
Root Respiration–Trait Relationships Are Influenced by Leaf Habit in Tropical Plants
Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, College of Tropical Agriculture and Forestry, Hainan University, Haikou 570228, China
*
Author to whom correspondence should be addressed.
Forests 2024, 15(5), 806; https://doi.org/10.3390/f15050806
Submission received: 28 March 2024
/
Revised: 23 April 2024
/
Accepted: 30 April 2024
/
Published: 2 May 2024
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:Root respiration is a critical physiological trait that significantly influences root system activity. Recent studies have associated root respiration with the economic functioning of roots; however, research on root respiration in tropical plants remains limited. This study examined the fine root respiration and the relationship between root respiration and root chemical and morphological traits in 16 tropical plant species, including both evergreen and deciduous species. Findings revealed that deciduous species exhibited higher root respiration compared to evergreen species. Root respiration positively correlated with root nitrogen concentration and specific root length and correlated negatively with root diameter and root tissue density across all species. The root respiration patterns in evergreen species aligned with those seen in all tree species, while deciduous species showed a distinct negative correlation with root tissue density and no significant correlations with other root traits. Principal component analysis revealed that the patterns of root variation in both evergreen and deciduous trees were multidimensional, with deciduous trees exhibiting acquisitive traits and evergreen trees displaying conservative traits. Random forest and multiple regression analysis showed that specific root length exerted the most significant influence on root respiration in both evergreen and deciduous trees. These findings are ecologically significant, enhancing our understanding of root respiration in tropical plants and its impact on ecosystem functions. They contribute valuable insights and support the conservation and management of tropical vegetation.
1. Introduction
Approaches based on plant functional traits are often considered the best way to explore ecosystem processes and functions [1,2]. Plant functional traits are sensitive to environmental change and influence ecosystem carbon and nutrient cycling [3,4]. Plant aboveground traits therefore serve as key indicators of resource acquisition strategy that are easy to observe and measure [5]. Given the pivotal role of leaf functional traits, it is widely recognized that terrestrial plants exhibit a trade-off phenomenon, which is called leaf economics spectrum [6,7]. Similarly, plant roots support carbon, nutrient, and water cycling to facilitate plant growth and moderate below-ground ecosystem processes [8]. Within the root system, fine roots (≤2 mm in diameter) are the most metabolically active and their primary productivity accounts for 22% of the total global terrestrial [9,10]. Due to the multifunctionality of roots, plants develop different combinations of morphological, chemical, and physiological traits of root systems to acquire nutrients and adapt to the environment [3]. A root economics spectrum (RES) hypothesis has been proposed to describe fine root acquisition patterns and how these patterns respond to environmental change [11].
RES has received more attention in studies concerning plant resource acquisition strategies. In several studies, the RES has been found to exhibit one-dimensional characteristics, with a negative correlation between root nitrogen concentration (RNC) and root tissue density (RTD) [11,12]. However, the bulk of the evidence suggests that root trait syndromes are two-dimensional or multi-dimensional [13,14]. As reported by Kong et al. [15], root diameter (RD) and branching architecture are two distinct primary dimensions of root trait co-variation. The traits linked with mycorrhizal associations (e.g., specific root length [SRL], RD) have been verified to be independent of the traits associated with the classical RES [16]. Compared with chemical and morphological traits, root physiological traits (e.g., root respiration and root exudates rate) are often neglected in previous studies of the covariation of multivariate root traits [11,17]. Therefore, it remains uncertain whether root physiological traits employ comparable acquisition strategies to other root traits. Clarifying whether root physiological trait correlations in tropical plants conform to the RES will provide new insights into root functional relationships.
Fine root respiration is a crucial physiological trait as it provides energy for ion uptake, synthesis, and the maintenance of root biomass and is associated with carbon acquisition capacity [11,18]. How root systems impact plant growth and ecosystem processes is closely tied to the interplay among fine root chemical, morphological, and physiological traits [3]. Although root respiration shapes nutrient acquisition and plays a vital role in the RES [18,19], previous studies testing the RES hypothesis predominantly focused only on chemical and morphological traits, given the relative difficulty of measuring respiration [20,21,22,23]. But there are also several studies in grassland, temperate forest, and subtropical forest ecosystems that incorporate root respiration into the RES. For example, Reich et al. [24] and Han and Zhu [19] studied the relationships of root traits and showed that the root respiration–SRL relationships are weak or even insignificant, which are not completely in line with predictions by the RES. However, in temperate regions, a significant negative relationship between root respiration and SRL was found [17,25]. It is evident that controversy exists regarding the correlation between root respiration and other root traits based on the RES. Therefore, investigating how root respiration is related to other root traits in tropical plant species should enhance our knowledge on the RES and root nutrient acquisition strategies in tropical regions [11,17].
Plant leaf habits (deciduous and evergreen) are an essential classification method for understanding the co-variation of root traits and the ecological effects of their interrelationships [19,26]. Previous studies showed that leaf habit exhibits different effects on root traits. While a study indicated that leaf habits do not consistently correlate with root respiration [27], there are also studies indicating that deciduous and evergreen species can directly or indirectly affect root respiration by influencing other root traits [19,28]. Studies in subtropical regions also showed that deciduous species cluster on the acquisitive side with higher root respiration, RNC, and SRL, whereas evergreen species cluster on the conservative side with lower root respiration, SRL, and RNC, indicating that deciduous and evergreen species possess different root traits [19,29]. Evergreen and deciduous species are common leaf habits in forest ecosystems from temperate to tropical biomes [29,30]. They exhibit different leaf habit traits that can affect leaf photosynthetic capacity, nutrient allocation, and productivity in forest ecosystem processes [31,32]. For example, deciduous species allocate more resources to roots, while evergreens allocate more biomass to leaves and stems [33]. Whether root traits of deciduous and evergreen species in tropical regions display the same resource strategies as those in temperate or subtropical regions remains uncertain.
In this study, 16 representative tropical plant species were selected in a botanical garden of Hainan Province. We selected these plant species based on their widespread presence in tropical regions, encompassing a majority of the family, their demonstration of consistent growth patterns, and not being categorized as rare, protected tree species. The study hypothesized that (1) plant leaf habit influences root respiration in tropical plants, (2) the associations between root respiration and root morphological and chemical traits vary among leaf habits, and (3) the root respiration–trait relationships in tropical plants align with the one-dimensional RES, regardless of leaf habit. To test these hypotheses, the fine root respiration rate, as well as a number of root morphological traits (e.g., RD, RTD, and SRL) and chemical traits (e.g., root carbon concentration [RCC], RNC, and the root C/N ratio), were measured for all 16 tropical plant species. The study aimed to quantify the fine root respiration rate and its relationships with other root traits, and to examine the effects of plant leaf habit on these relationships in tropical plants. The results will be useful in understanding the root nutrient acquisition strategies of tropical plants in the conditions of different leaf habits.
2. Materials and Methods
2.1. Study Site Information
The study site was located in Hainan Tropical Botanical Garden (19°52′ N, 109°50′ E) in Danzhou, Hainan Province, China (Figure 1). Danzhou is situated on the southern edge of the monsoon climate in East Asia, within the tropical zone. Thus, the site experiences both dry and wet seasons each year. The wet season extends from May to October, while the dry season lasts from November to April of the following year. The mean annual temperature ranges from 21.5 to 28.5 °C, and the mean annual rainfall is 1815 mm. Soils are latosols, with an average pH of about 5.2. The Hainan Tropical Botanical Garden holds significant tropical plant resources in China, covering 32 hectares. There are more than 1000 tropical species planted in the arboretum, mainly belonging to the Arecaceae, Moraceae, and Fabaceae families, including species such as Artocarpus heterophyllus Lam., Roystonea regia (Kunth) O.F. Cook, and Pterocarpus indicus Willd.
2.2. Root Sample Collection
In August 2022, 16 tropical woody plants, representing a broad range of tropical taxa including some relatively common species, were selected for study in Hainan Tropical Botanical Garden. Plant species were distributed randomly in the sampling area. Based on leaf habit, these species were categorized as deciduous and evergreen species (Table 1). For each plant species, five matured and healthy individuals of a similar size were selected as replicates for fine root collection. Before sampling root samples, the litter layer was removed. At a distance of 1–2 m from the tree trunk/shrub stem, we identified coarse roots that could be traced back to the target tree/shrub. Then a 40 × 40 × 40 cm soil profile was dug along with the coarse roots and picked out fine roots (diameter < 2 mm) attached to the coarse roots. The fine roots were carefully cut off using sterilized scissors. Any dead roots exhibiting a dark brown or black color with frayed edges were discarded. For each target individual, fine roots were sampled from four radial directions. Subsequently, the root samples obtained from the four directions were mixed to create one composite sample. A total of 80 samples were collected. All samples were placed in sterile bags, stored in a cooler box, and quickly transported (within 10 min) to the laboratory for root respiration and functional trait analysis.
2.3. Root Respiration Rate
Before measurement, the soil was carefully removed from the roots using sterilized brushes. The root respiration rate was measured using the method proposed by Roumet et al. [11]. Briefly, root samples were put into sealed jars (c. 250 mL) for incubation at 25 °C. After an hour, the gas samples were collected using a 50 mL syringe to assess the CO2 concentration in each jar before sealing the lid. Additional gas samples were collected after one hour. All gas samples were injected into aluminum foil gas-collecting bags (c. 50 mL) for later analysis. The CO2 concentration was analyzed for each sample using a Delta V Advantage isotope ratio mass spectrometer (IRMS; Thermo Fisher Scientific, Bremen, Germany). The root respiration rate was calculated as the amount of CO2 released by the root system in one hour divided by the collection time (1 h) and root dry mass.
2.4. Root Morphological and Chemical Traits
After the root respiration measurements were completed, root morphological and chemical traits were assessed. Root samples were scanned on an Epson Perfection v850 Pro scanner (Epson Perfection v850 Pro; Epson, Ontario, Canada) and images were analyzed in WinRHIZO Pro 2020a (Regent Instruments Inc., Quebec City, QC, Canada). The average root diameter (RD, mm), length (RL, cm), and volume (RV, cm3) were obtained. After measurement, root samples were dried to a constant weight at 65 °C to obtain the root dry weight (RDW, g). The root tissue density (RTD, g cm−3) and specific root length (SRL, m g−1) were calculated as follows:
The root carbon concentrations (RCC, mg g−1) and root nitrogen concentrations (RNC, mg g−1) were measured using an elemental analyzer (Elementar vario MACRO cube, Elementar Analysensysteme GmbH, Hanau City, Germany). The root C/N ratio was calculated.
2.5. Statistical Analysis
Before analysis, the data were tested for normality and homoscedasticity using Kolmogorov–Smirnov and Levene’s tests, respectively, and the data that did not fit normal distribution were log or sqrt transformed. Differences across all plant species were tested using one-way analysis of variance (ANOVA) with Tukey’s honest significant difference (HSD) test. A Student’s t-test was used to compare the differences in root traits between deciduous and evergreen species. In order to evaluate the phylogenetic influence on root traits, a plant phylogenetic tree was constructed using a phylogeny pruned from Zanne 2014 in Phylomatic version 3.0 (accessed on 21 November 2023, http://phylodiversity.net/phylomatic). We calculated Blomberg’s K and Pagel’s λ in ‘phytools’ R package to evaluate the phylogenetic signal for root respiration and root traits [34,35]. Linear regression was used to describe the relevance of root respiration and the root morphological and chemical traits. Pearson correlations were calculated among the root respiration rate, root morphological, and chemical traits. A principal component analysis (PCA) was conducted using the root traits for all species, deciduous species, and evergreen species. To compare the relative importance of root morphological and chemical traits for root respiration rate variation across species and plant leaf habit, random forest models were implemented using the randomForest package in R. The importance of a factor was calculated as the increase in the mean square error (%IncMSE). After checking potential collinearity, the multiple regression analysis was used to describe the importance of root morphological and chemical traits to root respiration rate. All analyses were conducted in R 4.0.3 [36]. Significance was assigned to effects with p < 0.05.
3. Results
3.1. Root Respiration Rate Variation among Plant Species and Leaf Habits
The root respiration rate across 16 plant species ranged from 0.99 ± 0.10 (mean ± SE) to 4.61 ± 1.37 nmol CO2 g−1 s−1, representing a 4.6-fold difference (Figure 2A). Among plant species, Cocos nucifera L., Murraya paniculata (L.) Jacq., Litchi chinensis Sonn., and P. indicus showed significant differences in root respiration rate, and P. indicus possessed the highest respiration rate (p < 0.05). The root respiration varied from 2.17 ± 0.60 to 4.61 ± 1.37 nmol CO2 g−1 s−1 for deciduous species and it ranged from 0.99 ± 0.10 to 3.66 ± 0.64 nmol CO2 g−1 s−1 for evergreen species. The deciduous species exhibited significantly higher root respiration than the evergreen species (p = 0.04, Figure 2B). Plant leaf habit significantly affected root chemical traits (Table 2). The RNC was higher in deciduous species than evergreen species, while the C/N and RCC were lower in deciduous tree species (Table 2, p < 0.05).
3.2. Correlations among Root Respiration Rate and Other Root Traits
A plant phylogenetic tree was conducted to assess its influence on root traits (Figure S1). Blomberg’s K and Pagel’s λ test showed that root respiration and root traits were not influenced by plant phylogeny among the 16 plant species (Table S1). Across all 16 study species, the RNC and SRL positively correlated with the root respiration rate (Figure 3A,E, p < 0.001), while the RD, RTD, and C/N ratio negatively correlated with the root respiration rate (Figure 3B,C, p < 0.001). The relevance between root respiration and other root traits was affected by leaf habit (Figure 3). Root respiration in evergreen species showed a positive correlation with the RNC and SRL and a negative correlation with the C/N, RD, and RTD (p < 0.001). But the root respiration was only negatively correlated with the RTD in deciduous species (Figure 3).
Pairwise trait associations were assessed using Pearson correlation coefficients (Figure 4). Across all species, root chemical and morphological traits correlated inconsistently with the hypothesis of the RES, as the RD did not correlate with the RNC (R = −0.13, p > 0.05) and RTD (R = −0.11, p > 0.05). Similarly, the RNC was not correlated with the SRL and RTD (R = 0.17, R = −0.13, p > 0.05) (Figure 4A). Deciduous and evergreen species affected these relationships (Figure 4B,C). In deciduous species, the RD was positively correlated with the RNC (R = 0.71, p < 0.001) but not with the RTD (R = −0.29, p > 0.05) and the SRL was negatively correlated with the RNC (R = −0.50, p = 0.026, Figure 4B). In evergreen species, the RNC was negatively associated with the RD (R = −0.51, p < 0.001) and SRL (R = 0.47, p < 0.001), but not associated with the RTD (R = −0.13, p > 0.05, Figure 4C). The PCA results were consistent with the correlation analysis, revealing that root trait variation was not one-dimensional. Across all plant species, two main trait variation axes were observed, accounting for 40.8% and 22.3% of the total variation, respectively (Figure 5A, Table S2). On the first PCA axis, the RNC, root respiration, RD, and SRL had high scores, while the RTD and RCC exhibited a high score on the second axes (Table S2). Two variation axes were observed in the PCAs for evergreen and deciduous species (Figure 5B,C, Table S2). Specifically, compared with evergreen species, deciduous species are located at the high end of the RNC, SRL, and root respiration and lower RD (Figure 5A and Figure S2, Table S2).
3.3. Effects of Root Traits on Root Respiration Rate
Random forest models were used to analyze the effects of root chemical and morphological traits on root respiration (Figure 6). Considering all species, the root respiration rate of species was mainly affected by the SRL (%IncMSE = 16.485, p = 0.009), RNC (%IncMSE = 12.185, p = 0.009), and RD (%IncMSE = 10.647, p = 0.019), and the most influential factor for root respiration was the SRL (Figure 6A). The deciduous and evergreen species affected the importance of each root trait for root respiration. In deciduous and evergreen species, the most influential factor for root respiration was the SRL (evergreen species (%IncMSE = 13.403, p = 0.009), deciduous species (%IncMSE = 13.030, p = 0.009), Figure 6B,C). Multiple regression analysis also showed that the most important factor for the root respiration rate was the SRL among all species, deciduous species, and evergreen species (Table 3).
4. Discussion
4.1. Effects of Plant Leaf Habit on Root Respiration and Root Respiration–Trait Relationships in Tropical Plants
Root respiration rates significantly differed among the plant species studied (i.e., C. nucifera, M. paniculata, L. chinensis, and P. indicus), and leaf habit (deciduous and evergreen species) had significant effects on root respiration rates (Figure 2A,B). These results supported our first hypothesis. In a recent review of root respiration in 245 plant species in global temperate forests, deciduous and evergreen species rather than other leaf habits affect root respiration [19]. Similarly, the root respiration of deciduous was higher than evergreen species in the present study (Figure 2B). The following possibilities might explain this result. First, deciduous species clustered on the acquisitive side with higher root respiration, RNC, and SRL, whereas evergreen species clustered on the conservative side with lower root respiration, RNC, and SRL (Figure S2). Furthermore, leaf respiration of deciduous species is weakened after leaf withering, while root respiration will accelerate to support normal plant growth [37]. Additionally, in the study, leguminous plants (e.g., P. indicus, Delonix regia (Bojer ex Hook.) Raf) were included in deciduous species; they have root systems that exhibit higher respiratory carbon costs to support N2 fixation compared with other plants [38], thereby resulting in higher root respiration than evergreen species.
As hypothesized, the relationships between root respiration and root traits in tropical species were influenced by plant leaf habit. In evergreen species, the root respiration exhibited a positive correlation with the RNC and SRL and a negative correlation with the RD, RTD and C/N, while the root respiration rate was only negatively correlated with the RTD in deciduous species (Figure 3). In general, RNC and SRL are closely related to root respirations [11,39,40]. Similarly, our results showed that SRL was an important trait to determine the root respiration regardless of leaf habit (Figure 6, Table 3 and Table S3). Nitrogen is a major factor limiting plant growth and metabolism [41]. All life activities depend on the participation of nitrogen elements, including enzyme synthesis, which is closely related to root respiration [24]. SRL, representing the ratio of length to weight of a root segment, is a crucial parameter for evaluating nutrient uptake potential [24] and is known to be associated with root respiration [39]. However, there were no correlations between the root respiration and RNC and SRL in deciduous species (Figure 3). According to the results, although the root respiration of deciduous species exhibited a positive trend with the RNC and SRL, there is no significant relationships (Figure 3A,E). This might be attributed to the limited sample number of deciduous species. Prior studies have indicated that the number of species can influence the association between root respiration and other traits [11,25]. Moreover, the research was conducted in a botanical garden, where the sampled plants have the same precipitation, temperature, and topography. But the characteristics of plants will affect the rhizosphere environment. For example, legumes species can develop symbiotic root nodules with nitrogen-fixing bacteria to ensure nitrogen availability for growth [42]. It has been proven that the development of nitrogen-fixing nodules on leguminous plant roots impacts the root morphological characteristics, and the symbiotic relationship between root nodules and roots may partly substitute resource uptake compared to the root length [43]. Additionally, leguminous plants contain high proportions of insoluble nitrogen, which is primarily utilized for constructing structural components like cell walls and exhibits low metabolic activity [44]. It deviates from the anticipated positive correlation between nitrogen concentration and metabolic activity [24,41]. In our study, half of deciduous tree species are members of the Leguminosae family (Table 1). This might result in the absence of a significant correlation between the root respiration and SRL and RNC in deciduous species. Testing how the relationship between root respiration and other root morphological and chemical traits changes between deciduous and evergreen species will help us understand the underground resource acquisition strategies of plants with different leaf habits.
4.2. Tropical Plant Root Traits Followed a Multidimensional Pattern of Variation Rather Than a One-Dimensional RES
In deciduous species, the RNC was negatively correlated with the RTD (Figure 4B), demonstrating that deciduous species employed a trade-off strategy when utilizing resources. This implies that increasing investment in one particular trait resource by plants will result in a corresponding reduction in investments in other trait resources [6]. Plants can efficiently achieve optimal allocation of resources by adjusting the balance among different traits [6,11]. Pearson correlation analysis showed that there was no significant correlation between the RTD and RNC in evergreen species in the study (Figure 4C). However, the PCA results showed that samples in the RNC and RTD space were distributed evenly (Figure 5C), indicating there might be two types of trade-offs in evergreen trees: some trees had a high RNC and low RTD, while others had a high RTD and low RNC. This caused a trade-off offset in evergreen trees in the Pearson correlation analysis and exhibited no significant negative correlation. But the trade-off between the RNC and RTD existed in evergreen species. Additionally, a trade-off relationship between the SRL and RD was exhibited, regardless of leaf habit (Figure 4). This finding aligns with previous research indicating that species with a higher SRL tend to exhibit finer RD [45,46]. This root architecture enhances the capacity of plants to effectively uptake resources, demonstrating the trade-off relationship among key traits involved in resource acquisition in plants [11,45]. The PCAs revealed that deciduous species are located at the high end of the RNC, SRL, and root respiration, and lower RD compared to evergreen species (Figure 5 and Figure S2, Table S2). Although our experiment was conducted in a garden with the same growth environment, the results demonstrated that deciduous and evergreen species adjust the relationship between root functional traits to optimize the efficiency of resource acquisition and utilization, forming ecological strategies that are more suitable for survival in the environment.
As expected, the PCA results indicated that tropical plant root traits variation was multidimensional rather than one-dimensional, regardless of leaf habit (Figure 5, Table S2). Across all species, root respiration, RNCs, and SRLs were negatively correlated with PC1, while RDs were positively correlated with PC1; the RTD was most strongly correlated with PC2, while RCCs were negatively correlated with PC2 (Figure 5A, Table S2). These findings were consistent with previous research suggesting the multidimensional variability of roots [15,17,47]. Changes in soil conditions can result in the multidimensionality of root trait variation [47,48,49]. In high soil fertility environments, plant RTD and root branching are reduced, along with an increase in SRL and RD [47]. In subtropical regions, the RNC and root respiration increase with rising soil temperatures [49,50]. Despite the fact that our experiment was conducted in a botanical garden where climatic and topographical conditions were uniform, the different quantity and quality of litters provided by individual plant species’ input to soil result in divergent physicochemical properties [51,52]. Therefore, plant root traits must diversify and form various combinations to effectively acquire resources from complex soil environments. Indeed, observations from a wide range of species indicated that plants have the capacity to develop roots featuring various combinations of SRL and RTD [47]. This complexity in the interaction of root traits in tropical plants challenges the notion of a one-dimensional RES [47,53].
Notably, only plants with arbuscular mycorrhizal associations were examined in this study due to the tropical woody plants are dominated by arbuscular mycorrhizal species [54,55]. However, root resource acquisition strategies are also influenced by mycorrhizal type [23,56]. Mycorrhizal type can impact the relationships between fine root chemical (e.g., RNC) or morphological (e.g., SRL) traits and root respiration [57]. Distinguishing mycorrhizal types can provide a more detailed understanding of plant resource acquisition strategies and ecosystem dynamics in tropical regions. Therefore, mycorrhizal types would be considered in our future studies.
5. Conclusions
The present study examined root respiration and the associations between root respiration and the chemical and morphological characteristics of roots in tropical plant species. Our results showed that deciduous and evergreen species impact root respiration and its correlation with other root traits, suggesting that leaf habit will influence root resource acquisition strategies by changing the relationships among root physiological, chemical, and morphological traits. The SRL was the most important trait affecting root respiration, regardless of leaf habit. The PCA results indicated that deciduous species exhibited an acquisition strategy, while evergreen species showed a conservative strategy. Furthermore, although the plants selected in the experiment were grown in the same climatic and topographical environment, the results supported the notion that root respiration and other root traits are multi-dimensional patterns of variation rather than one-dimensional RES predictions, regardless of leaf habit. The varied resource acquisition strategies among species may promote the coexistence of different species in the same environment. These findings enhance our comprehension of the variability in root respiration among tropical plants, the significance of root respiration in the root economics spectrum, and the diverse strategies for root resource acquisition.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/f15050806/s1: Figure S1: Phylogeny trees mapped onto each of the 16 species in this study; Figure S2: Results of independent- samples t- test for elucidating the differences between deciduous and evergreen species along PC1 and PC2; Table S1: The test of the phylogenetic signal of the root traits, including Blomberg’s K and Pagel’s λ.; Table S2: Results of the PCA on root morphological traits (e.g., SRL, RD, and RTD), chemical traits, and respiration rate for the 16 study species and deciduous with evergreen; Table S3: The relationships between root respiration and the co-variation in root traits (PC1 and PC2).
Author Contributions
Methodology, data curation, and writing—original draft, D.D.; methodology, investigation, and validation, D.D. and M.Y.; resources and funding acquisition, Y.S.; writing—review and editing, Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Hainan Provincial Natural Science Foundation of China (No. 421QN193, No. 423RC428), the National Natural Science Foundation of China (No. 32201629), and Hainan University (No. KYQD(ZR)-21038).
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We would like to thank Daniel Petticord at the University of Cornell for his assistance with English language and grammatical editing of the manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Faucon, M.P.; Houben, D.; Lambers, H. Plant Functional Traits: Soil and Ecosystem Services. Trends Plant Sci. 2017, 20, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Sun, L.; Gan, D.; Fu, L.; Zhu, B. Root functional traits are key determinants of the rhizosphere effect on soil organic matter decomposition across 14 temperate hardwood species. Soil Biol. Biochem. 2020, 151, 108019. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.W.; Kuebbing, S.E.; Karlsen-Ayala, E.; Bradford, M.A. Evidence for the primacy of living root inputs, not root or shoot litter, in forming soil organic carbon. New Phytol. 2018, 221, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Xing, H.; Shi, Z.; Liu, S.; Chen, M.; Xu, G.; Cao, X.; Zhang, M.; Chen, J.; Li, F. Leaf traits divergence and correlations of woody plants among the three plant functional types on the eastern Qinghai-Tibetan Plateau, China. Front. Plant Sci. 2023, 14, 1128227. [Google Scholar] [CrossRef] [PubMed]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-K.; Pan, X.; Liu, G.-F.; Li, W.-B.; Dai, W.-H.; Tang, S.-L.; Zhang, Y.-L.; Xiao, T.; Chen, L.-Y.; Xiong, W.; et al. Novel evidence for within-species leaf economics spectrum at multiple spatial scales. Front. Plant Sci. 2015, 6, 901. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Lavely, E.; Ma, Z. Fine-root and mycorrhizal traits help explain ecosystem processes and responses to global change. New Phytol. 2014, 204, 455–458. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Aguilar-Trigueros, C.A.; Flaig, I.C.; Rillig, M.C. Root trait responses to drought are more heterogeneous than leaf trait responses. Funct. Ecol. 2020, 34, 2224–2235. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root structure–function relationships in 74 species: Evidence of a root economics spectrum related to carbon economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef]
- de la Riva, E.G.; Marañón, T.; Pérez-Ramos, I.M.; Navarro-Fernández, C.M.; Olmo, M.; Villar, R. Root traits across environ-mental gradients in Mediterranean woody communities: Are they aligned along the root economics spectrum? Plant Soil 2018, 424, 35–48. [Google Scholar] [CrossRef]
- Liu, C.; Xiang, W.; Zou, L.; Lei, P.; Zeng, Y.; Ouyang, S.; Deng, X.; Fang, X.; Liu, Z.; Peng, C. Variation in the functional traits of fine roots is linked to phy-logenetics in the common tree species of Chinese subtropical forests. Plant Soil 2019, 436, 347–364. [Google Scholar] [CrossRef]
- Weigelt, A.; Mommer, L.; Andraczek, K.; Iversen, C.M.; Bergmann, J.; Bruelheide, H.; Fan, Y.; Freschet, G.T.; Guerrero-Ramírez, N.R.; Kattge, J.; et al. An integrated framework of plant form and function: The belowground perspective. New Phytol. 2021, 232, 42–59. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Ma, C.; Zhang, Q.; Li, L.; Chen, X.; Zeng, H.; Guo, D. Leading dimensions in absorptive root trait variation across 96 subtropical forest species. New Phytol. 2014, 203, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, J.; Weigelt, A.; van der Plas, F.; Laughlin, D.C.; Kuyper, T.W.; Guerrero-Ramirez, N.; Valverde-Barrantes, O.J.; Bruelheide, H.; Freschet, G.T.; Iversen, C.M.; et al. The fungal collaboration gradient dominates the root economics space in plants. Sci. Adv. 2020, 6, eaba3756. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Bai, W.; Zhang, Y.; Zhang, W. Multi-dimensional patterns of variation in root traits among coexisting herbaceous species in temperate steppes. J. Ecol. 2018, 106, 2320–2331. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenes that reduce the metabolic costs of soil exploration: Opportunities for 21st century agriculture. Plant Cell Environ. 2015, 38, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Zhu, B. Linking root respiration to chemistry and morphology across species. Glob. Chang. Biol. 2021, 27, 190–201. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast-slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.J.W.; van Ruijven, J.; Kuyper, T.W.; Mohren, G.M.J.; Sterck, F.J. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef] [PubMed]
- Erktan, A.; Roumet, C.; Bouchet, D.; Stokes, A.; Pailler, F.; Munoz, F. Two dimensions define the variation of fine root traits across plant communities under the joint influence of ecological succession and annual mowing. J. Ecol. 2018, 106, 2031–2042. [Google Scholar] [CrossRef]
- An, N.; Lu, N.; Fu, B.; Chen, W.; Keyimu, M.; Wang, M. Evidence of differences in covariation among root traits across plant growth forms, mycorrhizal types, and biomes. Front. Plant Sci. 2022, 12, 785589. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Tjoelker, M.G.; Pregitzer, K.S.; Wright, I.J.; Oleksyn, J.; Machado, J. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecol. Lett. 2008, 11, 793–801. [Google Scholar] [CrossRef]
- McCormack, M.L.; Adams, T.S.; Smithwick, E.A.H.; Eissenstat, D.M. Predicting fine root lifespan from plant functional traits in temperate trees. New Phytol. 2012, 195, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Ataka, M.; Han, M.; Han, Y.; Gan, D.; Xu, T.; Guo, Y.; Zhu, B. Root exudation as a major competitive fine-root functional trait of 18 coexisting species in a subtropical forest. New Phytol. 2020, 229, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Iii, F.S.C.; Tryon, P.R. Habitat and leaf habit as determinants of growth, nutrient absorption, and nutrient use by Alaskan taiga forest species. Can. J. For. Res. 1983, 13, 818–826. [Google Scholar] [CrossRef]
- Wang, Q.; Xiao, J.; Ding, J.; Zou, T.; Zhang, Z.; Liu, Q.; Yin, H. Differences in root exudate inputs and rhizosphere effects on soil N transformation between deciduous and evergreen trees. Plant Soil 2021, 458, 277–289. [Google Scholar] [CrossRef]
- Li, J.; Chen, X.; Niklas, K.J.; Sun, J.; Wang, Z.; Zhong, Q.; Hu, D.; Cheng, D. A whole-plant economics spectrum including bark functional traits for 59 subtropical woody plant species. J. Ecol. 2020, 110, 248–261. [Google Scholar] [CrossRef]
- Chen, X.; Le, X.; Niklas, K.J.; Hu, D.; Zhong, Q.; Cheng, D. Divergent leaf nutrient-use strategies of coexistent evergreen and deciduous trees in a subtropical forest. J. Plant Ecol. 2023, 16, rtac093. [Google Scholar] [CrossRef]
- Galmán, A.; Abdala-Roberts, L.; Zhang, S.; Berny-Mier y Teran, J.C.; Rasmann, S.; Moreira, X. A global analysis of ele-vational gradients in leaf herbivory and its underlying drivers: Effects of plant growth form, leaf habit and climatic correlates. J. Ecol. 2018, 106, 413–421. [Google Scholar] [CrossRef]
- Schweiger, R.; Castells, E.; Da Sois, L.; Martínez-Vilalta, J.; Müller, C. highly species-specific foliar metabolomes of diverse woody species and relationships with the leaf economics spectrum. Cells 2021, 10, 644. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, K.W.; van Langevelde, F.; Ward, D.; Bongers, F.; da Silva, D.A.; Prins, H.H.T.; de Bie, S.; Sterck, F.J. Deciduous and evergreen trees differ in juvenile biomass allometries because of differences in allocation to root storage. Ann. Bot. 2013, 112, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef] [PubMed]
- Pagel, M. Inferring the historical patterns of biological evolution. Nature 1999, 401, 877–884. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Mo, W.; Lee, M.-S.; Uchida, M.; Inatomi, M.; Saigusa, N.; Mariko, S.; Koizumi, H. Seasonal and annual variations in soil respiration in a cool-temperate deciduous broad-leaved forest in Japan. Agric. For. Meteorol. 2005, 134, 81–94. [Google Scholar] [CrossRef]
- Tjoelker, M.G.; Craine, J.M.; Wedin, D.; Reich, P.B.; Tilman, D. Linking leaf and root trait syndromes among 39 grassland and savannah species. New Phytol. 2005, 167, 493–508. [Google Scholar] [CrossRef]
- Makita, N.; Kosugi, Y.; Dannoura, M.; Takanashi, S.; Niiyama, K.; Kassim, A.R.; Nik, A.R. Patterns of root respiration rates and morphological traits in 13 tree species in a tropical forest. Tree Physiol. 2012, 32, 303–312. [Google Scholar] [CrossRef]
- Liang, S.; Guo, H.; McCormack, M.L.; Qian, Z.; Huang, K.; Yang, Y.; Xi, M.; Qi, X.; Ou, X.; Liu, Y.; et al. Positioning absorptive root respiration in the root economics space across woody and herbaceous species. J. Ecol. 2023, 111, 2710–2720. [Google Scholar] [CrossRef]
- LeBauer, D.S.; Treseder, K.K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally dis-tributed. Ecology 2008, 89, 371–379. [Google Scholar] [CrossRef]
- Yang, J.; Lan, L.; Jin, Y.; Yu, N.; Wang, D.; Wang, E. Mechanisms underlying legume–rhizobium symbioses. J. Integr. Plant Biol. 2022, 64, 244–267. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, H.; Zhang, W.; Han, B.; Zhou, H.; Lu, X.; Deng, Y.; Liu, K.; Shao, X. Arbuscular Mycorrhizal Fungi Alter the Interaction Effects Between Bacillus and Rhizobium on Root Morphological Traits of Medicago ruthenica L. J. Soil Sci. Plant Nutr. 2023, 23, 2868–2877. [Google Scholar] [CrossRef]
- Whitehead, D.C.; Goulden, K.M.; Hartley, R.D. The distribution of nutrient elements in cell wall and other fractions of the herbage of some grasses and legumes. J. Sci. Food Agric. 1985, 36, 311–318. [Google Scholar] [CrossRef]
- Jia, S.; McLaughlin, N.B.; Gu, J.; Li, X.; Wang, Z. Relationships between root respiration rate and root morphology, chemistry and anatomy in Larix gmelinii and Fraxinus mandshurica. Tree Physiol. 2013, 33, 579–589. [Google Scholar] [CrossRef]
- Ma, Z.; Guo, D.; Xu, X.; Lu, M.; Bardgett, R.D.; Eissenstat, D.M.; McCormack, M.L.; Hedin, L.O. Evolutionary history resolves global organization of root functional traits. Nature 2018, 555, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidi-mensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Jarvi, M.P.; Burton, A.J. Root respiration and biomass responses to experimental soil warming vary with root diameter and soil depth. Plant Soil 2020, 451, 435–446. [Google Scholar] [CrossRef]
- Chen, T.; Lin, C.; Song, T.; Guo, R.; Cai, Y.; Chen, W.; Xiong, D.; Jiang, Q.; Chen, G. Does root respiration of subtropical Chinese fir seedlings acclimate to seasonal temperature variation or experimental soil warming? Agric. For. Meteorol. 2021, 308, 108612. [Google Scholar] [CrossRef]
- Salazar, A.; Rousk, K.; Jónsdóttir, I.S.; Bellenger, J.; Andrésson, S. Faster nitrogen cycling and more fungal and root biomass in cold ecosystems under experimental warming: A meta-analysis. Ecology 2020, 101, e02938. [Google Scholar] [CrossRef]
- Baumert, V.L.; Vasilyeva, N.A.; Vladimirov, A.A.; Meier, I.C.; Kögel-Knabner, I.; Mueller, C.W. Root exudates induce soil macroaggregation facilitated by fungi in subsoil. Front. Environ. Sci. 2018, 6, 140. [Google Scholar] [CrossRef]
- Staszel-Szlachta, K.; Lasota, J.; Szlachta, A.; Błońska, E. The impact of root systems and their exudates in different tree species on soil properties and microorganisms in a temperate forest ecosystem. BMC Plant Biol. 2024, 24, 45. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Wang, J.; Wu, H.; Valverde-Barrantes, O.J.; Wang, R.; Zeng, H.; Kardol, P.; Zhang, H.; Feng, Y. Nonlinearity of root trait relationships and the root economics spectrum. Nat. Commun. 2019, 10, 2203. [Google Scholar] [CrossRef] [PubMed]
- Brundrett, M.C.; Tedersoo, L. Evolutionary history of mycorrhizal symbioses and global host plant diversity. New Phytol. 2018, 220, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
- Steidinger, B.S.; Crowther, T.W.; Liang, J.; van Nuland, M.E.; Werner, G.D.; Reich, P.B.; Nabuurs, G.J.; De-Miguel, S.; Zhou, M.; Picard, N.; et al. Climatic controls of decompo-sition drive the global biogeography of forest-tree symbioses. Nature 2019, 569, 404–408. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Kong, D.; Zhang, Z.; Cai, Q.; Xiao, J.; Liu, Q.; Yin, H. Climate and soil nutrients differentially drive multidimensional fine root traits in ectomycorrhizal-dominated alpine coniferous forests. J. Ecol. 2020, 108, 2544–2556. [Google Scholar] [CrossRef]
- Gao, J.; Zhou, M.; Shao, J.; Zhou, G.; Liu, R.; Zhou, L.; Liu, H.; He, Y.; Chen, Y.; Zhou, X. Fine root trait-function relationships affected by mycorrhizal type and climate. Geoderma 2021, 394, 115011. [Google Scholar] [CrossRef]
Figure 1.
Distribution of the plots within the study area. The image is obtained from Google Earth.

Figure 2.
Root respiration rates across the 16 study species (A) and the distribution of root respiration rates for deciduous and evergreen species (B). Box plots illustrate the interquartile range (25%–75%). * p < 0.05.
Figure 2.
Root respiration rates across the 16 study species (A) and the distribution of root respiration rates for deciduous and evergreen species (B). Box plots illustrate the interquartile range (25%–75%). * p < 0.05.
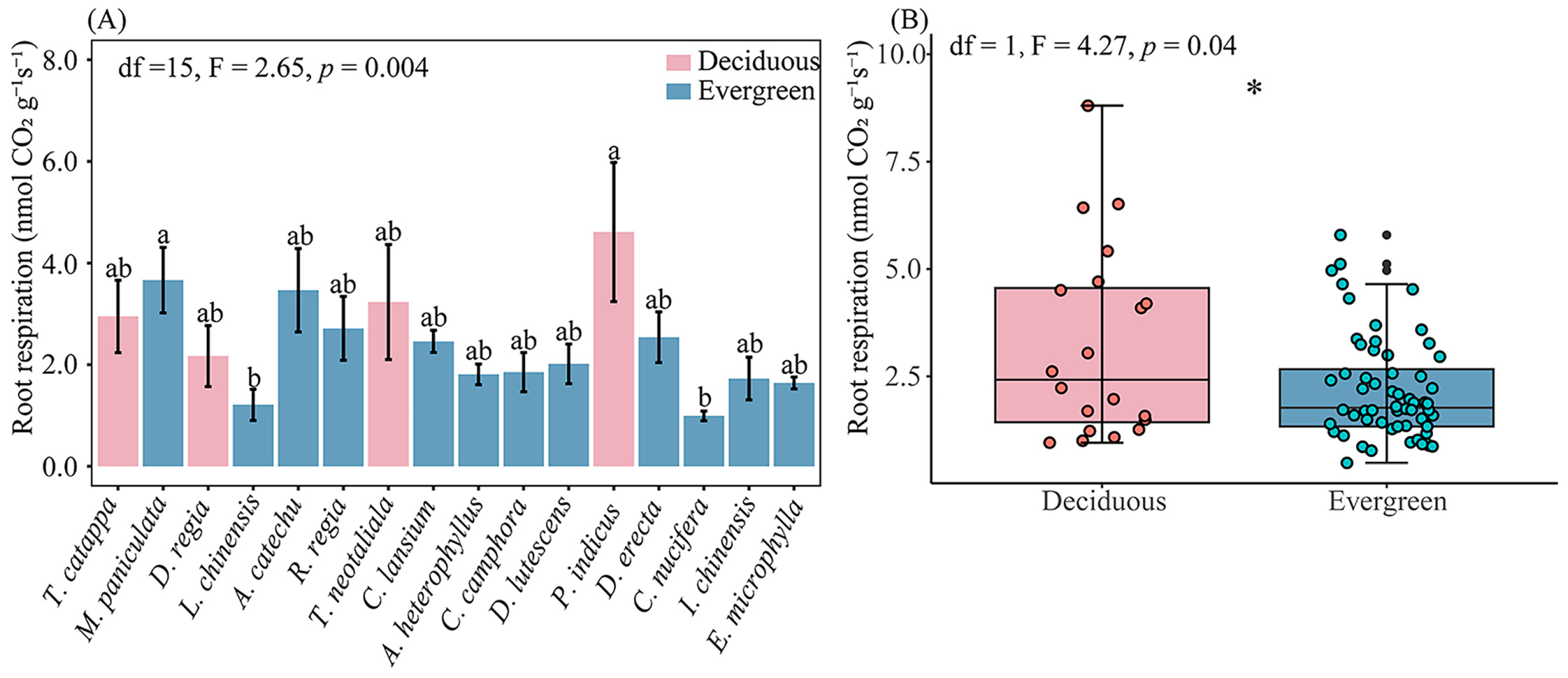
Figure 3.
Relationships between root respiration rate and morphological traits (specific root length [SRL], root diameter [RD], and root tissue density [RTD]) (A–C) and chemical traits (root nitrogen concentration [RNC], root carbon concentration [RCC], and root carbon-to-nitrogen ratio [C/N]) (D–F) for deciduous versus evergreen species.
Figure 3.
Relationships between root respiration rate and morphological traits (specific root length [SRL], root diameter [RD], and root tissue density [RTD]) (A–C) and chemical traits (root nitrogen concentration [RNC], root carbon concentration [RCC], and root carbon-to-nitrogen ratio [C/N]) (D–F) for deciduous versus evergreen species.

Figure 4.
Pairwise correlations among root traits across all species (A) and individual leaf habits: deciduous species (B) and evergreen species (C). C/N, root carbon-to-nitrogen ratio; RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RTD, root tissue density; SRL, specific root length; RR, root respiration rate. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 4.
Pairwise correlations among root traits across all species (A) and individual leaf habits: deciduous species (B) and evergreen species (C). C/N, root carbon-to-nitrogen ratio; RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RTD, root tissue density; SRL, specific root length; RR, root respiration rate. * p < 0.05; ** p < 0.01; *** p < 0.001.

Figure 5.
Principal component analysis for root traits in all species (A), deciduous species (B), and evergreen species (C). RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RR, root respiration rate; RTD, root tissue density; SRL, specific root length.
Figure 5.
Principal component analysis for root traits in all species (A), deciduous species (B), and evergreen species (C). RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RR, root respiration rate; RTD, root tissue density; SRL, specific root length.
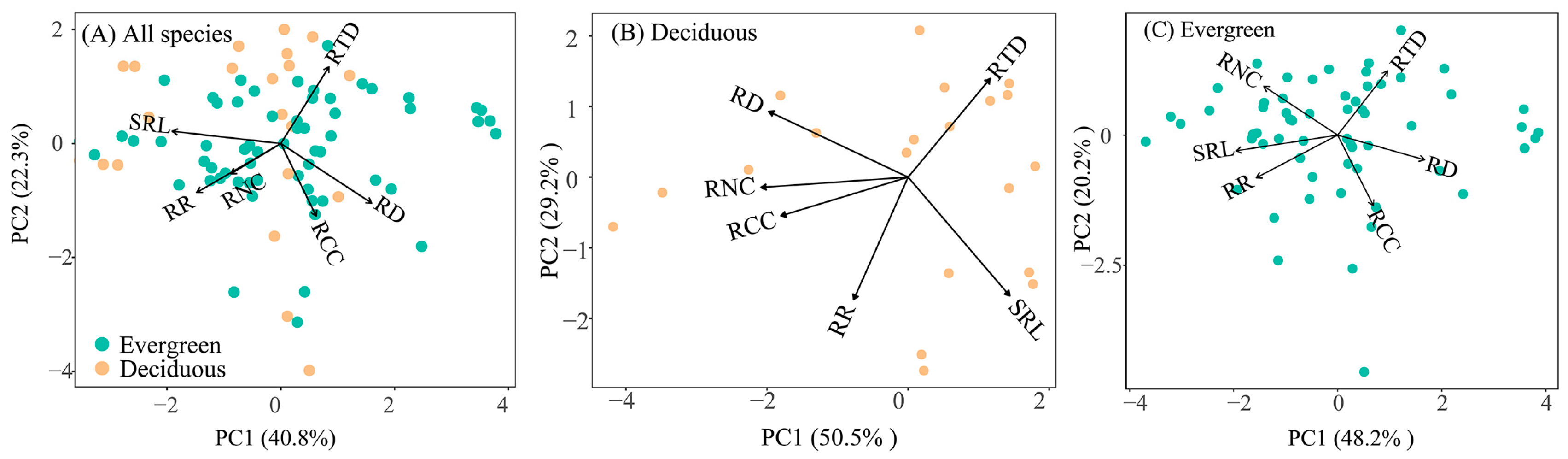
Figure 6.
Relative importance of root morphological and chemical traits for the root respiration rate, as determined using random forest modeling of all species (A), evergreen species (B), and deciduous species (C). %IncMSE represents the % increase in the mean square error. C/N, root carbon-to-nitrogen ratio; RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RTD, root tissue density; SRL, specific root length. **, p < 0.01; *, p < 0.05.
Figure 6.
Relative importance of root morphological and chemical traits for the root respiration rate, as determined using random forest modeling of all species (A), evergreen species (B), and deciduous species (C). %IncMSE represents the % increase in the mean square error. C/N, root carbon-to-nitrogen ratio; RCC, root carbon concentration; RD, root diameter; RNC, root nitrogen concentration; RTD, root tissue density; SRL, specific root length. **, p < 0.01; *, p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Detailed information on the 16 study plant species.
| Species | Family | Leaf Habit | Mean DBH 1/Ground Diameter 2 (cm) | Height (m) |
|---|---|---|---|---|
| Areca catechu L. | Arecaceae | Evergreen | 12.58 ± 0.46 | 8.14 ± 0.31 |
| Artocarpus heterophyllus Lam. | Moraceae | Evergreen | 61.08 ± 6.72 | 12.42 ± 1.28 |
| Cinnamomum camphora (L.) J. Presl | Lauraceae | Evergreen | 67.06 ± 2.56 | 22.78 ± 0.77 |
| Clausena lansium (Lour.) Skeels | Rutaceae | Evergreen | 9.56 ± 1.21 | 4.78 ± 0.17 |
| Cocos nucifera L. | Arecaceae | Evergreen | 34.78 ± 1.13 | 9.22 ± 0.33 |
| Dypsis lutescens (H. Wendl.) | Arecaceae | Evergreen | 7.80 ± 0.58 | 3.66 ± 0.16 |
| Duranta erecta L. | Verbenaceae | Evergreen | 2.16 ± 0.12 | 0.87 ± 0.01 |
| Ehretia microphylla Lam. | Boraginaceae | Evergreen | 2.22 ± 0.27 | 0.62 ± 0.03 |
| Ixora chinensis Lam. | Rubiaceae | Evergreen | 1.78 ± 0.14 | 0.51 ± 0.03 |
| Litchi chinensis Sonn. | Sapindaceae | Evergreen | 39.36 ± 4.56 | 9.36 ± 0.91 |
| Murraya paniculata (L.) Jack | Rutaceae | Evergreen | 2.08 ± 0.31 | 0.83 ± 0.02 |
| Roystonea regia (Kunth) O.F. Cook | Arecaceae | Evergreen | 35.28 ± 2.44 | 15.72 ± 1.00 |
| Delonix regia (Bojer ex Hook) Raffin | Fabaceae | Deciduous | 48.24 ± 5.57 | 14.24 ± 0.28 |
| Pterocarpus indicus Willd. | Fabaceae | Deciduous | 48.98 ± 2.05 | 20.72 ± 0.40 |
| Terminalia catappa L. | Combretaceae | Deciduous | 47.48 ± 4.84 | 24.38 ± 0.69 |
| Terminalia neotaliala Capuron | Combretaceae | Deciduous | 24.16 ± 1.45 | 12.08 ± 0.51 |
1 Mean diameter at breast height; 2 The diameters of the plants are shorter than one meter in height and the stem or trunk measures 30 cm at ground level. Data represent mean ± standard error.
Table 2.
Root functional traits for species belonging to different leaf habits.
| Traits | Deciduous | Evergreen | t | df | p |
|---|---|---|---|---|---|
| SRL (m g−1) | 5.71 ± 0.69 | 5.26 ± 0.32 | 0.52 | 29.29 | 0.600 |
| RD (mm) | 1.05 ± 0.07 | 1.12 ± 0.03 | −1.21 | 27.45 | 0.240 |
| RTD (g cm−3) | 0.25 ± 0.01 | 0.24 ± 0.01 | 1.60 | 49.87 | 0.120 |
| RNC (mg g−1) | 76.10 ± 12.05 | 43.45 ± 2.44 | 2.87 | 26.74 | 0.008 |
| RCC (m g−1) | 429.21 ± 10.1 | 459.70 ± 5.96 | −2.50 | 28.42 | 0.020 |
| C/N | 8.04 ± 0.98 | 19.76 ± 1.13 | −3.60 | 31.00 | 0.001 |
SRL, specific root length; RD, root diameter; RTD, root tissue density; RNC, root nitrogen concentration; RCC, root carbon concentration; C/N, root carbon-to-nitrogen ratio. Data represent mean ± standard error.
Table 3.
Multiple linear regression analysis of the relationship between root traits and root respiration.
Table 3.
Multiple linear regression analysis of the relationship between root traits and root respiration.
| Regression Weights | Standardized Regression Weights | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Beta | Std. Error | Beta | t | p | Vif | R2 | F(p) | ||
| All species | (Intercept) | −0.16 | 0.04 | 0.00 | −4.04 | <0.001 | |||
| SRL (m g−1) | 0.50 | 0.10 | 0.47 | 5.10 | <0.001 | 1.03 | 0.36 | 21.83 (p < 0.001) | |
| RNC (mg g−1) | 0.31 | 0.10 | 0.30 | 3.29 | 0.002 | 1.03 | |||
| Multiple regression analysis model | RR = −0.16 + 0.50SRL + 0.31RNC | ||||||||
| Deciduous | (Intercept) | −0.09 | 0.03 | 0.00 | −2.8 | 0.006 | |||
| SRL (m g−1) | 0.46 | 0.11 | 0.44 | 4.11 | <0.001 | 1.25 | |||
| RTD (mg g−1) | 3.83 | 4.23 | 0.09 | 0.90 | 0.370 | 1.02 | 0.30 | 10.88 (p < 0.001) | |
| RD (mm) | −0.03 | 0.02 | −0.16 | −1.57 | 0.120 | 1.23 | |||
| Multiple regression analysis model | RR= −0.09 + 0.46 SRL + 3.83 RTD − 0.03RD | ||||||||
| Evergreen | (Intercept) | −0.77 | 0.35 | 0.00 | −2.24 | 0.028 | |||
| SRL (m g−1) | 0.60 | 0.10 | 0.57 | 6.86 | < 0.001 | 1.06 | 0.31 | 17.44 (p < 0.001) | |
| RCC (mg g−1) | 0.15 | 0.06 | 0.20 | 1.62 | 0.11 | 1.06 | |||
| Multiple regression analysis model | RR = −0.77 + 0.60SRL + 0.15RCC | ||||||||
RR, root respiration rate; SRL, specific root length; RNC, root nitrogen concentration; RD, root diameter; RCC, root carbon concentration. Vif, Variance Inflation Factor. Regression Weights, unstandardized regression coefficient; Standardized Regression Weights, standardized regression coefficient.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Deng, D.; Sun, Y.; Yang, M. Root Respiration–Trait Relationships Are Influenced by Leaf Habit in Tropical Plants. Forests 2024, 15, 806. https://doi.org/10.3390/f15050806
AMA Style
Deng D, Sun Y, Yang M. Root Respiration–Trait Relationships Are Influenced by Leaf Habit in Tropical Plants. Forests. 2024; 15(5):806. https://doi.org/10.3390/f15050806
Chicago/Turabian StyleDeng, Danting, Yanfei Sun, and Meiqiu Yang. 2024. "Root Respiration–Trait Relationships Are Influenced by Leaf Habit in Tropical Plants" Forests 15, no. 5: 806. https://doi.org/10.3390/f15050806
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.