
Advances in Endangered Plant Research: Ammopiptanthus’s Responses to Biotic and Abiotic Stressors
1
Guangxi Key Laboratory of Special Non-Wood Forests Cultivation and Utilization, Guangxi Xylophyta Spices Research Center of Engineering Technology, Guangxi Forestry Research Institute, Nanning 530002, China
2
National Engineering Research Center of Tree Breeding and Ecological Restoration, College of Biological Sciences and Technology, Beijing Forestry University, Beijing 100083, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Forests 2024, 15(5), 890; https://doi.org/10.3390/f15050890
Submission received: 24 April 2024
/
Revised: 13 May 2024
/
Accepted: 16 May 2024
/
Published: 20 May 2024
(This article belongs to the Special Issue Abiotic Stress in Tree Species)
Abstract
:Ammopiptanthus, a unique evergreen broadleaf shrub endemic to the desert regions of Northwest China, exhibits a remarkable capacity for aeolian erosion control and dune stabilization. Attributed to its robust tolerance in terms of xerotolerance, halophytic adaptations, extreme thermotolerance, resistance to biotic and abiotic degradation and its defensive strategies against herbivory, Ammopiptanthus has emerged as an exemplary model organism for the study of plant resilience to diverse environmental stressors. Current research on Ammopiptanthus is scattered, lacking a systematic review, which poses a disadvantage for subsequent in-depth studies and the effective conservation of this endangered resource. In recent years, natural Ammopiptanthus communities have been severely disrupted, and the species’ natural range is rapidly shrinking. Here, this review summarizes the signaling pathways in the Ammopiptanthus response to biotic stress (especially the early signaling events), as well as the research advances in the resistance interactions between biotic and abiotic stresses. Then, the synergistic effects of multiple environmental pressures on Ammopiptanthus could be established, which may provide guidance for further studies on the resistance mechanism of Ammopiptanthus and be beneficial to its natural community protection and reconstruction.
1. Introduction
Ammopiptanthus, belonging to the leguminous genus, is an ancient relic species from the Tertiary period of the ancient Mediterranean region in the Central Asian arid plant flora in the arid Northwest China [1,2]. Simultaneously, it stands as the sole evergreen broadleaf shrub in the northwestern deserts of China. Ammopiptanthus comprises only two species, namely the Ammopiptanthus nanus (M.Pop.) Cheng f. and the Ammopiptanthus mongolicus (Maxim) Cheng f [1,3]. Both of these species are classified as third-level endangered species and second-level endangered species at the national level in China, respectively [2]. Their habitats are characterized by extreme aridity, with annual precipitation levels below 200 mm, high annual evaporation rates reaching up to 2000 mm, and extreme temperature variations [4]. These regions can experience scorching high temperatures of up to 40 °C and plummeting low temperatures as low as −30 °C [5]. The soil is often infertile, accompanied by salinity and alkalinity stress, and they are also susceptible to severe pest infestations [4]. Over the past decade, many researchers have devoted themselves to in-depth investigations of the physiological and molecular mechanisms by which Ammopiptanthus species respond to both biotic and abiotic stressors [6,7,8,9,10,11,12]. The primary objective of this article is to provide a comprehensive review of the defensive responses and stress resistance mechanisms in Ammopiptanthus, shedding light on the physiological and molecular patterns employed by Ammopiptanthus when facing environmental stressors, as well as their interplay in resistance mechanisms. This in-depth discussion holds the promise of not only safeguarding this unique plant species but also promoting its sustainable development and utilization. Furthermore, it contributes to the maintenance of ecological balance in desert regions.
2. Defense Response in Ammopiptanthus
Defense response refers to the adaptive process in which plants exhibit a series of specific internal metabolic changes and external structural alterations to enhance their survival capabilities in the face of invasion by insects and pathogens. This process encompasses various biological processes, including the recognition of external stimuli, the transduction of stimulus signals, the regulation of defense gene expression, the synthesis and accumulation of bioactive substances, and the realization of defensive effects [13]. In the perpetual contest between flora and fauna, devoid of the smoke of war, the strategies by which plants harness their own resources to mount a defense against pests have remained a focal point of scientific inquiry [14].
2.1. Insect Populations Affecting on Ammopiptanthus
In the natural environment, pests represent a pivotal threat to the conservation status of Ammopiptanthus, contributing significantly to its vulnerability and endangerment [4]. Owing to the adverse environmental conditions, the diversity of plant species that serve as fodder for indigenous herbivorous insects is comparatively limited. The phytophage-induced pressures on the population succession of Ammopiptanthus are escalating, posing significant ecological challenges [15,16]. According to the findings of Liu et al. [17], their research revealed a staggering number of 36 insect species that pose a threat to Ammopiptanthus, with 32 of them affecting the above-ground portions of the plant [17]. This group of pests primarily belongs to the order Coleoptera, including insects from families such as Tenebrionidae, Curculionidae, Chrysomelidae, and other related families. Additionally, insects from the order Hemiptera (such as aphids), Lepidoptera (toxic moths), Heteroptera (including shield bugs), Dictyoptera (cockroaches), Orthoptera (crickets and grasshoppers), Dermaptera (earwigs), and Thysanoptera (thrips) also pose threats to Ammopiptanthus (Table 1). Additionally, a seed-damaging pest, the legume pod borer (Etiella zinckenella), can cause damage rates of up to 80% in artificially cultivated areas and 60% in natural growth areas of Ammopiptanthus in Ningxia, with a spatial distribution that exhibits an aggregated pattern [18]. During the young leaf, flowering, and fruiting stages, Ammopiptanthus is most susceptible to insect infestations [17]. Faced with such severe pest infestations, effective pest control to maintain stable pest populations and prevent outbreaks is a key focus for researchers in recent years (in Figure 1). Pest control methods can be categorized into physical control, chemical control, and biological control based on the approach adopted [19,20]. Physical control primarily involves manual removal or destruction of pest cocoons. Chemical control heavily relies on pesticide application, and both of these methods may have limitations or significant environmental side effects. In contrast, biological control holds the promise of being an environmentally friendly approach to pest management. Researchers employ various methods to implement biological pest control. These methods include the use of parasitoid predators to control pest populations, utilizing pheromones to attract and capture female or male insects, and introducing O. ericae in their larval stage carrying nucleopolyhedrovirus (CyunNPV) into pest-infested areas to control pest densities (in Figure 1).
2.2. Early Biotic Stress Signaling in Ammopiptanthus
The viability of plants is contingent upon their adeptness in perceiving and responding to extracellular signals with celerity and efficacy, thereby facilitating the genesis and sustenance of resilient and efficacious defensive strategies [21]. Upon herbivory, plants detect insect-mediated mechanical damage and oral secretion (OS)-derived elicitors, triggering foliar cell-based signaling pathways [22,23]. This detection precipitates a series of early signaling events that are integral to the activation of the plant’s defense mechanisms, thereby enabling a rapid and orchestrated response to the biotic stress imposed by the feeding insects [24]. Initially, the herbivore feeding site activates pattern recognition receptors (PRRs) that discern molecules within the OSs of insects [25]. Subsequently, there is an immediate alteration in the transmembrane potential (Vm), leading to a rapid burst of ROS (reactive oxygen species) and a [Ca2+]cyt [26,27]. This constitutes the plant’s early response to herbivory. Within minutes, these early signaling events trigger the activation or suppression of transcription factors within the plant cells, thereby modulating hormonal signaling and inducing the accumulation of defense-related compounds through the upregulation of defense genes [28,29,30]. Research involving Gao et al.’s [31] subjects has indicated that O. ericae OSs trigger the transmembrane flux of Ca2+ in Ammopiptanthus leaves. It is noteworthy that calcium channel blockers (GdCl3) do not fully suppress the flux of Ca2+ within this pathway. This observation suggests that in addition to the Ca2+ channels located on the lipid membrane, there are alternative pathways that induce the influx of Ca2+. Furthermore, the treatment by OSs has been found to result accumulation of H2O2, which is facilitated by the enzymatic activity of NADPH oxidase [31]. Treatment with the extracellular Ca2+ chelator EGTA completely inhibited the OS-induced H2O2 accumulation in leaf cells, indicating that the accumulation of H2O2 induced by OSs relies on the transmembrane influx of extracellular Ca2+ [31]. This investigation represents one of the pioneering studies on the early signaling responses of Ammopiptanthus to the herbivory elicited by insects. Different stimuli or different insect feeding behaviors can induce distinct early signaling events in plants [24,32]. The researchers posited that the herbivorous activities of insects, on the one hand, result in the ingress of OSs into cells, which subsequently serve as chemical stimuli to the compromised regions of the plant’s cellular structure. Concurrently, this feeding triggers the early signaling events within the plant, initiated by the mechanical trauma inflicted by the insects [33,34,35]. Gao et al. [36] reported that after mechanical stimulation, Ammopiptanthus leaf cells undergo a rapid and pronounced influx of Ca2+, reaching a peak at approximately 5 min post-perturbation (circa 160 pmol·cm−2·s−1), after which the influx exhibits a declining trend [36]. Upon the application of LaCl3 to inhibit the Ca2+ channels, the mechanostimuli-induced Ca2+ influx is completely abolished. For mechanical stimulation, the transmembrane influx of Ca2+ is primarily derived from the extracellular environment, which primarily denotes the region where calcium ions are retained outside the plasma membrane or the test solution’s Ca2+ content. It enters the cell through voltage-gated Ca2+ channels on the membrane and voltage-insensitive stress-activated Ca2+ channels [36]. The early Ca2+ signal response patterns elicited by mechanical stimulation are notably distinct from those induced by insect OSs, with the source of intracellular Ca2+ accumulation differing despite both stimuli leading to Ca2+ transmembrane influx. Therefore, analyzing the Ca2+ signal generation mode is crucial to elucidating the insect-resistant signaling pathway of Ammopiptanthus. Researchers suggest that this might be due to certain substances carried by insect OSs that can bind to the plant cell membrane, inducing the formation of calcium channels that allow Ca2+ to pass through [35]. Liu found that, in addition to calcium channels, Ca2+-ATPase and Ca2+/H+ antiporters also participate in the transmembrane transport of Ca2+ induced by insect OSs (unpublished data). H+ transmembrane transport directly influences cytoplasmic pH, and pH-sensitive proteins change their activity in response to variations in cytoplasmic pH to adapt to environmental changes [37]. In Ammopiptanthus, salt stress induces an enhancement in H+-ATPase activity, facilitating the extrusion of H+ ions across the membrane [4]. This provides the proton motive force for the Na+/H+ antiporters to pump excess Na+ out of the cell while simultaneously allowing the H+ transported out by the H+-ATPase to re-enter the cell, thereby maintaining intracellular pH balance [4,38,39]. Unlike the H+ signal induced by salt stress, in the case of mechanical damage, Ammopiptanthus cells exhibit H+ transmembrane influx, and this H+ influx depends on the transmembrane influx of Ca2+ and the accumulation of intracellular H2O2 [36]. It would be intriguing to explore the potential involvement of H+ fluxes and H+-ATPases in the modulation of the possible contributions to Ammopiptanthus’s defensive responses against herbivory.
2.3. Metabolic Changes in Ammopiptanthus
Faced with severe biotic stress from pest infestations, plants have evolved over a long ecological timeframe to develop multiple effective biological and physiological mechanisms that enhance its survival capabilities and mitigate damage caused by pests [40,41]. These defense mechanisms encompass both direct responses, such as the synthesis of toxic secondary metabolites for pest deterrence, and indirect strategies, like the emission of specific volatile organic compounds (VOCs) such as terpenes to attract natural predators of pests [42]. Additionally, these volatiles can serve as an early warning signal to surrounding plants, inducing collective defense responses [43,44]. Sun et al. [40] performed an in-depth metabolomic analysis of A. nanus following its interaction with the herbivorous O. ericae. Their findings revealed significant alterations in the plant’s metabolome, which includes primary metabolites critical for growth and development and secondary metabolites related to defense mechanisms in response to the herbivorous challenge [40]. Jasmonic acid is a pivotal secondary metabolite that plays a crucial role in the plant’s defense mechanism against insect herbivory [33]. The investigation conducted by Chen et al. [4] elucidates that subsequent to herbivorous feeding, significant accumulation of JA occurs in both the foliar and root tissues of Ammopiptanthus [4].
According to the early literature, the branches and leaves of Ammopiptanthus have been used in traditional medicine. They have a slightly bitter and pungent taste and are known to possess some level of toxicity. In some cases, animals have exhibited signs of intoxication after consuming them, and excessive consumption can even lead to death [45]. This might be attributed to the presence of toxic compounds such as alkaloids, lignans, flavonoids, and other substances in Ammopiptanthus leaves and young branches [41,45]. Gao revealed that a water decoction of A. mongolicus at a concentration of 1 g∙L−1 is effective in achieving toxicity against pine wood nematodes in pine seedlings. In contrast, a higher concentration of 6 g∙L−1 of camelthorn, a dominant desert plant, extract is required to exhibit a more substantial insecticidal effect [46]. When treated with a methanol extract from A. mongolicus, it was found that the extract significantly inhibited the growth of diamondback moth larvae at a concentration of 0.2 g∙mL−1 [47]. These early studies on the metabolic products of Ammopiptanthus have confirmed that this plant indeed has the capability to synthesize compounds related to insect resistance. Plants exhibiting enhanced responses to repeated herbivory by conspecific insects represent a crucial adaptive strategy to cope with the cyclical outbreaks of pests in natural ecosystems [48,49]. Building on this, Sun et al. [50] conducted research and discovered that the Ammopiptanthus mongolicus exhibits a low plasticity in its metabolic response to the feeding of O. ericae. However, its biochemical defense mechanisms can significantly modulate and suppress the pest’s subsequent feeding behavior on the plant. Furthermore, this memory-like metabolic alteration and the induced signal amplification effect are predominantly pronounced in the directly affected leaves, while a relatively weaker effect is observed in the leaves of adjacent branches, indicating a certain degree of spatial heterogeneity within the plant’s defense response [50]. Sun et al. [41] also conducted a systematic study of the metabolic changes in Ammopiptanthus exposed to Herbivore-Induced Plant Volatiles (HIPVs). When exposed to HIPVs, the metabolic pathways of Ammopiptanthus in leaves undergo changes that are similar to those induced by insect feeding, for example, the increase in metabolites such as glycolate and ethanol in the leaves. However, the effects on the metabolic pathway of roots are different; a reduction in the levels of various amino acids (such as valine, alanine, aspartate, threonine) and other low-molecular-weight compounds within the foliar tissues, in conjunction with a corresponding increase in these metabolites within the root system, suggests that plants, in response to HIPVs, may reconfigure their metabolic pathways to enhance their defensive preparedness against subsequent insect assaults [41]. In the preceding chapter, we examined the divergent Ca2+ signaling pathways triggered by mechanical injury versus those induced by insect herbivory. Compared to continuous mechanical damage, the changes in primary and secondary metabolite content in treated leaves and systemic leaves of Ammopiptanthus induced by O. ericae feeding were more significant. For instance, in leaves subjected to insect feeding, the contents of compounds such as hydroxybenzenes, proline, acetone, thymol, aspartic acid, and some unidentified sugars and organic acids were notably higher compared to the control. In contrast, the contents of compounds like sucrose, pinene, ethanol, lactic acid, and cucurbitacins in mechanically damaged leaves were significantly lower than in the control [40]. It was also observed that both insect feeding and mechanical damage, when followed by application of OSs after 24 h, resulted in an increase in the alkaloid content of Ammopiptanthus. This increase was noted for alkaloids such as nicotine, cucurbitacins, and piperidine (unpublished data). Phenolic compounds are a major group of chemicals in plants that have toxic effects on herbivorous insects [51]. After insect feeding, an increase in the content of hydroxybenzenes and related compounds was identified in Ammopiptanthus, indicating that insect feeding can promote the accumulation of phenolic compounds in the plants (unpublished data). This is one way in which Ammopiptanthus directly defends itself against herbivores. The accumulation of defensive compounds like alkaloids and phenolic compounds induced by insect feeding may be closely related to the JA pathway [52,53]. As previously mentioned, upon herbivory stress exerted by O. ericae on the leaves of the A. nanus, there is a significant upregulation of endogenous JA levels within the leaf tissue. This biochemical response reflects the plant’s adaptive mechanisms and the activation of its defense strategies in response to adverse conditions [4].
In summary, we have summarized the defense signaling pathway of Ammopiptanthus against insect herbivory. In brief, when Ammopiptanthus is subjected to insect feeding, the elicitors carried in the insect OS are recognized by the corresponding receptors on the damaged cell membrane, resulting in the transmembrane influx of extracellular Ca2+ through plasma membrane Ca2+ channels. Simultaneously, intracellular calcium stores release Ca2+ into the cytoplasm, causing a rapid increase in cytosolic Ca2+ ([Ca2+]cyt) concentration. The increase in [Ca2+]cyt might lead to the opening of inward-rectifier K+ channels on the plasma membrane, causing membrane depolarization, while the insect herbivory-induced efflux of H+ may balance the membrane depolarization caused by the influx of K+. The elevation in [Ca2+]cyt activates plasma membrane NADPH oxidase, which catalyzes the production of superoxide anions (O2·−), leading to the formation of H2O2 under the action of superoxide dismutase (SOD). H2O2 permeates the membrane and enters the cytoplasm, resulting in a burst of cytoplasmic reactive ROS. The accumulation of intracellular ROS may further promote the accumulation of [Ca2+]cyt. The increased Ca2+ binds to downstream calcium sensors, including calmodulin (CaM), calmodulin-like proteins (CMLs), calcineurin B-like proteins (CBLs), calcium-dependent protein kinases (CPKs), and calcium/calmodulin-dependent protein kinases (CDPKs) [54]. These calcium-binding proteins facilitate the phosphorylation or activation of associated proteins, thereby triggering the transcription of specific stress-responsive genes, leading to the biosynthesis of secondary metabolites such as alkaloids and phenolic compounds [55,56]. During this process, the excessive accumulation of [Ca2+]cyt is transported out of the cell via plasma membrane Ca2+-ATPase and Ca2+/H+ antiporters to prevent ion toxicity (in Figure 2).
3. Abiotic Resistance Mechanisms in Ammopiptanthus
Ammopiptanthus, a plant species native to arid regions, exhibits a remarkable resilience to abiotic stressors such as water deficit, soil salinity, and temperature extremes, which typically hinder growth and development in many plant species. This robustness is attributed to the plant’s sophisticated mechanisms of adaptation that enable it to flourish in environments characterized by water scarcity, high salinity, and fluctuating temperatures. The adaptive strategies employed by Ammopiptanthus can be broadly categorized into three interconnected facets: physiological adjustments, modulation of protective enzyme systems, and metabolic level alterations [15,57,58,59].
3.1. Physiological Adjustments
These encompass a range structural and physiological adaptations. In its long-standing struggle against adversities, the Ammopiptanthus has evolved a unique xerophytic structure [60,61]. The plant exhibits a well-developed root system with an abundance of lateral roots. The upper and lower epidermis of the leaves bear a dense layer of epidermal hairs [60]. The stomata are sunken, and the spongy tissue of the mesophyll is degenerated, while the palisade tissue is well developed [61,62]. The cells have thick walls, a dense cytoplasm, and contain abundant intracellular inclusions. The leaves are relatively large with a thick cuticle on the adaxial surface [62]. The extensive root system of Ammopiptanthus not only serves as a vital condition for its survival in desert regions but also forms the essential foundation for its role in preventing wind erosion and mitigating the damage caused by sandstorms [63]. This robust root architecture provides the necessary anchorage in the shifting sands and contributes to soil stabilization, thereby protecting the plant from the harsh and erosive effects of desert winds. When subjected to low temperatures, Ammopiptanthus growth is slowed, resulting in shorter plants and thicker leaves. As temperatures rise, the stems elongate and the leaf area and plant biomass gradually increase [64]. When confronted with extreme temperature fluctuations, Ammopiptanthus can promptly modulate its physiological state to maximize carbon dioxide assimilation efficiency during the spring and summer seasons, thereby sustaining high photosynthetic capacity. Under winter conditions, the plant increases the level of dark-sustained thermal energy dissipation to dissipate excess excitation energy within the photosynthetic system, thereby mitigating photo-oxidative damage to leaf tissues due to cold stress [64]. The insertion of AmCBF1 from A. mongolicus into Upland cotton R15 enhanced the drought and cold stress tolerance of transgenic cotton and improved photosynthesis [65] Additionally, Fei and colleagues [66] have elucidated certain aspects of the cold tolerance physiology in Ammopiptanthus. Utilizing a suite of biochemical techniques, they have successfully isolated and purified a variety of highly active antifreeze proteins (AFPs) from the leaves of Ammopiptanthus, which are known to enhance the plant’s frost resistance [66]. AFPs constitute a category of bioactive macromolecules that are instrumental in the cold acclimation and frost tolerance of flora [67,68]. By reducing the freezing point and inhibiting the formation of extracellular ice, AFPs mitigate cellular damage and desiccation, which are commonly associated with low-temperature stress [67]. These proteins are particularly valuable as they augment the plant’s ability to endure sub-zero temperatures without the deleterious intracellular ice formation that can lead to cellular demise [67].
3.2. Modulation of Protective Enzyme Systems
In addition to its physiological structures, several defense mechanisms are crucial for the survival of Ammopiptanthus. Under abiotic stress, the upregulation of protective enzymes such as superoxide dismutase (SOD), catalase (CAT), and peroxidases (POD) occurs to mitigate the damaging effects of reactive oxygen species (ROS) [69]. These enzymes function in concert to detoxify ROS and maintain cellular redox homeostasis, thereby preserving cellular integrity and function [70,71,72]. In the study conducted by Shi et al., the expression patterns of ROS scavenging enzyme genes under cold, drought, salt, and high-temperature stress conditions were investigated [73]. It was discovered that the expression of superoxide dismutase (AmSOD) and glutathione peroxidase (AmGPX) were upregulated across all four types of stress conditions. Additionally, the research revealed that catalase (AmCAT) exhibited a significantly higher expression level when dealing with excessive H2O2 compared to glutathione peroxidase (AmAPX), thioredoxin (AmPrx), and AmGPX. This suggests that in the antioxidant defense system of Artemisia argyi, AmCAT may play a more critical role in decomposing excess H2O2 to prevent oxidative damage. Furthermore, they noted that AmSOD, AmGPX, and AmAPX showed a more pronounced response to high-temperature stress [73]. Upon the overexpression of the dehydration-responsive element-binding protein 5.1 (AnDREB5.1) in Nicotiana tabacum, the resulting transgenic lines demonstrated a robust enhancement in their antioxidative defense systems. The elevated expression of AnDREB5.1 has been observed to modulate the transcription of additional genes implicated in stress responses [74]. The overexpression of the AmNAC24 gene enhances the activity of antioxidant enzymes, thereby reducing the excessive accumulation of ROS [75]. Research by Song et al. revealed that the A. mongolicus has a stronger cold adaptation compared to the A. nanus. Low-temperature stress reduces the photosynthetic efficiency in both species, but it also increases the activities of antioxidant enzymes like SOD, POD, and APX. To a certain extent, this serves as a safeguard for the photosynthetic membrane system against harm caused by ROS, ensuring that both Ammopiptanthus can sustain photosynthesis even in low-temperature conditions [76]. Han et al. [77] report that Ammopiptanthus’s type 2C protein phosphatase gene plays a negative role in ABA-induced stomatal closure, resulting in transgenic plants having higher transpiration rates than the wild type when induced with PEG6000. However, this process also enhances the plant’s absorption of inorganic salt ions, which is considered a survival strategy for Ammopiptanthus under prolonged drought stress [77]. The increase in soluble small molecules not only helps maintain cell osmotic pressure but also lowers the cytoplasmic freezing point, preventing damage to the membrane system caused by the formation of ice crystals at low temperature. Inositol galactoside synthase is involved in catalyzing the biosynthesis of inositol galactoside. Inositol galactose and sucrose synthesize raffinose through the action of raffinose synthase and raffinose series oligosaccharides, including raffinose sugar. RFO is the second largest sugar in plant tissues [78]. As a nonstructural carbohydrate, it plays an important role in plant growth and development and resistance to abiotic stress [78,79]. Low temperatures, osmotic stress, and NaCl induce the upregulation of the Ammopiptanthus inositol galactosyltransferase gene (AnGolS1) [9]. PLD acts as a signaling amplifier by producing a lipid second messenger, PA, which is involved in physiological responses during development and stress processes. Yu et al. cloned the PLD gene from Ammopiptanthus, and through functional verification in Arabidopsis, they found that AnPLD is primarily located in the cytoplasm. Its expression is upregulated in response to low temperature, drought, and salt stress. Compared to wild-type plants, the transgenic plants exhibited stronger drought tolerance [80]. AnBADH is the betaine aldehyde dehydrogenase gene in Ammopiptanthus, responsible for catalyzing the biosynthesis of betaine. Research has shown that this gene is also induced in response to low temperature, drought, and salt stress [6]. These evolved defense mechanisms and protective enzyme systems contribute to the survival and propagation of Ammopiptanthus under drought conditions [6,9,69,73,74,75,76,77,78,79,80]. In-depth studies of these functional genes not only help protect Ammopiptanthus and ecosystem recovery but also provide the opportunity to explore resistance traits in other plant species and even develop genetically modified plants to cope with environmental stresses such as drought [6,9,69,73,74,75,76,77,78,79,80].
3.3. Metabolic Level Alterations
The plant’s metabolic pathways are also subject to reprogramming in response to abiotic stress. This includes the biosynthesis of osmoprotectants (e.g., proline, glycine, betaine) that stabilize cellular structures and the activation of stress-responsive genes that encode for heat shock proteins (HSPs) and other molecular chaperones that aid in protein refolding and prevent aggregation. Through these multifaceted adaptations, Ammopiptanthus has evolved to not only withstand but also to proliferate in environments that pose significant challenges to the growth and survival of less resilient species. Overexpression of AmDHN132 (Dehydrins (DHNs), also known as group 2 late embryogenesis abundant (LEA) proteins), AmDHN154, and AmDHN200 in transgenic Arabidopsis has been found to enhance salt, osmotic, and cold tolerance [81]. Moreover, Fu’s research demonstrated that Ammopiptanthus subjected to 20% PEG6000 treatment exhibited 45 differentially expressed proteins in its leaves compared with the control group. These proteins are closely associated with processes such as photosynthesis, ROS scavenging, protein synthesis, processing and degradation, material transport, defense responses, RNA processing, and amino acid metabolism. For instance, there is an upregulation in the expression levels of the chloroplast function protein YCF73, which is involved in the assembly and structural stability of Photosystem I (PSI). Concurrently, alterations occur in the abundance of peroxisomal membrane proteins belonging to the Mpv17/PMP22 family, which are closely associated with the scavenging of reactive oxygen species (ROS). A significant upregulation of RNA-binding proteins may be implicated in post-transcriptional gene expression regulatory mechanisms [82]. In our previous research, we have successfully identified a protein from A. nanus belonging to the WRKY gene family. Our studies indicate that by increasing the expression of AnWRKY29 in transgenic plants, there is a significant increase in the accumulation of inorganic ions and organic solutes within the plant, thereby enhancing the plant’s tolerance to drought [83]. Our findings underscore the importance of AnWRKY29 and AnHSP90 influencing trehalose content in A. nanus for osmotic stress adaptation, vital for its survival and ecological robustness [84]. During the integrated analysis of the physiological, biochemical, and transcriptomic characteristics of A. nanus leaves under osmotic stress conditions, Sumbur identified a series of adaptive changes in A. nanus [85]. Specifically, a reduction in stomatal conductance was observed, indicating a physiological response of stomatal closure to minimize water vapor loss [61,62]; the synthesis pathway of cuticular waxes in the epidermis was enhanced, as evidenced by changes in the total wax content and its components, representing a chemical defense mechanism of the plant against osmotic stress [61,62]; and, concurrently, transcriptomic analysis revealed adjustments in the gene expression patterns related to the synthesis of very long-chain fatty acids, alkane metabolism, and wax transport [61,62]. Further research found that under osmotic stress, A. nanus effectively regulated the accumulation of cuticular waxes by upregulating the expression of key genes such as KCS (3-ketoacyl-CoA synthase) and KCR (3-ketoacyl-CoA reductase), involved in the synthesis of very-long-chain fatty acids and members of the ABCG family of transport proteins that regulate wax transport [85]. This molecular-level regulatory mechanism enhances the plant’s tolerance to abiotic stress, especially in water-limited desert environments [85]. Yang et al. [64] concluded that the cellular homeostasis was maintained by adjusting its metabolic pathway under high-temperature stress. In the face of high-temperature stress, the plant biosynthesizes a variety of antioxidants, including anthocyanins, flavonoids, isoflavonoids, etc., to enhance its antioxidant defense system, thereby protecting cells from oxidative damage. In addition, it regulates its photosynthetic-related metabolic pathways, such as porphyrin and chlorophyll biosynthesis pathways and carotenoids metabolism in order to adapt to high temperatures. These adjustments help protect the photosynthetic organs from light damage and maintain the efficient use of light energy. At the same time, the metabolic pathways of plant hormones such as abscisic acid (ABA) and other signaling molecules also change under high temperature stress to regulate plant growth and development and enhance their adaptability to abiotic stress [64].
In-depth research into these mechanisms not only aids in the protection and restoration of Ammopiptanthus and its ecosystem but also facilitates the development of transgenic plants resistant to abiotic stress, adapting to adverse environmental conditions. Furthermore, these studies contribute to our understanding of the adaptation mechanisms of Ammopiptanthus to extreme temperatures, including responses to both low and high temperatures and the expression of relevant genes, providing valuable insights for improving agriculture and ecological conservation.
4. Synergistic Effects of Multiple Environmental Pressures
Each plant must compete for its survival, especially under stressful conditions, when these resources are limited. How to integrate resources to deal with multiple environmental pressures is recognized as an important international research hot spot [86]. Recent studies have indicated that Ammopiptanthus exhibits interactive resistance when responding to external biotic and abiotic stressors [4]. Investigating the interactions and resistance mechanisms of plants when confronted with abiotic (such as drought, salinity, extreme temperatures, etc.) and biotic (such as pathogens, pests, etc.) stress factors is of paramount importance for a profound understanding of plant adaptability and survival strategies [87,88]. Researchers note that the dense trichomes on the leaves and stems of plants not only help them adapt to arid environments but also contribute to their defense against extreme temperatures and ultraviolet radiation. Additionally, these trichomes potentially serve as physical barriers against pests and pathogens [89,90]. Specifically, our current study has shown that the formation of trichomes is significantly increased in A. nanus plants subjected to salt stress (Unpublished data). The R2R3 MYB-dependent auxin signaling pathway plays an important role in the regulation of the formation and development of tomato trichomes. It can increase the density of type II and type VI surface trichomes and enhance the resistance of tomato plants to spider mites [89]. This suggests that the presence of a physical barrier effect to deter the invasion of pests, as well as a mechanism involving the secretion of insecticidal chemical compounds, may exist in A. nanus.
Wang found that the trypsin inhibitor gene AmTI (trypsin inhibitors, TIs), cloned from A. mongolicus, can be induced and upregulated by dehydration, low-temperature, and salt stress [91]. Notably, the expression of trypsin inhibitor genes was initiated within one hour of exposure to these stressors. The most significant upregulation of AmTI expression was observed at the three-hour mark under low-temperature and dehydration stress, and this heightened expression remained stable even after 24 h of exposure [91]. Trypsin inhibitors are archetypal anti-herbivory peptides that can be induced and upregulated by abiotic stress [92,93]. They exert their insecticidal action by inhibiting the activity of proteolytic enzymes, specifically trypsin, within the digestive tract of insects and other animals. This inhibition interferes with the normal proteolytic processes essential for digestion and absorption of proteins, thereby impeding the growth and development of the organisms [94]. This suggests that the expression of AmTI, induced by abiotic stress, is likely to confer insect resistance in Ammopiptanthus. It indicates a potential for cross-resistance mechanisms to operate when the plant is responding to both abiotic and biotic stressors. Dehydrins are important stress-response proteins involved in physiological processes induced by various stress factors in plants, including water stress, salt stress, extreme temperatures, and pathogen infections [95]. Chitinase is a glycoside hydrolase enzyme that can hydrolyze chitin. It plays a significant role in plant defense mechanisms because it can break down the cell walls of plant pathogens, inducing a defense response in plants [96]. This makes it a crucial component in the plant’s resistance to both pathogens and pests [96,97]. Cao conducted a comprehensive whole-genome analysis to identify chitinase family genes in A. nanus, revealing that specific genes such as EVM0022783, EVM0020238, and EVM0003645 are notably upregulated in response to both low-temperature and osmotic stress conditions [98]. In our previous research, we discovered that AnWRKY29 can synergize with AnHSP90 to influence the synthesis of trehalose, thereby enhancing the A. nanus drought resistance [84]. Additionally, it has been reported that trehalose can augment the insect resistance of plants [99]. Therefore, the question of whether an increase in trehalose synthesis under arid conditions can contribute to the enhancement of A. nanus insect resistance remains a subject of potential investigation. This suggests that the physiological responses and signaling components induced by external environmental factors in Ammopiptanthus share commonalities. This commonality could form the basis for the development of cross-resistance when responding to various stressors and is also a manifestation of resource conservation in plants. These studies confirm our perspective.
Previous research by our team has also confirmed this hypothesis. When A. nanus were exposed to MeJA, it was observed that their salt tolerance increased [100]. Compared to control plants, the fumigated plants had reduced K+ efflux from the roots, significantly increased Na+ efflux, and significantly increased H+ efflux [100]. The results indicated that proper concentration of MeJA treatment could increase the salt resistance of the Ammopiptanthus seedlings [100]. This also suggests that JA, a key plant hormone associated with anti-insect defense, may play an important role in Ammopiptanthus when responding to both insect herbivory and salt stress. Recent studies indicate that pre-treatment with herbivory by O. ericae increases JA levels in A. nanus. As a result, H+-ATPase is activated [4]. When compared to plants not exposed to herbivory, the induction of NaCl increases the efflux of H+, reduces K+ efflux, and increases Na+ efflux, subsequently enhancing the plant’s K+/Na+ ratio [4]. This indicates that insect herbivory on Ammopiptanthus can activate the key enzyme OPR3 in the JA pathway, inducing JA accumulation. The activation of the JA pathway plays a role in regulating the selective absorption of Ca2+, H+, K+, and Na+ in the roots. When Ammopiptanthus roots encounter salt stress, plants that have undergone insect herbivory accumulate Ca2+ rapidly. The activity of the plasma membrane H+-ATPase is activated, leading to enhanced efflux of H+, providing a proton motive force for the Na+/H+ antiporter to pump excess Na+ out of the cell. Simultaneously, the substantial efflux of H+ might mitigate the depolarization of the plasma membrane caused by salt stress, reducing the outward flow of K+. This helps maintain the intracellular K+/Na+ ratio, promoting ionic balance in the cytoplasm and enhancing Ammopiptanthus’ adaptation to salt stress [4]. Previous studies have shown that SA plays a crucial role in the plant’s response to biotic stress [101,102]. It is typically associated with the plant’s immune system. When plants are attacked by pathogens such as bacteria, fungi, viruses, or pests, the synthesis and accumulation of SA increase. This activation triggers the plant’s systemic acquired resistance (SAR), enhancing its resistance to pathogens. Plants may need adaptive adjustments between biotic and abiotic stresses [103]. SA may play a pivotal role in this process, regulating the plant’s responses to better adapt to multiple stressors. Yang et al. [64] have elucidated the robust adaptability of Ammopiptanthus in response to extreme temperature stress through physiological and transcriptomic analyses. Under conditions of high temperature, there is a significant fluctuation in the concentrations of plant hormones such as ABA, SA, and cytokinins (CKs), which play pivotal roles in the plant’s thermotolerance mechanisms [64].
Arbuscular mycorrhizal fungi (AMF) are a type of fungi that form a symbiotic relationship with various plants, including those that grow in nutrient-poor soils like Ammopiptanthus. These fungi, including endomycorrhiza and ectomycorrhiza, interact with plant roots to establish a mutually beneficial symbiosis [104,105]. For Ammopiptanthus, which grow in nutrient-deficient soils, mycorrhizal associations are crucial for nutrient absorption and water uptake [106]. AMF and Rhizobium bacteria are two important types of symbiotic microorganisms that can interact with plant roots to form arbuscular mycorrhiza and root nodules, respectively [107]. AMF establish mutualistic relationships with a wide range of plants, aiding in nutrient uptake, while rhizobium bacteria form symbiotic relationships with leguminous plants, assisting in nitrogen fixation [108,109,110]. Recently, dark septate endophytes (DSEs) have also received more attention. Li et al. [111] isolated DSE from Gymnocarpos przewalskii and established a symbiotic relationship with Ammopiptanthus, leading to increased root biomass and shoot growth, which is beneficial for its adaptation to arid environments [111]. Through interactions with fungi, Ammopiptanthus triggers a series of biochemical responses, including the synthesis of secondary metabolites, some of which have toxic or antagonistic effects against potential pathogens [111]. Hou et al. [112] explores the effects of DSEs from heavy-metal habitats on non-host Medicago sativa and A. mongolicus under cadmium stress, finding that DSEs can enhance plant growth, nutrient uptake, and tolerance to Cd, with potential for soil remediation in heavy metal-polluted ecosystems [112]. These findings enhance our comprehension of the interactions between dark septate endophytes and plants within environments polluted by heavy metals. Furthermore, symbiotic fungi limit the invasion and growth of pathogenic microorganisms through mechanisms such as physical barriers or competitive exclusion. In summary, the mutualistic symbiosis between Ammopiptanthus and root-associated fungi is an important area of ecological and biological research, helping deepen our understanding of plant-microbe interactions. It also provides valuable insights for the restoration and protection of ecosystems in arid regions. This symbiotic relationship plays a critical role in enhancing Ammopiptanthus’ survival and ecosystem stability.
Indeed, the concept of cross-resistance or cross-tolerance highlights how plants can adapt more effectively to a variety of stresses, thereby promoting ecosystem stability. This further supports our perspective that since a plant’s own resources are limited, there are commonalities in the physiological responses and signaling components induced by various environmental factors (Table 2). These commonalities likely form the basis for the development of cross-resistance when plants respond to different stresses (in Figure 3).
5. Conclusions
Ammopiptanthus, the only evergreen broadleaf shrub in the arid desert regions of northwestern China, plays a crucial role in the natural ecosystems of this area. Current research on Ammopiptanthus mainly focuses on the following aspects: (1) studies on its resistance physiology and molecular mechanisms; (2) natural population distribution, conservation, and utilization; and (3) investigations into mycorrhizal fungi resources and symbiotic mechanisms. In the future, for the rare species like Ammopiptanthus, there are two main aspects to consider. On one hand, there should be increased efforts to protect the wild natural populations and establish conservation areas beyond natural reserves [63]. On the other hand, it is essential to continue researching the stress resistance mechanisms of Ammopiptanthus. More attention should be given to the crosstalk of signals between various stressors, such as the interaction between biotic and abiotic stress resistance. The role of mycorrhizal fungi in aiding Ammopiptanthus in resisting various stress factors should also be further investigated. Some studies are also devoted to improving the vegetation recovery of Ammopiptanthus by regulating the supply of water, nitrogen, and phosphorus [63]. Looking ahead, it is crucial to build a comprehensive stress signaling network for Ammopiptanthus. This will greatly aid in the identification and utilization of key stress resistance genes, potentially leading to the enhancement of crop resistance and productivity through genetic improvement using these genes.
Author Contributions
Acquired funding, contributed to writing and editing, supervised, Y.C.; wrote the manuscript, S.W.; contributed to writing and editing, supervision, S.L.; conceived the study and reviewed the literature, Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Guangxi Natural Science Foundation grant (2024GXNSFAA010144) and Guangxi Natural Science Foundation grant (2023GXNSFBA026168).
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Yingbai Shen from Beijing Forestry University for his guidance in writing our manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Gao, F.; Wang, X.; Li, X.; Xu, M.; Li, H.; Merhaba, A.; Sun, H.; Wei, S.; Feng, J.; Zhou, Y. Long-read sequencing and de novo genome assembly of Ammopiptanthus nanus, a desert shrub. GigaScience 2018, 7, giy074. [Google Scholar] [PubMed]
- Du, Z.; He, Y.; Wang, H.; Wang, C.; Duan, Y. Potential geographical distribution and habitat shift of the genus Ammopiptanthus in China under current and future climate change based on the MaxEnt model. J. Arid. Environ. 2021, 184, 104328. [Google Scholar] [CrossRef]
- Li, R.; Yan, C.; Zhao, Y.; Wang, P.; Qiu, G.Y. Discriminating growth stages of an endangered mediterranean relict plant (Ammopiptanthus mongolicus) in the arid Northwest China using hyperspectral measurements. Sci. Total Environ. 2019, 657, 270–278. [Google Scholar] [PubMed]
- Chen, Y.; Cao, C.; Guo, Z.; Zhang, Q.; Li, S.; Zhang, X.; Gong, J.; Shen, Y. Herbivore exposure alters ion fluxes and improves salt tolerance in a desert shrub. Plant Cell Environ. 2020, 43, 400–419. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Yu, H.; Liu, Y.; Jiao, P.; Zhou, S.; Zhang, S.; Li, W.; Fu, F. Heterologous expression of antifreeze protein gene AnAFP from Ammopiptanthus nanus enhances cold tolerance in Escherichia coli and tobacco. Gene 2014, 539, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhou, X.; Wang, Y.; Zhou, S.; Fu, F.; Li, W. A betaine aldehyde dehydrogenase gene from Ammopiptanthus nanus enhances tolerance of Arabidopsis to high salt and drought stresses. Plant Growth Regul. 2017, 83, 265–276. [Google Scholar] [CrossRef]
- Lu, C.; Yin, L.; Li, K. Proteome expression patterns in the stress tolerant evergreen Ammopiptanthus nanus under conditions of extreme cold. Plant Growth Regul. 2010, 62, 65–70. [Google Scholar] [CrossRef]
- Chen, S.; Geng, X.; Lou, J.; Huang, D.; Mao, H.; Lin, X. Overexpression of a plasmalemma Na+/H+ antiporter from the halophyte Nitraria sibirica enhances the salt tolerance of transgenic poplar. Plant Sci. 2024, 343, 112061. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, L.; Chen, L.; Ma, H.; Ruan, Y.; Xu, T.; Xu, C.; He, Y.; Qi, M. Molecular cloning and expression of an encoding galactinol synthase gene (AnGolS1) in seedling of Ammopiptanthus nanus. Sci. Rep. 2016, 6, 36113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Bai, L.; Zhang, L.; Chen, G.; Fan, J.; Xu, S.; Guo, Z. Identification and characterization of AnICE1 and AnCBFs involved in cold tolerance from Ammopiptanthus nanus. Plant Physiol. Biochem. 2021, 168, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Mahboob, M.G.; Arena, C.; Kader, M.A.; Sultana, S.; Hasan, A.K.; Wang, J.; Sarker, T.; Zhang, R.; Barmon, M. The modulation of water, nitrogen, and phosphorous supply for growth optimization of the evergreen shrubs Ammopiptanthus mongolicus for revegetation purpose. Front. Plant Sci. 2021, 12, 766523. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Liu, Q.; Liu, F.; Zheng, L.; Bing, J.; Zhou, Y.; Gao, F. Gene profiling of the ascorbate oxidase family genes under osmotic and cold stress reveals the role of AnAO5 in cold adaptation in Ammopiptanthus nanus. Plants 2023, 12, 677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Chen, Y.; Zhang, H.; Shen, Y. The role of 1-penten-3-one in plant defense responses. Plant Physiol. J. 2019, 55, 225–231. [Google Scholar] [CrossRef]
- Eulgem, T. Regulation of the Arabidopsis defense transcriptome. Trends Plant Sci. 2005, 10, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Yu, Y.; Yuan, Y.; Huang, H.; Yan, C. Genetic diversity and geographic differentiation in endangered Ammopiptanthus (Leguminosae) populations in desert regions of northwest China as revealed by ISSR analysis. Ann. Bot. 2005, 95, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Fleischer, R.C.; Tarr, C.L.; Pratt, T.K. Genetic structure and mating system in the palila, an endangered Hawaiian honeycreeper, as assessed by DNA fingerprinting. Mol. Ecol. 2008, 3, 383–392. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, X.; Feng, B. The types of pests and their damage areas on the endangered plant Ammopiptanthus nanus. In Proceedings of the Congress and Symposium on the 60th Anniversary of the Entomological Society of China, Chongqing, China, 9–13 November 2004. [Google Scholar]
- Li, X.; Yang, Y.; Zhang, D.; Mu, T.; Yu, F.; Zhou, Y. Damage rate and spatial distribution type of Etiella zinckenella on Ammopiptanthus in different habitats. North Hortic. 2016, 24, 108–111. [Google Scholar]
- Oliveira, J.L.D.; Campos, E.V.R.; Bakshi, M.; Abhilash, P.C.; Fraceto, L.F. Application of nanotechnology for the encapsulation of botanical insecticides for sustainable agriculture: Prospects and promises. Biotechnol. Adv. 2014, 32, 1550–1561. [Google Scholar] [CrossRef] [PubMed]
- Khursheed, A.; Rather, M.A.; Jain, V.; Wani, A.R.; Rasool, S.; Nazir, R.; Malik, N.A.; Majid, S.A. Plant based natural products as potential ecofriendly and safer biopesticides: A comprehensive overview of their advantages over conventional pesticides, limitations and regulatory aspects. Microb. Pathog. 2022, 173, 105854. [Google Scholar] [CrossRef]
- Zebelo, S.A.; Maffei, M.E. Role of early signalling events in plant-insect interactions. J. Exp. Bot. 2015, 66, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Maischak, H.; Grigoriev, P.A.; Vogel, H.; Boland, W.; Mithofer, A. Oral secretions from herbivorous lepidopteran larvae exhibit ion channel-forming activities. FEBS Lett. 2007, 581, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Arimura, G.; Mithofer, A. Natural elicitors, effectors and modulators of plant responses. Nat. Prod. Rep. 2012, 29, 1288–1303. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E.; Mithofer, A.; Boland, W. Before gene expression: Early events in plant-insect interaction. Trends Plant Sci. 2007, 12, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Reymond, P. Receptor kinases in plant responses to herbivory. Curr. Opin. Biotechnol. 2021, 70, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.R.; Mithofer, A.; Will, T.; Felle, H.H.; Furch, A.C. Herbivore-triggered electrophysiological reactions: Candidates for systemic signals in higher plants and the challenge of their identification. Plant Physiol. 2016, 170, 2407–2419. [Google Scholar] [CrossRef] [PubMed]
- Furstenberg-Hagg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Maffei, M.E. Calcium and secondary CPK signaling in plants in response to herbivore attack. Biochem. Biophys. Res. Commun. 2010, 400, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Fan, M.; Yang, M.; Zhao, J.; Zhang, W.; Su, Y.; Xiao, L.; Deng, H.; Xie, D. Injury activates Ca2+/Calmodulin-Dependent Phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis. Mol. Cell 2018, 70, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A.; Engelberth, J.; Alborn, H.T.; Tumlinson, J.H., III.; Teal, P.E.A. Phytohormone-based activity mapping of insect herbivore-produced elicitors. Proc. Natl. Acad. Sci. USA 2009, 106, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Zhang, J.; Shen, Y. Regurgitant from Orgyia ericae Germar induces calcium influx and accumulation of hydrogen peroxide in Ammopiptanthus mongolicus (Maxim. ex Kom.) Cheng f. cells. Acta Ecol. Sin. 2012, 32, 6520–6526. [Google Scholar] [CrossRef]
- Waadt, R.; Seller, C.A.; Hsu, P.-K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, Y.-T.; Liu, Y.-X.; Hao, G.-F.; Yang, X.-Q. Molecular interaction network of plant-herbivorous insects. Adv. Agrochem. 2024, 3, 74–82. [Google Scholar] [CrossRef]
- Kallure, G.S.; Kumari, A.; Shinde, B.A.; Giri, A.P. Characterized constituents of insect herbivore oral secretions and their influence on the regulation of plant defenses. Phytochemistry 2022, 193, 113008. [Google Scholar] [CrossRef] [PubMed]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 2019, 70, 527–557. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Shen, Y.; Huang, Q. Influx of Ca2+-dependent H+ to suspended cells of Ammopiptanthus mongolicus triggered by mechanical stimulation. Sci. Silvae Sin. 2012, 48, 36–41. [Google Scholar] [CrossRef]
- Li, Y.; Zeng, H.; Xu, F.; Yan, F.; Xu, W. H+-ATPases in plant growth and stress responses. Annu. Rev. Plant Biol. 2022, 73, 495–521. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, C.; Xue, Y.; Liu, X.; Chen, S.; Song, C.; Yang, Y.; Guo, Y. Calcium-activated 14-3-3 proteins as a molecular switch in salt stress tolerance. Nat. Commun. 2019, 10, 1199. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Han, X.; Ma, L.; Wu, Y.; Liu, X.; Fu, H.; Liu, G.; Lei, X.; Guo, Y. Dynamic changes of phosphatidylinositol and phosphatidylinositol 4-phosphate levels modulate H+-ATPase and Na+/H+ antiporter activities to maintain ion homeostasis in Arabidopsis under salt stress. Mol. Plant 2021, 14, 2000–2014. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, H.; Zhang, T.; Cao, C.; Zong, S.; Luo, Y.; Shen, Y. Metabolites of Ammopiptanthus mongolicus induced by Orgyia ericae attack and mechanical wounding. Plant Physiol. Biochem. 2013, 69, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, X.; Cao, C.; Mei, X.; Wang, N.; Yan, S.; Zong, S.; Luo, Y.; Yang, H.; Shen, Y. Similar metabolic changes induced by HIPVs exposure as herbivore in Ammopiptanthus mongolicus. PLoS ONE 2014, 9, e95474. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wang, M.; Wu, M.; Li, X.; Liu, H.; Niu, N.; Li, S.; Chen, L. Volatile organic compounds (VOCs) from plants: From release to detection. TrAC Trend Anal. Chem. 2023, 158, 116872. [Google Scholar] [CrossRef]
- Heil, M. Herbivore-induced plant volatiles: Targets, perception and unanswered questions. New Phytol. 2014, 204, 297–306. [Google Scholar] [CrossRef]
- Hao, X.; Wang, S.; Fu, Y.; Liu, Y.; Shen, H.; Jiang, L.; McLamore, E.S.; Shen, Y. The WRKY46–MYC2 module plays a critical role in E-2-hexenal-induced anti-herbivore responses by promoting flavonoid accumulation. Plant Commun. 2023, 5, 100734. [Google Scholar] [CrossRef]
- Xu, G.; Pan, B.; Mingling, X. A study on alkaloids of Ammopiptanthus mongolicus. Arid. Zone Res. 1994, 11, 50–52. [Google Scholar]
- Gao, W.; Zhu, G.; Liu, Q. On the nematocidal activity of Ammopiptanthus mongolicus and Peganum harmala L against Bursaphelenchus xylophilus. J. Tianjin Norm. Univ. 2009, 29, 55–58. [Google Scholar] [CrossRef]
- Lei, X.; He, D.; He, Y.; Ma, Y. Inhibitory effects of the methanol extracts from the caudices of Ammopiptanthus mongolicus on development of the larvae of the diamondback moth, Plutella xylostella. Plant Prot. 2008, 34, 100–103. [Google Scholar] [CrossRef]
- Poelman, E.H.; Broekgaarden, C.; Van Loon, J.J.A.; Dicke, M. Early season herbivore differentially affects plant defence responses to subsequently colonizing herbivores and their abundance in the field. Mol. Ecol. 2008, 17, 3352–3365. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Saona, C.; Chalmers, J.A.; Raj, S.; Thaler, J.S. Induced plant responses to multiple damagers: Differential effects on an herbivore and its parasitoid. Oecologia 2005, 143, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, X.; Shen, Y.; Yang, H. Metabolic effects of early-season herbivores on the response to conspecific attack in later-season in Ammopiptanthus mongolicus (Maxim.) Cheng f. Plant Physiol. J. 2014, 50, 61–67. [Google Scholar] [CrossRef]
- Wari, D.; Aboshi, T.; Shinya, T.; Galis, I. Integrated view of plant metabolic defense with particular focus on chewing herbivores. J. Integr. Plant Biol. 2022, 64, 449–475. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2019, 48, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Liao, Y.; Jian, G.; Jia, Y.; Zeng, L.; Gu, D.; Li, H.; Yang, Y. Light induces an increasing release of benzyl nitrile against diurnal herbivore Ectropis grisescens Warren attack in tea (Camellia sinensis) plants. Plant Cell Environ. 2023, 46, 3464–3480. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Moeder, W.; Yoshioka, K.; Shan, L. A tale of many families: Calcium channels in plant immunity. Plant Cell 2022, 34, 1551–1567. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Xi, J.; Du, L.; Suttle, J.C.; Poovaiah, B.W. Coupling calcium/calmodulin-mediated signaling and herbivore-induced plant response through calmodulin-binding transcription factor AtSR1/CAMTA3. Plant Mol. Biol. 2012, 79, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Kloth, K.J.; Wiegers, G.L.; Busscher-Lange, J.; van Haarst, J.C.; Kruijer, W.; Bouwmeester, H.J.; Dicke, M.; Jongsma, M.A. AtWRKY22 promotes susceptibility to aphids and modulates salicylic acid and jasmonic acid signalling. J. Exp. Bot. 2016, 67, 3383–3396. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Yang, H.; Lu, Q.; Zhang, X. Endemic shrubs in temperate arid and semiarid regions of northern China and their potentials for rangeland restoration. AoB Plants 2015, 7, plv063. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Guo, M.; Yue, G.; Li, J.; Yang, S.; Zhao, P.; Su, Y. An unusual strategy of stomatal control in the desert shrub Ammopiptanthus mongolicus. Plant Physiol. Biochem. 2018, 125, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zhang, J.; Xin, Z.; Huang, Y.; Han, C.; Li, Y.; Lu, Q. Ecological stoichiometric characteristics in organs of Ammopiptanthus mongolicus in different habitats. Plants 2023, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z. Probe into drought-resisting mechanism of Ammopitanthus mongolicus (Maxim) Cheng F. J. Desert Res. 2000, 20, 71–74. [Google Scholar] [CrossRef]
- Wang, H.; Jia, G.; Ding, Q. Research progress of abiotic stress tolerant mechanisms and application prospect of Ammopiptanthus mongolicus Maxim. Chin. Agric. Sci. Bull. 2005, 21, 121–125. [Google Scholar] [CrossRef]
- Zhou, Y.; Liu, C.; Feng, J.; Jia, X. Advances of dhought-resistance and frigid-resistance mechanism recearch on Ammopiptanthus mongolicus. J. Desert Res. 2001, 21, 312–316. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Lu, T.; Wang, J.; Shi, W. Identification of the efficacy of ex situ conservation of Ammopiptanthus nanus based on its ETS-SSR markers. Plants 2023, 12, 2670. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Liu, Y.; Han, H.; Zhao, X.; Chen, S.; Li, G.; Shi, S.; Feng, J. Physiological and transcriptome analyses reveal the response of Ammopiptanthus mongolicus to extreme seasonal temperatures in a cold plateau desert ecosystem. Sci. Rep. 2022, 12, 10630. [Google Scholar] [CrossRef] [PubMed]
- Lu, G.; Wang, L.; Zhou, L.; Su, X.; Guo, H.; Cheng, H. Overexpression of AmCBF1 enhances drought and cold stress tolerance, and improves photosynthesis in transgenic cotton. PeerJ 2022, 10, e13422. [Google Scholar] [CrossRef] [PubMed]
- Fei, Y.B.; Cao, P.X.; Gao, S.Q.; Wang, B.; Wei, L.B.; Zhao, J.; Chen, G.; Wang, B.H. Purification and structure analysis of antifreeze proteins from Ammopiptanthus mongolicus. Prep. Biochem. Biotechnol. 2008, 38, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wei, L.; Wang, G. Multistep purification of an antifreeze protein from Ammopiptanthus mongolicus by chromatographic and electrophoretic methods. J. Chromatogr. Sci. 2003, 41, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wei, L. Purification of boiling-soluble antifreeze protein from the Legume Ammopiptanthus mongolicus. Prep. Biochem. Biotechnol. 2003, 33, 67–80. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signalling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Al Azzawi, T.N.I.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. The key roles of ROS and RNS as a signaling molecule in plant–microbe interactions. Antioxidants 2023, 12, 268. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Niu, M.X.; Feng, C.H.; He, F.; Zhang, H.; Bao, Y.; Liu, S.J.; Liu, X.; Su, Y.; Liu, C.; Wang, H.L.; et al. The miR6445-NAC029 module regulates drought tolerance by regulating the expression of glutathione S-transferase U23 and reactive oxygen species scavenging in Populus. New Phytol. 2024, 242, 2043–2058. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Liu, M.; Shi, J.; Zheng, G.; Wang, Y.; Wang, J.; Chen, Y.; Lu, C.; Yin, W. Reference gene selection for qPCR in Ammopiptanthus mongolicus under abiotic stresses and expression analysis of seven ROS-scavenging enzyme genes. Plant Cell Rep. 2012, 31, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Zheng, L.; Cao, S.; Liu, Q.; Wei, S.; Zhou, Y.; Gao, F. AnDREB5.1, a A5 group DREB gene from desert shrub Ammopiptanthus nanus, confers osmotic and cold stress tolerances in transgenic tobacco. Physiol. Plant. 2024, 176, e14272. [Google Scholar] [CrossRef] [PubMed]
- Dorjee, T.; Cui, Y.; Zhang, Y.; Liu, Q.; Li, X.; Sumbur, B.; Yan, H.; Bing, J.; Geng, Y.; Zhou, Y.; et al. Characterization of NAC gene family in Ammopiptanthus mongolicus and functional analysis of AmNAC24, an osmotic and cold-stress-induced NAC gene. Biomolecules 2024, 14, 182. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhao, H.; Miao, Y.; Li, G. Impacts of low temperature on photosynthesis physiology of Ammopiptanthus nanus (Leguminosae) and Ammopiptanthus mongolicus seedlings. J. Xinjiang Univ. 2009, 5, 342–346. [Google Scholar] [CrossRef]
- Han, L.; Li, J.; Jin, M.; Su, Y. Functional analysis of a type 2C protein phosphatase gene from Ammopiptanthus mongolicus. Gene 2018, 653, 29–42. [Google Scholar] [CrossRef]
- Klotke, J.; Kopka, J.; Gatzke, N.; Heyer, A.G. Impact of soluble sugar concentrations on the acquisition of freezing tolerance in accessions of Arabidopsis thaliana with contrasting cold adaptation—Evidence for a role of raffinose in cold acclimation. Plant Cell Environ. 2004, 27, 1395–1404. [Google Scholar] [CrossRef]
- Kito, K.; Yamane, K.; Yamamori, T.; Matsuhira, H.; Tanaka, Y.; Takabe, T. Isolation, functional characterization and stress responses of raffinose synthase genes in sugar beet. J. Plant Biochem. Biotechnol. 2017, 27, 36–45. [Google Scholar] [CrossRef]
- Yu, H.Q.; Yong, T.M.; Li, H.J.; Liu, Y.P.; Zhou, S.F.; Fu, F.L.; Li, W.C. Overexpression of a phospholipase Dα gene from Ammopiptanthus nanus enhances salt tolerance of phospholipase Dα1-deficient Arabidopsis mutant. Planta 2015, 242, 1495–1509. [Google Scholar] [CrossRef]
- Cui, H.; Wang, Y.; Yu, T.; Chen, S.; Chen, Y.; Lu, C. Heterologous expression of three Ammopiptanthus mongolicus dehydrin genes confers abiotic stress tolerance in Arabidopsis thaliana. Plants 2020, 9, 193. [Google Scholar] [CrossRef]
- Fu, X.; Xiao, Z.; Gao, F.; Zhou, Y. Proteomics analysis of Ammopiptanthus mongolicus leaves under drought stress. Biotechnol. Bull. 2017, 33, 69–80. [Google Scholar] [CrossRef]
- Wang, S.; Liu, Y.; Hao, X.; Wang, Z.; Chen, Y.; Qu, Y.; Yao, H.; Shen, Y. AnWRKY29 from the desert xerophytic evergreen Ammopiptanthus nanus improves drought tolerance through osmoregulation in transgenic plants. Plant Sci. 2023, 336, 111851. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Hao, X.; Liu, Y.; Chen, Y.; Qu, Y.; Wang, Z.; Shen, Y. AnWRKY29 and AnHSP90 synergistically modulate trehalose levels in a desert shrub leaves during osmotic stress. Physiol. Plant. 2024, 176, e14237. [Google Scholar] [CrossRef] [PubMed]
- Sumbur, B.; Zhou, M.; Dorjee, T.; Bing, J.; Ha, S.; Xu, X.; Zhou, Y.; Gao, F. Chemical and transcriptomic analyses of leaf cuticular wax metabolism in Ammopiptanthus mongolicus under osmotic stress. Biomolecules 2024, 14, 227. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Bobadilla, M.; Vitiello, A.; Erb, M.; Poelman, E.H. Plant defense strategies against attack by multiple herbivores. Trends Plant Sci. 2022, 27, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Ryu, C.M. Insect stings to change gear for healthy plant: Improving maize drought tolerance by whitefly infestation. Plant Signal. Behav. 2016, 11, e1179420. [Google Scholar] [CrossRef] [PubMed]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kannaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenstrom, E.; Niinemets, U. Drought-tolerance of wheat improved by rhizosphere bacteria from harsh environments: Enhanced biomass production and reduced emissions of stress volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Xu, X.; Luo, Y.; Gong, Z.; Hu, X.; Wu, M.; Liu, Y.; Yan, F.; Zhang, X.; Zhang, W.; et al. R2R3 MYB-dependent auxin signalling regulates trichome formation, and increased trichome density confers spider mite tolerance on tomato. Plant Biotechnol. J. 2021, 19, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Sun, L.; Dong, M.; Fan, S.; Shi, K.; Qu, Y.; Zhu, L.; Shi, J.; Wang, W.; Liu, Y.; et al. Novel players in organogenesis and flavonoid biosynthesis in cucumber glandular trichomes. Plant Physiol. 2023, 192, 2723–2736. [Google Scholar] [CrossRef]
- Wang, X.; Li, J.; Li, L.; Zhang, F.; Wang, M. Cloning and expression analysis of a trypsin inhibitor gene AmTI from Ammopiptanthus mongolicus. J. Inn. Mong. Agr. Univ. 2012, 33, 103–108. [Google Scholar] [CrossRef]
- Green, T.R.; Ryan, C.A. Wound-induced proteinase inhibitor in plant leaves: A possible defense mechanism against insects. Science 1972, 175, 776–777. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Cheng, J.; Yin, M.; Wu, J. NaMLP, a new identified Kunitz trypsin inhibitor regulated synergistically by JA and ethylene, confers Spodoptera litura resistance in Nicotiana attenuata. Plant Cell Rep. 2023, 42, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.; Song, N.; Chen, S.; Wu, J. NaKTI2, a Kunitz trypsin inhibitor transcriptionally regulated by NaWRKY3 and NaWRKY6, is required for herbivore resistance in Nicotiana attenuata. Plant Cell Rep. 2020, 40, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Collinge, D.B.; Kragh, K.M.; Mikkelsen, J.D.; Nielsen, K.K.; Rasmussen, U.; Vad1, K. Plant chitinases. Plant J. 1993, 3, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Alves, P.C.M.S.; Ahmad, I.; Gaffoor, I.; Acevedo, F.E.; Peiffer, M.; Jin, S.; Han, Y.; Shakeel, S.; Felton, G.W.; et al. Turnabout is fair play: Herbivory-induced plant chitinases excreted in fall armyworm frass suppress herbivore defenses in maize. Plant Physiol. 2016, 171, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, Y.; Li, Z.; Shi, W.; Gao, F.; Zhou, Y.; Zhang, G.; Feng, J. Genome-wide identification and expression analyses of the chitinases under cold and osmotic stress in Ammopiptanthus nanus. Genes 2019, 10, 472. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Louis, J.; Ayre, B.G.; Reese, J.C.; Pegadaraju, V.; Shah, J. TREHALOSE PHOSPHATE SYNTHASE11-dependent trehalose metabolism promotes Arabidopsis thaliana defense against the phloem-feeding insect Myzus persicae. Plant J. 2011, 67, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gao, H.; Shen, Y. Effects of MeJA fumigation to leaves on ion flux in apical region of Ammopiptanthus nanus and salt resistance. Plant Physiol. J. 2013, 50, 477–482. [Google Scholar] [CrossRef]
- Mishra, S.; Roychowdhury, R.; Ray, S.; Hada, A.; Kumar, A.; Sarker, U.; Aftab, T.; Das, R. Salicylic acid (SA)-mediated plant immunity against biotic stresses: An insight on molecular components and signaling mechanism. Plant Stress. 2024, 11, 100427. [Google Scholar] [CrossRef]
- Nawaz, M.; Sun, J.; Shabbir, S.; Khattak, W.A.; Ren, G.; Nie, X.; Bo, Y.; Javed, Q.; Du, D.; Sonne, C. A review of plants strategies to resist biotic and abiotic environmental stressors. Sci. Total Environ. 2023, 900, 165832. [Google Scholar] [PubMed]
- Chen, J.; Clinton, M.; Qi, G.; Wang, D.; Liu, F.; Fu, Z.Q.; Spoel, S. Reprogramming and remodeling: Transcriptional and epigenetic regulation of salicylic acid-mediated plant defense. J. Exp. Bot. 2020, 71, 5256–5268. [Google Scholar] [CrossRef] [PubMed]
- Gutjahr, C.; Gobbato, E.; Choi, J.; Riemann, M.; Johnston, M.G.; Summers, W.; Carbonnel, S.; Mansfield, C.; Yang, S.-Y.; Nadal, M.; et al. Rice perception of symbiotic arbuscular mycorrhizal fungi requires the karrikin receptor complex. Science 2015, 350, 1521–1524. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M.; Zobel, M. How mycorrhizal associations drive plant population and community biology. Science 2020, 367. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Xie, T.; Wang, Y.; Yang, J.; Yang, X. Growth promotion and mechanism of arbuscular mycorrhizal fungi (AMF) on Ammopiptanthus mongolicus seedlings. Arid. Zone Res. 2023, 40, 78–89. [Google Scholar]
- Ma, J.; Wang, W.; Yang, J.; Qin, S.; Yang, Y.; Sun, C.; Pei, G.; Zeeshan, M.; Liao, H.; Liu, L.; et al. Mycorrhizal symbiosis promotes the nutrient content accumulation and affects the root exudates in maize. BMC Plant Biol. 2022, 22, 64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, J.; Zhu, H. Genetic and molecular mechanisms underlying symbiotic specificity in legume-rhizobium interactions. Front. Plant Sci. 2018, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, J.P.; Oresnik, I.J. The rhizobium-legume symbiosis: Co-opting successful stress management. Front. Plant Sci. 2022, 12, 796045. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, X.; Huo, H.; Yuan, G.; Sun, Y.; Zhang, D.; Cao, Y.; Xu, L.; Wei, G. Phylogenetic diversity of Ammopiptanthus rhizobia and distribution of rhizobia associated with Ammopiptanthus mongolicus in diverse regions of northwest China. Microb. Ecol. 2016, 72, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, X.; Hou, L.; Ren, Y.; Wang, S.; Su, F. Dark septate endophytes isolated from a xerophyte plant promote the growth of Ammopiptanthus mongolicus under drought condition. Sci. Rep. 2018, 8, 7896. [Google Scholar] [CrossRef]
- Hou, L.; Yu, J.; Zhao, L.; He, X. Dark septate endophytes improve the growth and the tolerance of medicago sativa and Ammopiptanthus mongolicus under cadmium stress. Front. Microbiol. 2020, 10, 3061. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Pest control methods (drawn by Figdraw).
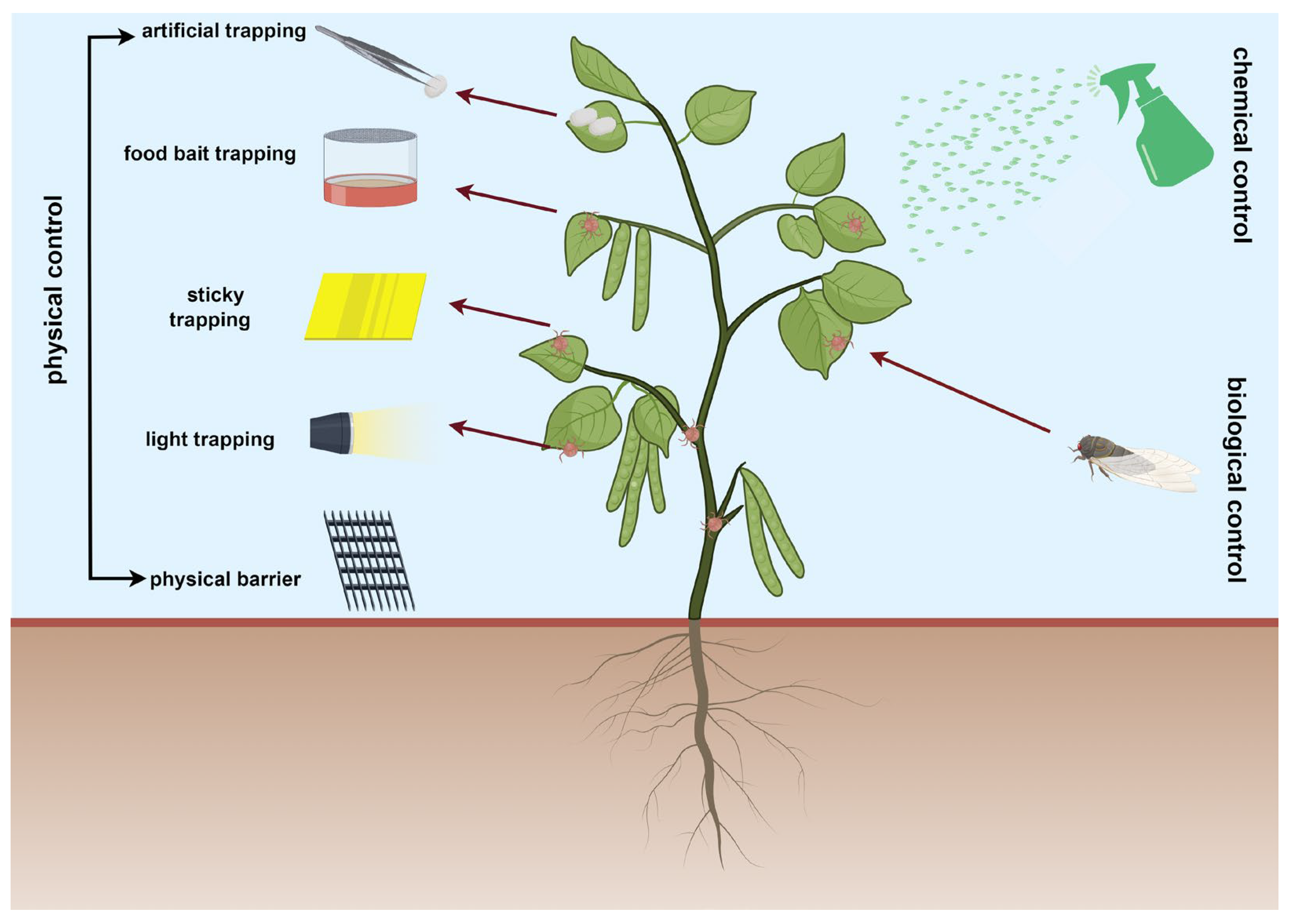
Figure 2.
Early biotic stress signaling in Ammopiptanthus. OS, oral secretion; Ca2+ receptor: calmodulin (CaM), calmodulin-like proteins (CMLs), calcineurin B-like proteins (CBLs), calcium-dependent protein kinases (CPKs), calcium/calmodulin-dependent protein kinases (CDPKs); JA, jasmonic acid; LOX, lipoxygenase; TF, transcription factor).
Figure 2.
Early biotic stress signaling in Ammopiptanthus. OS, oral secretion; Ca2+ receptor: calmodulin (CaM), calmodulin-like proteins (CMLs), calcineurin B-like proteins (CBLs), calcium-dependent protein kinases (CPKs), calcium/calmodulin-dependent protein kinases (CDPKs); JA, jasmonic acid; LOX, lipoxygenase; TF, transcription factor).

Figure 3.
Ammopiptanthus exhibits interactive resistance when responding to external biotic and abiotic stressors. (PRR, pattern recognition receptor; SA, salicylic acid; JA, jasmonic acid; ROS, reactive oxygen species; SOD, superoxide dismutase; POD, peroxidase; APX, ascorbate peroxidase; TIs, trypsin inhibitors) (drawn by Figdraw).
Figure 3.
Ammopiptanthus exhibits interactive resistance when responding to external biotic and abiotic stressors. (PRR, pattern recognition receptor; SA, salicylic acid; JA, jasmonic acid; ROS, reactive oxygen species; SOD, superoxide dismutase; POD, peroxidase; APX, ascorbate peroxidase; TIs, trypsin inhibitors) (drawn by Figdraw).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Types of ground pests and their damage areas [17].
Table 1.
Types of ground pests and their damage areas [17].
| Order | Family | Feeding Site | Representative Species |
|---|---|---|---|
| Coleooptera | Teneberionidae | Leaves; flowers; young fruits; seeds; roots | Platyope mongolica Faldermann; Anatolica mucronate Reitter; Platyope ordossica Semenow; Anatolica amoenula Reitter; Pterocoma reiterri Frivoldszky et al. |
| Curculionidea | Leaves; flowers; young fruits; seeds; roots | Deracanthus jakovlevi Suvorov; Chloebius psittacinus Boheman et al. | |
| Rutelidae | Leaves; flowers; young fruits | Proagopertha lucidula Faldermann | |
| Melolonthidae | Leaves; flowers; young fruits | Chioneosoma reitteri Semenov | |
| Meloidae | Flowers; young fruits | Mylabris mongolica (Dokhlouroff) | |
| Cleridea | Flowers; young fruits | Trichodes sinae Chevrolat | |
| Chrysomelidae | Leaves; flowers; young fruits | Diorhabda rybakowi Weise | |
| Elateridae | Roots | Pleonomus canaliculatus Faldermann | |
| Homoptera | Psyllidae | Leaves; flowers; young fruits | Psylla mongolicus Loginova |
| Lepidoptera | Lymantriidae | Leaves; young fruits | Orgyia ericae Germar |
| Hemiptera | Lygaeidae | Flowers; young fruits | Lygaeus equestris Linnaeus |
| Pentatomidae | Flowers; young fruits | Rhaphigaster nebulosa Poda; Brachynema germarii Kolenati | |
| Miridae | Flowers | ||
| Orthoptera | Tettigoniidae | Leaves; flowers; young fruits; seeds | |
| Thysanoptera | Thripidae | Flowers |
Table 2.
Synergistic effects of multiple environmental pressures.
| Category | Pathway | Description | Involved | Stresses Addressed |
|---|---|---|---|---|
| Physical Defense | Trichome Formation | Increase density to cope with stress; dense trichomes on leaves and stems act as a physical barrier against pests and pathogens, and adapt to arid environments. | Trichomes | Drought, extreme temperatures, UV radiation, pathogens and pests |
| Ion Regulation | Selective Absorption of Ca2+, H+, K+, and Na+ in Roots | Such activation of H+-ATPase enhances H+ efflux, providing a proton motive force for the Na+/H+ antiporter to pump excess Na+ out of the cell. | H+-ATPase, Na+/H+ antiporter, calcium channel, etc. | Pests, salt stress |
| Chemical Defense | Stress Proteins | Involved in physiological processes induced by various stress factors | Trypsin inhibitors | Dehydration, low temperature, salt stress |
| Dehydrins | Water stress, salt stress, extreme temperatures, pathogen infections | |||
| Plant Defense Enzyme | Synthesis of plant defense enzymes | Chitinase; trehalose phosphate synthase | Pathogens, pests, and extreme temperatures; osmotic stress | |
| Plant Hormones | Plant hormones play pivotal roles in the plant’s response to various stresses and in signaling pathways. | SA, JA, ABA et al. | Water stress, biotic stress, extreme temperatures, salt stress, pathogens, pests | |
| Symbiotic Relationships | Mycorrhizal and Endophytic Symbiosis | Form symbiotic relationships with fungi, enhancing nutrient absorption and adaptation. | AMF, DSE et al. | Drought, pathogens, heavy metal pollution |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, S.; Liang, S.; Liu, Y.; Chen, Y. Advances in Endangered Plant Research: Ammopiptanthus’s Responses to Biotic and Abiotic Stressors. Forests 2024, 15, 890. https://doi.org/10.3390/f15050890
AMA Style
Wang S, Liang S, Liu Y, Chen Y. Advances in Endangered Plant Research: Ammopiptanthus’s Responses to Biotic and Abiotic Stressors. Forests. 2024; 15(5):890. https://doi.org/10.3390/f15050890
Chicago/Turabian StyleWang, Shuyao, Shenghua Liang, Yahui Liu, and Yingying Chen. 2024. "Advances in Endangered Plant Research: Ammopiptanthus’s Responses to Biotic and Abiotic Stressors" Forests 15, no. 5: 890. https://doi.org/10.3390/f15050890
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.