collected in Jingle County, Shanxi Province, China with the potassium hydroxide method was used in this study. The HA has been proven to have an excellent promoting effect on the regeneration of mung bean roots in our previous study [27]. This HA performed specific properties as follows: elemental composition including 70.20% carbon, 25.79% oxygen, 2.45% hydrogen, 1.40% nitrogen and 0.16% sulfur; contained 79.29 ± 0.54% total humic acid, 68.20 ± 0.38% free humic acid, 1.74 ± 0.11% calcium and magnesium, 3.86 ± 0.06 mmol g−1 total acid group, 0.34 ± 0.04 mmol g−1 carboxyl, 3.52 ± 0.03 mmol g−1 phenolic hydroxyl; F/T value was 86.01%, E4/E6 value was 4.39, E270/E400 value was 3.08, E280/E472 value was 5.04 and E472/E664 value was 3.88.
2.2. Treatments and Sampling
The test was conducted in the greenhouse of Shanxi Agricultural University. Switchgrass seedlings in the five-leaf stage were subjected to three treatments: (1) Non-stressed control (CK): tap water; (2) Salt Stress (SS): 250 mmol/L NaCl; (3) Salt Stress (250 mmol/L NaCl) plus 0.05% HA (SS+HA). Switchgrass was transplanted into a 32 cm diameter pot individually and 9 pots of switchgrass were contained in each treatment. Thereinto, the salt concentration of 250 mmol/L was confirmed by reference to the salt concentration selected in the salt tolerance study of perennial ryegrass (Lolium perenne L.) [11], and the concentration of humic acid at 0.05% was selected because it promoted the rooting of switchgrass the most in our previous study [27]. Importantly, to avoid the salt-shock effect, aliquots of 200 mL NaCl solution at 50 mmol/L were used for the first irrigation, 150 mmol/L for the second irrigation, and 250 mmol/L for the third irrigation. These three irrigation sessions were completed within 24 h and maintained concentrations by measuring the conductivity of the soil thereafter. The control treatment irrigated the same amount of tap water at the same time. Sampling and measurement were performed on days 0, 7, 14, 21, 28, and 35 of the experiment. The third leaf of the apical leaf sequence from top to bottom was collected, a small amount of fresh leaf tissue was used for electrolyte leakage, malondialdehyde, and proline analysis, and the remaining parts were stored at −80 °C for the subsequent determination of physiological and chemical indexes. Three samples were taken from each treatment to eliminate individual differences, and each sample was repeated three times to eliminate measurement errors.
2.3. Growth Indexes
Thirty-five days after treatment, the plant height of switchgrass was measured from the soil level to the tip of the tallest leaf. The leaf length and leaf width were measured by selecting the third leaf from the top of each switchgrass. Thereafter, the entire plant was taken out, washed and dried in a drying oven at 105 °C for 30 min and then dried at 65 °C to constant weight. Finally, the dry weight of aboveground biomass of the single switchgrass was measured.
2.4. Photosynthetic Indexes
The photosynthetic indexes photosynthetic rate, stomatal conductance, intercellular carbon dioxide concentration, and transpiration rate were measured by a portable photosynthesis system (LI–6400XT, Licor Corporation, Lincoln, NE, USA) and determined during 8:00–11:00 a.m. This instrument was set at a temperature of 20 °C, relative humidity (RH) 50 ± 10%, photosynthetic photon flux density (PPFD) 400 μ mol·m−2 s−1, CO2 420 μ mol·mol−1 and flow rate 500 μ mol·s−1 in leaf chamber [28].
2.5. Malondialdehyde
To determine the concentration of malondialdehyde (MDA), fresh leaf samples were ground with a pestle in a mortar that contained 5 mL 0.6% thiobarbituric acid (TBA) in 10% trichloroacetic acid (TCA) [29]. Thereafter, samples were cooled on ice for 5 min after heating at 100 °C for 15 min, then centrifuged for 10 min at 5000 rpm, and the absorbance of the supernatant was measured at 450, 532, and 600 nm (OD450, OD532, and OD600), respectively. Eventually, the MDA content was calculated as follows:
MDA (nmol MDA g−1 FW) = 6.45 (OD532 − OD600) − 0.56 (OD450)
2.6. Relative Electrolyte Leakage
The plant’s relative electrolyte leakage (REL) was measured based on the conductivity measurement [30]. A total of 1 g fresh leaf samples were incubated for 4 h in 80 mL of dd H2O after cutting into disks of 0.8 cm to measure the electrical conductivity (C1) with a conductivity meter. Then the solution was boiled for 10 min and cooled down to room temperature to measure the electrical conductivity (C2). The REL was calculated as follows:
2.7. Proline
According to the method mentioned described by Wu et al., the proline content was determined by a spectrophotometer (Jenway 6705, Bibby Scientific, Staffordshire, UK) [11]. Fresh leaf samples (50 mg) were pulpified with 1.8 mL 3% (w/v) sulfosalicylic acid and boiled at 100 °C for 10 min. Then, 1 mL supernatant was mixed with 1 mL acetic acid and 1 mL acidic ninhydrin to heat at 100 °C for 40 min. The reaction mixture was extracted with 2 mL toluene after cooling and then record the absorbance at 520 nm in a spectrophotometer.
2.8. Superoxide Dismutase
The frozen leaf sample (500 mg) was homogenized with the homogenization solution of 10 mL HEPES buffer (50 mM) and 0.1 mM Na2-EDTA (pH 7.6). The homogenate was centrifugated (15,000 rpm for 15 min at 4 °C) and the extract was used to assay the superoxide dismutase (SOD). After sitting at 4 °C overnight, the extract was dialyzed against a dilute homogenizing solution to separate the low-molecular-mass substances that interfere with the SOD assay [31]. The activity of SOD was assayed with the procedure based on the observation of photochemical reduction inhibition of NBT (nitro blue tetrazolium) [32]. Then, 5 mL reaction mixture [50 mM HEPES (pH 7.6) + 0.l mM EDTA + 50 mM Na2CO3 (pH 10.4) + 13 mM methionine + 0.025% (w/v) Triton X-l00 + 75 mM NBT + 2 mM riboflavin + 0.2 mL enzyme extract] was prepared and then illuminated for 15 min by intensity of light (350 µMm−2s−1). One unit of activity of SOD was specified as the enzyme quantity causing a 50% inhibition of NBT reduction as observed spectrophotometrically (560 nm).
2.9. Ascorbate Peroxidase and Peroxidase
To obtain the enzyme extract for ascorbate peroxidase (APX) and peroxidase (POD) measurement in samples, 0.1 g frozen leaf was ground in liquid nitrogen with an ice-cooled mortar and pestle. The mixture was transferred into the buffer and then homogenized at 12,000 rpm for 20 min at 4 °C. Indeed, the buffer consisted of 25 mmol L−1 K–P (pH = 7), 0.4 mmol L−1 EDTA-4H, Tris-HCl, and 2% (w/v) polyvinylpolypyrrolidone [33].
Thereafter, 0.75 mL K–P (0.1 mol L−1) buffer (pH = 7), 0.75 mL ascorbic acid (1 mmol L−1), 0.75 mL EDTA-4H (0.4 mmol L−1), 0.03 mL H2O2 (10 mmol L−1), 0.57 mL of distilled water, and 10 mmol L−1 enzyme extract was mixed in a 3 mL reaction volume to assess the activity of ascorbate peroxidase. H2O2 was added to ascorbate to start the oxidation process. The decrease in absorbance at 290 nm was measured to reflect ascorbate oxidation [29].
To determine the activity of POD, 20 mM phosphate buffer (pH = 7), 5 mM 2-methoxy phenol (guaiacol), and 1 mM hydrogen peroxide were used to prepare the reaction mixture [34]. The absorbance at 470 nm after tetraguaiacol formatting was determined. For each unit of peroxidase activity, 1 mol of guaiacol oxidized for 1 min is catalyzed by the enzyme.
2.10. Phytohormones
The production of phytohormones including abscisic acid, polyamine, cytokinin, indoleacetic acid brassinolide, and jasmonic acid in leaves were all determined using the ELISA (enzyme-linked immunosorbent assay) kit (MLBio ELISA Kit producers, Shanghai, China) according to manufacturer’s instruction. Briefly, the leaves were ground on ice supplemented with quartz and sodium phosphate buffer. The mixture was collected and incubated at 4 °C for 6 h. The supernatant by centrifugation was collected to measure the contents of phytohormones in switchgrass [35].
2.11. Gene Expression
Total RNA from switchgrass was extracted by RNA kit (Tiangen Biotech, Beijing, China) after the leaf tissue was powdered in a pre-cooled mortar. RNA samples were treated with a gDNA Eraser to eliminate any DNA contamination and then used for cDNA synthesis by Evo M-MLV Reverse Transcription Kit (Accurate Biology, Changsha, China) according to the manufacturer’s instructions. Thereafter, real-time polymerase chain reaction (RT-PCR) was performed using the One-Step RT-PCR Kit (Vazyme, Nanjing, China). The universal primer sets were 5′-TCTGTTTTTCTCTATGTGCAGGA-3′ and 5′-CTGAGGAGCTGTTCTTGGCA-3′ for actin gene, 5′-CCAATTGGGTTCGCCGTGTT-3′ and 5′-TAGGACCTGCTCTTGAACGGGA-3′ for PvPIP1 gene (a gene related to aquaporins and upregulated in drought stress) [36]. Relative expression levels were calculated using the formula 2−ΔΔCT [37]. All the samples realized three biological replicates and three technical replicates in this measurement.
2.12. Data Analysis
SPSS 25.0 was employed for the analysis of differences among the detection data of all treatments with p < 0.05. Origin 2022 was employed for correlation analysis to assess the effects of the membrane aquaporin PvPIP1 gene on the photosynthetic function and membrane stability of switchgrass. Canoco 5 was employed for redundancy analysis to comprehensively evaluate the photosynthetic function of switchgrass in response to the membrane stability, antioxidant enzyme, and phytohormones under the influence of salt and humic acid.
3. Results
3.1. Growth Performance and Photosynthetic Function
As shown in Figure 1A, the influence of salt stress and HA on switchgrass was directly reflected in growth performance. Over 35 days of cultivation, salt stress inhibited the growth of switchgrass with plant height reduced to 70.8 ± 3.9 cm. This was 15.0% lower than that of the control (CK) (83.3 ± 3.6 cm) (p < 0.05). Moreover, the dry weight of the aboveground biomass was also reduced by 20.5% to 5.39 ± 0.38 g, as well as the decrease in leaf length and width (Table S2) (p < 0.05). However, HA increased the height of switchgrass to 99.9 ± 5.7 cm and increased the weight to 6.84 ± 0.59 g under salt stress, contributing to 41.1% and 26.9% enhancements in plant growth, respectively. It was noteworthy that the plant height of switchgrass with HA addition under salt stress (SS+HA) was even higher than that in the control without salt stress (CK). As for the dry weight of shoots, leaf length and leaf width, there were no significant differences between the control and treatment of SS+HA (p > 0.05), showing the reversing effect of HA on the growth parameters of switchgrass under salt stress.
Photosynthetic indexes analysis also showed the photosynthetic function of switchgrass was suppressed by salt stress but recovered and improved by HA. Compared with the control, salt stress decreased the net photosynthetic rate (Pn) (Figure 1B), transpiration rate (Tr) (Figure 1C), and stomatal conductance (Gs) (Figure 1D) but increased the intercellular carbon dioxide content (Ci) (Figure 1E) of switchgrass during the test period. The differences between the two groups became significant from day 14 and the changes eventually reached 20.1%, 18.86%, 10.23%, and 7.92% on day 35, respectively. However, with the addition of humic acid under salt stress (SS+HA), Pn, Tr, and Gs became 7.98–44.35%, 5.06–22.13%, and 8.77–20.25% higher than those of the salt stress treatment (SS), respectively. Thereinto, Pn was significantly increased from the 7th day after adding HA in the presence of salt stress. The differences in Tr and Gs between the treatments of SS and SS+HA also became significant from day 14. The Ci under salt stress was reduced by 9.39–22.13% during the cultivation with HA added, which was even 3.13–15.29% lower than that of the control and showed a significant difference from day 7. On the other hand, Figure 2 also showed under the influence of salt stress and HA, the photosynthesis indexes of switchgrass had a strong correlation with the growth performance. In detail, the growth indexes were positively correlated with Pn, Gs and Tr but negatively correlated with Ci. Moreover, the positive correlation of dry weight of switchgrass shoots with Tr and Gs, and the positive correlation of leaf width with Tr were statistically significant.
3.2. Membrane Stability and Oxidase Activity
The relative electrolyte leakage rate (REL) of blade, proline, malondialdehyde (MDA), superoxide dismutase (SOD), peroxidase content (POD), ascorbic acid peroxidase (APX) in switchgrass were all in dynamic changes under the influence of salt stress and HA (Figure 3). Compared with the control (CK), salt stress significantly increased the REL (Figure 3A), proline (Figure 3B), MDA (Figure 3C), SOD (Figure 3D), and POD (Figure 3E) of switchgrass from day 7 and underwent uptrends, which increased by 88.77%, 79.79%, 221.52%, 54.60% and 113.00% on the 32nd, 21st, 28th, 7th, and 14th day, respectively. By contrast, the APX activity was decreased by salt stress and it was 71.43% lower than that of the control on the 28th day. However, with the addition of HA, the effect of salt stress on the physiological indexes was significantly weakened. Over 35 days of cultivation, the REL, Pro, MDA, SOD, and POD in switchgrass were maximally reduced by 26.18%, 29.44%, 47.61%,46.84%, and 34.05% respectively. Among them, the content of Pro in the treatment of SS+HA tended to be similar to that of CK while the SOD and POD activity was even lower than that of CK. On the other hand, HA also effectively mitigated the decrease in APX caused by salt stress in switchgrass (Figure 3F). The APX activity in the treatment of SS+HA was increased to 164.29–228.57 μmol mm−1 g−1 during the test period, which was 77.78–333.29% higher than that in the salt stress treatment. Interestingly, the APX activity in the treatment of SS+HA was significantly higher (6.67% and 23.81%) than that in non-stressed control (CK) on day 21 and day 28.
3.3. Phytohormones
The effects of HA on phytohormone concentrations in switchgrass under salt stress are shown in Figure 4. Affected by salt stress, the contents of abscisic acid (ABA) (Figure 4A) and polyamine (PA) (Figure 4B) were decreased, while cytokinin (CTK) (Figure 4C), indoleacetic acid (IAA) (Figure 4D), brassinolide (BR) (Figure 4E) and jasmonic acid (JA) (Figure 4F) were increased since day 7. The changes of these phytohormones reached the maximum on the 35th, 14th, 14th, 7th, 14th and 35th days, respectively, which were −77.43%, −14.58%, 122.81%, 64.91%, 15.92% and 3.82% higher in comparison with that in control. With the addition of HA under salt stress (SS+HA), the content of phytohormones in switchgrass changed dramatically. Thereinto, HA increased ABA, PA, and JA in switchgrass by 17.03–344.45%, 7.74–15.85%, and 1.15–10.89% under salt stress, respectively. Moreover, the PA and JA in SS+HA were also higher than that of the control (CK) since the 21st and 7th days. Moreover, the contents of CTK, IAA, and BR in the treatment of SS+HA were reduced by 14.55–28.93%, 6.02–11.97%, and 9.17–15.94% as compared to the treatment of SS. It was noteworthy that the concentrations of CTK and IAA in SS+HA were still higher than that in control and the differences between these two treatments were still significant (p < 0.05). On the contrary, the BR content in switchgrass was 5.08–12.27% lower than that of the control except for the 14th and 35th days.
3.4. Gene Expression
As was illustrated in Figure 5A, the expression of membrane aquaporins PvPIP1 gene in switchgrass leaves was significantly changed (p < 0.05) under the influence of salt stress and HA. In comparison with the control, the relative expression of PvPIP1 in switchgrass sharply increased and then dropped during the 35 days of cultivation with the influence of salt stress. The relative expression of PvPIP1 underwent a significant increase on day 7 and day 14, which was 347.78% and 189.50% higher than that in the control. Thereafter, PvPIP1 expression decreased to a lower level than that in control from day 21, and the difference became significant on the 35th day (p < 0.05). Compared with the salt stress group (SS), adding HA decreased the expression of PvPIP1 within the first two weeks and then enhanced it from day 21 to day 35. In detail, the relative expression of the PvPIP1 gene reached 3.79 and 1.49, respectively, after 7 and 14 days of cultivation, representing a 15.73% and 48.57% decrease compared to that in salt stress. Thereafter, the expression of PvPIP1 in the treatment of SS+HA remained between 0.86 and 1.44 during day 21 and day 35, showing 41.26–135.21% enhancement in comparison with that in the treatment of SS. Meanwhile, the differences in PvPIP1 gene expression between the treatments SS+HA and CK on the 14th, 28th, and 35th days were not significant (p > 0.05), indicating a converting effect of HA on the upregulation of PvPIP1 expression caused by salt stress.
To further explore the relationships of PvPIP1 and leaf functions such as membrane stability and gas exchange, correlation analysis was performed and the results are shown in Figure 5B. Results showed that the expression of PvPIP1 in switchgrass was positively correlated with the net photosynthetic rate (Pn), transpiration rate (Tr), and proline content (Pro) in leaves but negatively correlated with the intercellular carbon dioxide content (Ci) and malondialdehyde (MDA) in leaves to some extent. The correlation coefficients of the PvPIP1 expression with its negative correlation index Ci and MDA were also only −0.47 and −0.39. However, it was noteworthy that there was a significant correlation (p < 0.05) between the PvPIP1 gene expression and Ci in switchgrass leaves. This provides an opportunity to further elucidate the mechanism of the promoting effect of HA on the growth of switchgrass under salt stress.
3.5. Redundancy Analysis
As the most direct index that influences plant growth, the photosynthetic function of switchgrass was analyzed by testing the physiological indexes such as peroxidase activity and phytohormones concentrations, respectively, by redundancy analysis in this study (Figure 5). Results showed that the photosynthetic index of switchgrass leaves was driven by enzymes and phytohormones in switchgrass under the effects of salt stress and HA. Indeed, the Ci was positively correlated with the contents of proline, MDA, REL, and SOD in switchgrass, and their correlations were MDA, SOD, REL, and proline in descending order (Figure 6A). Furthermore, similar positive correlations among photosynthetic indexes were also shown in the correlation analysis of phytohormones (Figure 5B). On the other hand, there were remarkably significant positive correlations among the Pn, Tr, and Gs in switchgrass. These three photosynthetic indexes were positively correlated with the APX and negatively correlated with the POD in switchgrass. Figure 5B observed the same results, which also showed that the Pn, Tr, and Gs in switchgrass were significantly correlated with the APX and POD in leaves.
As shown in Figure 6, the relative degree of explanation for phytohormones was 41.5%, which was lower than that of enzymes (80.2%). This indicated that phyto-hormones had less effect on the photosynthetic indexes of switchgrass in comparison with enzymes. Thereinto, Ci was positively correlated with BR, IAA, and CTK (BR > CTK > IAA) in switchgrass, and negatively correlated with JA, BA, and PA (JA > ABA > PA). While Pn was positively correlated with PA and BA (PA > BA), and negatively correlated with CTK, IAA, and BR (CTK > BR > IAA). However, the Tr and Gs in switchgrass did not appear a strong correlation with the phytohormones, which were only slightly negatively correlated with JA, CTK, and IAA in this study.
4. Discussion
In this study, the photosynthetic function and plant growth of switchgrass were seriously suppressed by salt stress but restored and even enhanced by HA (Figure 1). Photosynthesis is the most important physical and chemical process that converts carbon dioxide into organic matter and thus has a positive correlation with growth performance in a significant manner. Arif et al. have described that salinity can affect the ultrastructure, chlorophyll, and carotenoid synthesis in plant cells to interfere with photosynthesis. It was thought to be the salinity in this study that damaged the photosynthetic functions of witchgrass leaves to inhibit photosynthetic function [9], which was manifested directly as the increase in Ci and the decrease in Pn, Tr, and Gs directly. Then, it decreased the leaf length, leaf width and plant height of switchgrass and reduced biological yield eventually. On the contrary, the addition of HA effectively improved the photosynthetic function and growth parameters of switchgrass under salt stress. Popescu and Popescu have confirmed that HA could maintain the cell membrane stability and ultrastructure of mesophyll cells to alleviate the non-stomatal restriction of photosynthesis, which eventually enhanced photosynthesis in leaves under stress [38]. This study also showed that HA significantly enhanced the Pn, Tr, and Gs in switchgrass and reduced Ci under salt stress. The growth parameters of switchgrass in the treatment of SS+HA were also very close to those of the non-stressed control (CK), confirming the potential of HA in promoting the metabolism of switchgrass and enhancing plant resistance to salt stress [39]. Certainly, these enhancements were caused by the combined effects of physiology and biochemistry in switchgrass that need to be further verified [40].
Physiological indexes are of great significance to explore the effects of stress environment on plant ontology. In this study, REL, Pro, and MDA of switchgrass were all significantly enhanced by salt stress and then reduced with the addition of HA (Figure 3). It was well known to us that the REL and Pro in plants are generally used to reflect membrane stability given their correlation with membrane permeability [41,42]. MDA, the important product of membrane lipid peroxidation, is positively correlated with the degree of stress and membrane damage. Therefore, with the aggravation of injury, the accumulation of MDA in plant leaves increases under salt stress and indicates serious destruction of membrane structure and membrane permeability, which leads to the increased leakage rate of leaf electrolyte, while Pro rises at the same time to maintain cytoplasmic osmotic to defend against stress [43]. Numerous studies have confirmed that exogenous substances are effective in preventing cell membranes from being damaged and stabilizing proteins and enzyme activities to regulate the cell osmotic potential and alleviate salt stress in plants. Radhakrishnan and Lee reported that spermidine (Spd) mitigated the rising of total phenol and MDA contents to ameliorate salt stress effects in cucumber plants [44]. Hatami et al. found that HA increased the soluble proteins and enzyme synthesis to reduce electrolyte leakage and enhance almond rootstock growth under salt stress [24]. In the present study, HA resulted in the synchronized decrease in REL, proline, and MDA in switchgrass leaves under salt stress. These confirmed that HA can alleviate the damage of switchgrass from salt stress and promote the cell osmotic regulation and the stability of the membrane, thus improving the tolerance of switchgrass under salt stress and promoting switchgrass photosynthesis and growth.
The antioxidative system is indispensable in determining plant stress tolerance. With the influence of salt stress, the activity of SOD and POD in switchgrass were significantly increased but APX decreased (Figure 3) in this study. These antioxidant enzymes in plants can scavenge the reactive oxygen species (ROS) that could oxidize the lipid bilayer and destabilize the structure and function of the proteins, lipids, and nucleic acids, to reverse physiological setbacks under salt stress [45]. This means that APX activity in switchgrass was inhibited by salt stress, while SOD and POD activity was upregulated to resist the ROS from salt stress. However, adding HA under salt stress reduced the activity of SOD and POD but increased APX significantly in this study. It has been reported that exogenous substances could alleviate salt stress in plants by inducting antioxidant enzyme activity. Oliveira et al. found that brassinosteroids (EBR) increased APX activity in Eucalyptus urophylla plants by 51% and promoted plant salt tolerance [46]. Feng et al. also reported that the exogenous substance Spd significantly increased the activities of SOD, CAT, and APX and improved the salt tolerance of rice [19]. Yan et al. also observed that the overexpression of APX effectively promoted sweet potato growth and played an important role in improving its salt tolerance [47]. The phenomena observed in this study indicated that the application of HA possibly reduced the ROS in switchgrass to relieve the need for antioxidant enzymes SOD and POD on the one hand and promoted the production of APX in switchgrass to improve its salt tolerance on the other hand.
As essential substances produced inside plants, phytohormones play a critical role in regulating plants’ physiological processes and metabolisms. For instance, Liu et al. reported that H2O2 promoted the germination of Arabidopsis sp. seeds under salt stress by promoting the metabolism of ABA [48]. JA can not only mediate plant responses to salt stresses by regulating the transcription of stress-responsive genes including antioxidant genes under stress conditions [49], but also can interact with ABA to modulate the abiotic stress response and tolerance [50]. In this study, compared with the non-stressed control, ABA and PA in switchgrass were inhibited by salt stress, and CTK, IAA, and BR were increased to resist salt stress (Figure 4). With the addition of HA under salt stress, the increased ABA, PA, and JA confirmed that HA induced the production of these phytohormones in switchgrass to better cope with the salt stress. On the other hand, the decrease in CTC, IAA, and BR was also the signal that HA mitigated the salt damage to switchgrass, thereby reducing the response of these phytohormones under salt stress. These changes in phytohormones were beneficial to the growth and metabolism of switchgrass under salt stress.
It is known that membrane aquaporin is beneficial to the transmembrane transport of water and other small molecules, which can regulate physiological processes such as stomatal movement and cell growth that affect photosynthesis in leaves [51]. In this study, the relative expression of membrane aquaporin gene PvPIP1 in switchgrass increased significantly and then dropped sharply to a low level under salt stress (Figure 5A), which indicates that switchgrass tried to cope with salt stress by upregulating PvPIP1 expression but was heavily affected by salinity and underwent severe gene dysfunction in the latter test period. With the addition of HA under salt stress, the expression of PvPIP1 decreased within the first two weeks but remained higher level than that in the treatments of CK and SS over the last three weeks, suggesting that the HA may have upregulated the PvPIP1 expression to alleviate salt stress on switchgrass and enhanced potential to resist salt stress by humic acid [52]. In particular, the relative expression of membrane aquaporin gene PvPIP1 was positively correlated with Pn and Tr but significantly negatively correlated with Ci in this study (Figure 5B). This suggested that the expression of the PvPIP1 gene could promote the movement of stomatal, which accelerated leaf transpiration and gas exchange to reduce intercellular carbon dioxide accumulation and promoted photosynthesis in switchgrass. Moreover, the correlation analysis in this study also revealed that PvPIP1 expression in switchgrass was positively correlated with Pro and negatively correlated with MDA. This suggested that the expression of the PvPIP1 was beneficial in inducing the increase in osmoregulatory substance Pro to maintain the osmotic balance and eventually the membrane stability and integrity of cells, thus providing a better condition for photosynthesis of switchgrass under salt stress.
In addition to the expression of the PvPIP1 gene, redundancy analysis showed that the increased MDA in leaves by salt stress was the main factor that damaged the membrane function of switchgrass and contributed to the increase in REL (Figure 6A). This significantly declined the exchange of carbon dioxide in switchgrass leaves and limited photosynthetic function under salt stress. With the addition of HA under salt stress, APX in switchgrass increased and showed strong positive correlations with the main photosynthetic indexes Pn, Tr, and Gs. On the other hand, phytohormones PA, JA, and ABA in switchgrass were increased by HA and positively and strongly correlated with Pn (Figure 6B) under salt stress. It was confirmed by other researchers that the presence of ABA and JA could promote the expression of biosynthetic genes and cellular content [53]. This suggests that the HA induced the release of ABA, JA, and PA in switchgrass under salt stress to jointly improve the photosynthesis of switchgrass, thus promoting switchgrass growth under salt stress. This is evidence that HA promoting the photosynthetic function of switchgrass under salt stress is the result of the collaboration of multiple physiological and biochemical functions.
5. Conclusions
Salt stress increases the production of harmful substances such as MDA and ROS in switchgrass, reduces membrane stability and inhibits the photosynthetic function of leaves to restrict switchgrass growth. While HA can alleviate the damage of salt stress on switchgrass. The decrease in MDA and proline, the activation of the oxidative stress system (including SOD and POD), the increase in growth promotion phytohormones (including CTK, IAA, and BR), and the upregulation of PvPIP1 expression in switchgrass proved that the damage of salinity stress on switchgrass was alleviated by HA. Moreover, HA also effectively improved the stability of leaf membrane cells (significant decrease in REL), induced the increased production of APX and defense phytohormones PA, ABA, and JA, and stabilized the expression of PvPIP1 gene to improve the air exchange and photosynthesis capacity of leaves under stress. Taken together, these factors improved the salt tolerance of switchgrass and eventually promoted the growth of switchgrass under salt stress.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14051079/s1, Table S1: Effects of salt stress on plant height and survival rate of different varieties of switchgrass at the five-leaf stage; Table S2: Effect of salt stress and humic acid on switchgrass leaf length and leaf width.
Author Contributions
Conceptualization, Z.Y. and Q.Z.; methodology, M.Y. and X.Z.; formal analysis, J.Z. and X.H.; investigation, J.Z. and X.H.; resources, M.Y.; data curation, J.Z. and X.H.; writing—original draft preparation, J.Z.; writing—review and editing, Q.M., X.Z. and J.Z.; supervision, Q.Z. and M.Y.; project administration, Z.Y. and M.Y.; funding acquisition, Z.Y. and Q.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Key Research and Development Project of Shanxi Province (202202140601010), the Merit-based Open Competition Project of Shanxi Major Science and Technology Special Program (202201140601028), and the Special Project for Research Platforms and Talents of Shanxi Province (201805D211017). These three projects were all approved by Science and Technology Department of Shanxi Province, China.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhao, W.; Zhou, Q.; Tian, Z.; Cui, Y.; Liang, Y.; Wang, H. Apply biochar to ameliorate soda saline-alkali land, improve soil function and increase corn nutrient availability in the Songnen plain. Sci. Total Environ. 2020, 722, 137428. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef]
- Kim, J.; Liu, Y.; Zhang, X.; Zhao, B.; Childs, K.L. Analysis of salt-induced physiological and proline changes in 46 switchgrass (Panicum virgatum) lines indicates multiple response modes. Plant Physiol. Biochem. 2016, 105, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Niu, Z.; Yue, Y.; Su, D.; Ma, S.; Hu, L.; Hou, X.; Zhang, T.; Dong, D.; Zhang, D.; Lu, C.; et al. The characterization of streptomyces alfalfae strain 11f and its effect on seed germination and growth promotion in switchgrass. Biomass Bioenergy 2022, 158, 106360. [Google Scholar] [CrossRef]
- Yu, S.; Wu, J.; Wang, M.; Shi, W.; Xia, G.; Jia, J.; Kang, Z.; Han, D. Haplotype variations in QTL for salt tolerance in Chinese wheat accessions identified by marker-based and pedigree-based kinship analyses. Crop J. 2020, 8, 1011–1024. [Google Scholar] [CrossRef]
- Hu, Q.; Zhao, Y.; Hu, X.; Qi, J.; Suo, L.; Pan, Y.; Song, B.; Chen, X. Effect of saline land reclamation by constructing the “raised field -shallow trench” pattern on agroecosystems in yellow river delta. Agric. Water Manag. 2022, 261, 107345. [Google Scholar] [CrossRef]
- Anderson, E.K.; Voigt, T.B.; Kim, S.; Lee, D.K. Determining effects of sodicity and salinity on switchgrass and prairie cordgrass germination and plant growth. Ind. Crops Prod. 2015, 64, 79–87. [Google Scholar] [CrossRef]
- Zhuo, Y.; Zhang, Y.; Xie, G.; Xiong, S. Effects of salt stress on biomass and ash composition of switchgrass (Panicum virgatum). Acta Agric. Scand. Sect. B Soil Plant Sci. 2015, 65, 300–309. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Gao, G.; Yan, L.; Tong, K.; Yu, H.; Lu, M.; Wang, L.; Niu, Y. The potential and prospects of modified biochar for comprehensive management of salt-affected soils and plants: A critical review. Sci. Total Environ. 2024, 912, 169618. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Q.; Ervin, E.H.; Yang, Z.; Zhang, X. Physiological mechanism of enhancing salt stress tolerance of perennial ryegrass by 24-epibrassinolide. Front. Plant Sci. 2017, 8, 1017. [Google Scholar] [CrossRef] [PubMed]
- El Ghazali, G. Suaeda vermiculata forssk. Ex j.f. Gmel.: Structural characteristics and adaptations to salinity and drought: A review. Int. J. Sci. 2020, 9, 28–33. [Google Scholar] [CrossRef]
- de Freitas, P.A.F.; de Carvalho, H.H.; Costa, J.H.; Miranda, R.D.; Saraiva, K.D.D.; de Oliveura, F.D.B.; Coelho, D.G.; Prisco, J.T.; Gomes, E. Salt acclimation in sorghum plants by exogenous proline: Physiological and biochemical changes and regulation of proline metabolism. Plant Cell Rep. 2019, 38, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, J.J.; Miras-Moreno, B.; Araniti, F.; Salehi, H.; Bernardo, L.; Parida, A.; Lucini, L. Proteomics revealed distinct responses to salinity between the halophytes Suede maritima (l.) Dumort and Salicornia brachiata (roxb). Plants 2020, 9, 227. [Google Scholar] [CrossRef]
- Mbarki, S.; Sytar, O.; Cerda, A.; Zivcak, M.; Rastogi, A.; He, X.; Zoghlami, A.; Abdelly, C.; Brestic, M. Strategies to mitigate the salt stress effects on photosynthetic apparatus and productivity of crop plants. Salin. Responses Toler. Plants 2018, 1, 85–136. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; Ganie, S.A.; Hossain, M.S.; Gomaa, A.M. Seed priming with cypress leaf extract enhances photosynthesis and antioxidative defense in zucchini seedlings under salt stress. Sci. Hortic. 2022, 293, 110707. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Hussain, S.; Hafeez, M.B.; Azam, R.; Mehmood, K.; Aziz, M.; Ercisli, S.; Javed, T.; Raza, A.; Zahra, N.; Hussain, S.; et al. Deciphering the role of phytohormones and osmolytes in plant tolerance against salt stress: Implications, possible cross-talk, and prospects. J. Plant Growth Regul. 2023, 43, 38–59. [Google Scholar] [CrossRef]
- Feng, D.; Gao, Q.; Liu, J.; Tang, J.; Hua, Z.; Sun, X. Categories of exogenous substances and their effect on alleviation of plant salt stress. Eur. J. Agron. 2023, 142, 126656. [Google Scholar] [CrossRef]
- Souza, A.C.; Zandonadi, D.B.; Santos, M.P.; Canellas, N.O.A.; Soares, C.D.; Irineu, L.E.S.D.; de Rezende, C.E.; Spaccini, R.; Piccolo, A.; Olivares, F.L.; et al. Acclimation with humic acids enhances maize and tomato tolerance to salinity. Chem. Biol. Technol. Agric. 2021, 8, 40. [Google Scholar] [CrossRef]
- Mora, V.; Bacaicoa, E.; Zamarreno, A.; Aguirre, E.; Garnica, M.; Fuentes, M.; García-Mina, J. Action of humic acid on promotion of cucumber shoot growth involves nitrate-related changes associated with the root-to-shoot distribution of cytokinins, polyamines and mineral nutrients. J. Plant Physiol. 2010, 167, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Shukry, W.M.; Abu-Ria, M.E.; Abo-Hamed, S.A.; Anis, G.B.; Ibraheem, F. The efficiency of humic acid for improving salinity tolerance in salt sensitive rice (Oryza sativa): Growth responses and physiological mechanisms. Gesunde Pflanz. 2023, 75, 2639–2653. [Google Scholar] [CrossRef]
- Hatami, E.; Shokouhian, A.A.; Ghanbari, A.R.; Naseri, L.A. Alleviating salt stress in almond rootstocks using of humic acid. Sci. Hortic. 2018, 237, 296–302. [Google Scholar] [CrossRef]
- Saidimoradi, D.; Ghaderi, N.; Javadi, T. Salinity stress mitigation by humic acid application in strawberry (Fragaria x ananassa duch.). Sci. Hortic. 2019, 256, 108594. [Google Scholar] [CrossRef]
- Traversa, A.; Loffredo, E.; Gattullo, C.E.; Palazzo, A.J.; Bashore, T.L.; Senesi, N. Comparative evaluation of compost humic acids and their effects on the germination of switchgrass (Panicum vigatum L.). J. Soils Sediments 2014, 14, 432–440. [Google Scholar] [CrossRef]
- Tóth, Š.; Šoltysová, B.; Dupľák, Š.; Porvaz, P. Impact of Soil-Applied Humic Ameliorative Amendment on the Ligno-Cellulose Quality and Calorific Value of Switchgrass Panicum virgatum L. Agronomy 2023, 13, 1854. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, Q.; Yang, Z.; Yan, M.; Wu, W.; Wang, Y. Chemical characteristics and bioactivity of humic acids from weathered coals with different sources. Chin. J. Soil Sci. 2022, 53, 540–547. [Google Scholar] [CrossRef]
- Niu, Y.; Lyu, H.; Liu, X.; Zhang, M.; Li, H. Effects of supplemental lighting duration and matrix moisture on net photosynthetic rate of tomato plants under solar greenhouse in winter. Comput. Electron. Agric. 2022, 198, 107102. [Google Scholar] [CrossRef]
- Lahijanian, S.; Eskandari, M.; Akhbarfar, G.; Azizi, I.; Afazel, M.; Ghobadi, C. Morphological, physiological and antioxidant response of stevia rebaudiana under in vitro agar induced drought stress. J. Agric. Food Res. 2023, 11, 100495. [Google Scholar] [CrossRef]
- Katam, R.; Sakata, K.; Suravajhala, P.; Pechan, T.; Kambiranda, D.M.; Naik, K.S.; Guo, B.; Basha, S.M. Comparative leaf proteomics of drought-tolerant and -susceptible peanut in response to water stress. J. Proteom. 2016, 143, 209–226. [Google Scholar] [CrossRef]
- Alharby, H.F. Using some growth stimuli, a comparative study of salt tolerance in two tomatoes cultivars and a related wild line with special reference to superoxide dismutases and related micronutrients. Saudi J. Biol. Sci. 2021, 28, 6133–6144. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Rengel, Z. Waterlogging influences plant growth and activities of superoxide dismutases in narrow-leafed lupin and transgenic tobacco plants. J. Plant Physiol. 1999, 155, 431–438. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Mehri, H.; Ghobadi, C.; Baninasab, B.; Afazel, M. Morphological, physiological and antioxidant responses of some Iranian grapevine cultivars to methyl jasmonate application. J. Crop Sci. Biotechnol. 2013, 16, 277–283. [Google Scholar] [CrossRef]
- Sudhakar, C.; Lakshmi, A.; Giridarakumar, S. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) Under NaCl salinity. Plant Sci. 2001, 161, 613–619. [Google Scholar] [CrossRef]
- Li, X.; Sun, P.; Zhang, Y.; Jin, C.; Guan, C. A novel PGPR strain Kocuria rhizophila y1 enhances salt stress tolerance in maize by regulating phytohormone levels, nutrient acquisition, redox potential, ion homeostasis, photosynthetic capacity and stress-responsive genes expression. Environ. Exp. Bot. 2020, 174, 104023. [Google Scholar] [CrossRef]
- Ariani, A.; Gepts, P. Genome-wide identification and characterization of aquaporin gene family in common bean (Phaseolus vulgaris L.). Mol. Genet. Genom. 2015, 290, 1771–1785. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; He, T.; Huang, C.; Wang, K.; Shi, D.; Si, X.; Xu, Y.; Lyu, S.; Huang, J.; Li, Y. Genome-wide identification of KCS gene family in Carya illinoinensis and their roles under abiotic stress conditions. Sci. Hortic. 2023, 321, 112343. [Google Scholar] [CrossRef]
- Popescu, G.C.; Popescu, M. Yield, berry quality and physiological response of grapevine to foliar humic acid application. Bragantia 2018, 77, 273–282. [Google Scholar] [CrossRef]
- Chen, Q.; Qu, Z.; Ma, G.; Wang, W.; Dai, J.; Zhang, M.; Wei, Z.; Liu, Z. Humic acid modulates growth, photosynthesis, hormone and osmolytes system of maize under drought conditions. Agric. Water Manag. 2022, 263, 107447. [Google Scholar] [CrossRef]
- Li, N.; Zhang, Z.; Gao, S.; Lv, Y.; Chen, Z.; Cao, B.; Xu, K. Different responses of two Chinese cabbage (Brassica rapa L. ssp. pekinensis) cultivars in photosynthetic characteristics and chloroplast ultrastructure to salt and alkali stress. Planta 2021, 254, 102. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Turner, N.C. Turgor maintenance by osmotic adjustment: 40 years of progress. J. Exp. Bot. 2018, 69, 3223–3233. [Google Scholar] [CrossRef]
- El Nahhas, N.; AlKahtani, M.D.F.; Abdelaal, K.A.A.; Al Husnain, L.; AlGwaiz, H.I.M.; Hafez, Y.M.; Attia, K.A.; El-Esawi, M.A.; Ibrahim, M.F.M.; Elkelish, A. Biochar and jasmonic acid application attenuates antioxidative systems and improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) Irrigated with saline water. Plant Physiol. Biochem. 2021, 166, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Lee, I. Regulation of salicylic acid, jasmonic acid and fatty acids in cucumber (Cucumis sativus L.) by spermidine promotes plant growth against salt stress. Acta Physiol. Plant. 2013, 35, 3315–3322. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M.R. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, V.P.; Lima, M.D.R.; da Silva, B.R.S.; Batista, B.L.; Lobato, A.K.D. Brassinosteroids confer tolerance to salt stress in eucalyptus urophylla plants enhancing homeostasis, antioxidant metabolism and leaf anatomy. J. Plant Growth Regul. 2019, 38, 557–573. [Google Scholar] [CrossRef]
- Yan, H.; Li, Q.; Park, S.; Wang, X.; Liu, Y.; Zhang, Y.; Tang, W.; Kou, M.; Ma, D. Overexpression of CuZnSOD and APX enhance salt stress tolerance in sweet potato. Plant Physiol. Biochem. 2016, 109, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ye, N.; Liu, R.; Chen, M.; Zhang, J. H2O2 mediates the regulation of aba catabolism and GA biosynthesis in Arabidopsis seed dormancy and germination. J. Exp. Bot. 2010, 61, 2979–2990. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Song, L.; Gong, X.; Xu, J.; Li, M. Functions of jasmonic acid in plant regulation and response to abiotic stress. Int. J. Mol. Sci. 2020, 21, 1446. [Google Scholar] [CrossRef]
- Yoon, Y.; Seo, D.H.; Shin, H.; Kim, H.J.; Kim, C.M.; Jang, G. The role of stress-responsive transcription factors in modulating abiotic stress tolerance in plants. Agronomy 2020, 10, 788. [Google Scholar] [CrossRef]
- Hachez, C.; Besserer, A.; Chevalier, A.S.; Chaumont, F. Insights into plant plasma membrane aquaporin trafficking. Trends Plant Sci. 2013, 18, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Li, R.; Wang, J.; Wu, X.; Zhang, N.; Sun, Q.; Qi, Y.; Xing, Y.; Zhou, C.; Zhao, B.; et al. Cloning and sequence analysis of the plasma membrane aquaporins gene PvPIP1 in switchgrass. Acta Agrestia Sin. 2014, 22, 840–846. [Google Scholar] [CrossRef]
- de Ollas, C.; Dodd, I.C. Physiological impacts of ABA–JA interactions under water-limitation. Plant Mol. Biol. 2016, 91, 641–650. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of salt and humic acid on plant growth (A), net photosynthetic rate (B), transpiration rate (C), stomatal conductance (D) and intercellular carbon dioxide content (E) of switchgrass. Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
Figure 1.
Effects of salt and humic acid on plant growth (A), net photosynthetic rate (B), transpiration rate (C), stomatal conductance (D) and intercellular carbon dioxide content (E) of switchgrass. Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
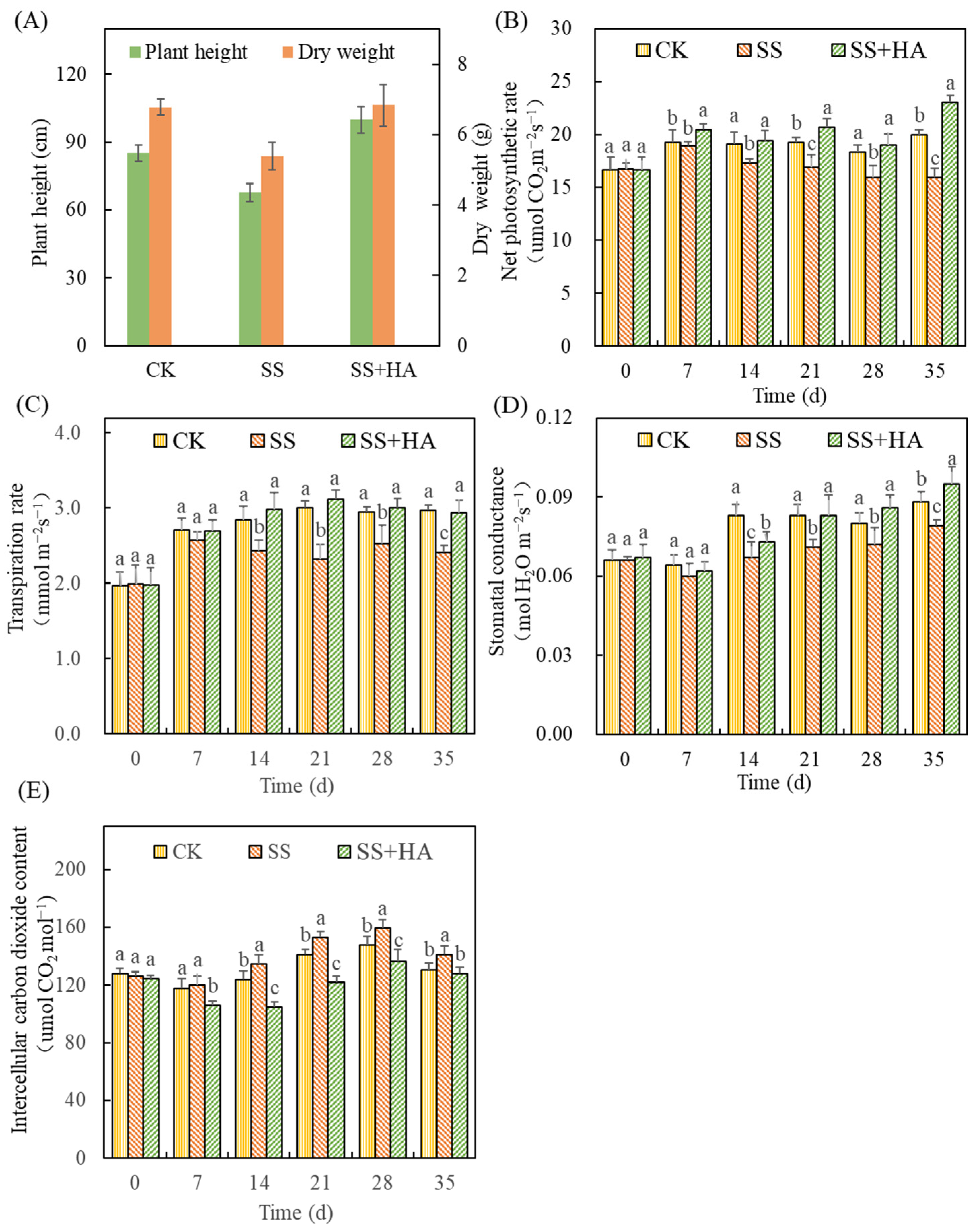
Figure 2.
Correlation analysis of growth performance and photosynthetic function of switchgrass.

Figure 3.
Effects of salt and humic acid on physiological indexes of switchgrass. Relative electrolyte leakage rate of blade (A), proline (B), malondialdehyde (C), superoxide dismutase (D), peroxidase (E), and ascorbic acid peroxidase (F). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
Figure 3.
Effects of salt and humic acid on physiological indexes of switchgrass. Relative electrolyte leakage rate of blade (A), proline (B), malondialdehyde (C), superoxide dismutase (D), peroxidase (E), and ascorbic acid peroxidase (F). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.

Figure 4.
Effect of salt and humic acid on phytohormones of switchgrass. Concentrations of abscisic acid (A), polyamine (B), cytokinin (C), indoleacetic acid (D), brassinolide (E), and jasmonic acid (F). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
Figure 4.
Effect of salt and humic acid on phytohormones of switchgrass. Concentrations of abscisic acid (A), polyamine (B), cytokinin (C), indoleacetic acid (D), brassinolide (E), and jasmonic acid (F). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
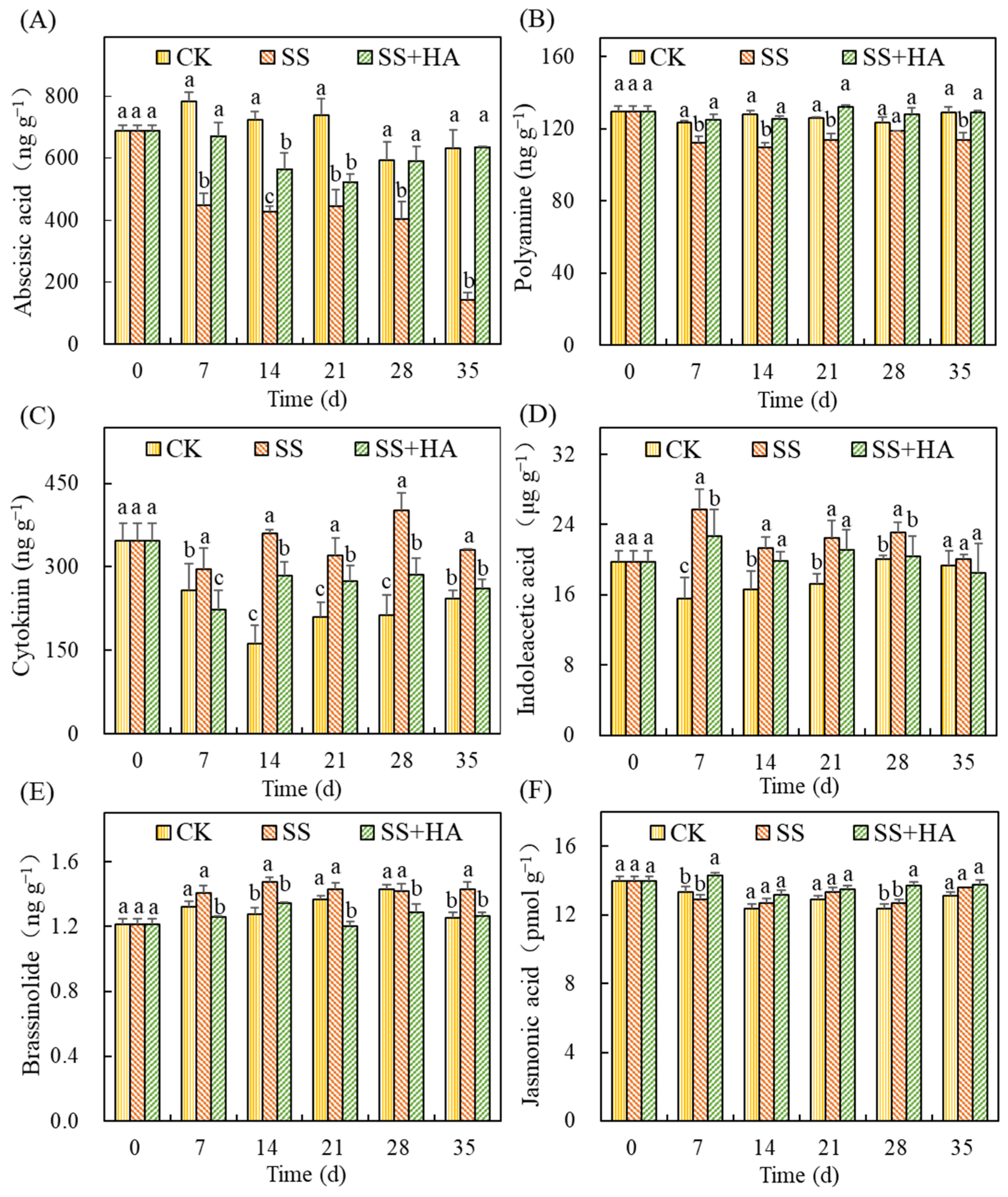
Figure 5.
Relative expression of the PvPIP1 gene in switchgrass with the addition of humic acid under salt stress (A); the correlation between PvPIP1 expression with photosynthetic indexes and membrane stability in switchgrass (B). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.
Figure 5.
Relative expression of the PvPIP1 gene in switchgrass with the addition of humic acid under salt stress (A); the correlation between PvPIP1 expression with photosynthetic indexes and membrane stability in switchgrass (B). Treatments with the same letters for each sampling date are not significantly different at p = 0.05.

Figure 6.
Redundancy analysis between physiological index and photosynthetic indexes of switchgrass (A); redundancy analysis between phytohormones and photosynthetic indexes of switchgrass (B) with the addition of humic acid under salt stress.
Figure 6.
Redundancy analysis between physiological index and photosynthetic indexes of switchgrass (A); redundancy analysis between phytohormones and photosynthetic indexes of switchgrass (B) with the addition of humic acid under salt stress.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, J.; Meng, Q.; Yang, Z.; Zhang, Q.; Yan, M.; Hou, X.; Zhang, X. Humic Acid Promotes the Growth of Switchgrass under Salt Stress by Improving Photosynthetic Function. Agronomy 2024, 14, 1079. https://doi.org/10.3390/agronomy14051079
AMA Style
Zhang J, Meng Q, Yang Z, Zhang Q, Yan M, Hou X, Zhang X. Humic Acid Promotes the Growth of Switchgrass under Salt Stress by Improving Photosynthetic Function. Agronomy. 2024; 14(5):1079. https://doi.org/10.3390/agronomy14051079
Chicago/Turabian StyleZhang, Jiaxing, Qiuxia Meng, Zhiping Yang, Qiang Zhang, Min Yan, Xiaochan Hou, and Xunzhong Zhang. 2024. "Humic Acid Promotes the Growth of Switchgrass under Salt Stress by Improving Photosynthetic Function" Agronomy 14, no. 5: 1079. https://doi.org/10.3390/agronomy14051079
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.