
Breakthrough Infections in SARS-CoV-2-Vaccinated Multiple Myeloma Patients Improve Cross-Protection against Omicron Variants

 , , , ,
, , , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Methods
2.1. Study Design
2.2. Participants
2.3. Procedures
2.4. SARS-CoV-2-Specific IgG Antibodies against Wuhan and Omicron Variants
2.5. Leukocyte and Lymphocyte Counts
2.6. Cellular Immune Responses
2.7. Statistical Methods
3. Results
3.1. Cohort Characteristics
3.2. Reduced Ab Levels and Omicron Cross-Neutralisation in Vaccinated Uninfected MM Patients
3.3. Degree of Vaccine Failure in Uninfected MM Patients Depends on Disease Stage and Treatment
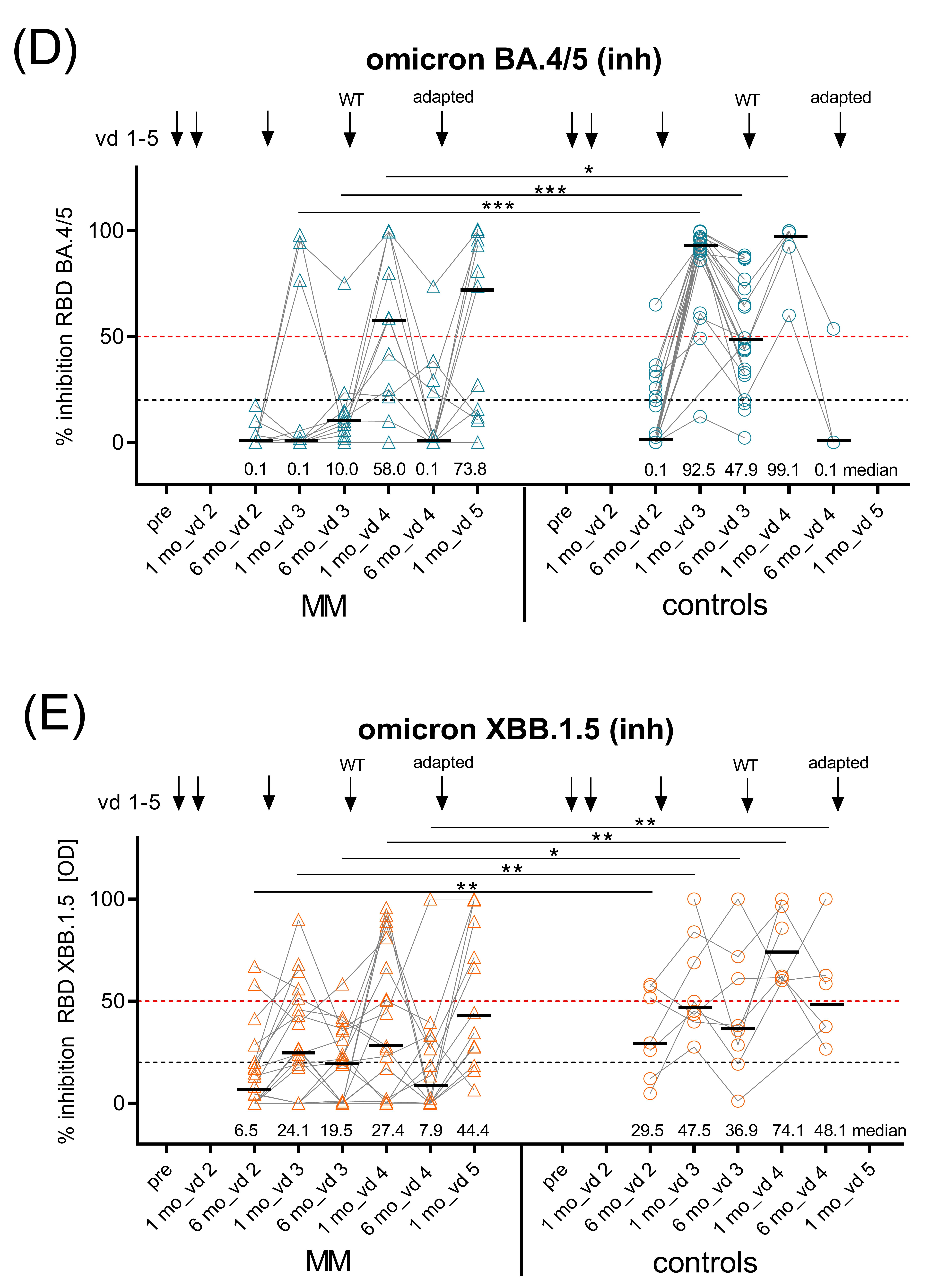
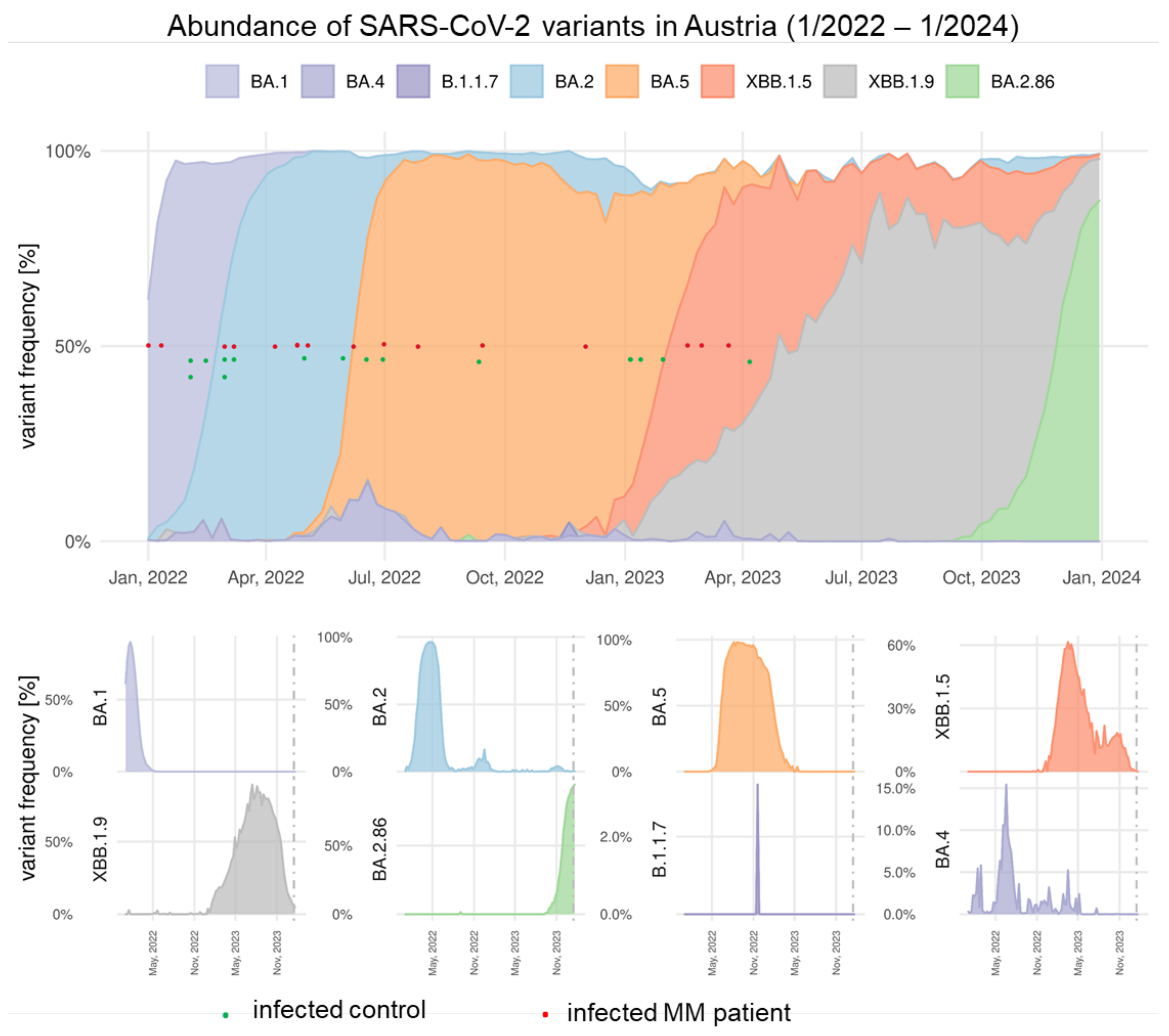
3.4. Breakthrough Infections Increase Ab Levels and Omicron-Cross Protection in Vaccinated MM Patients
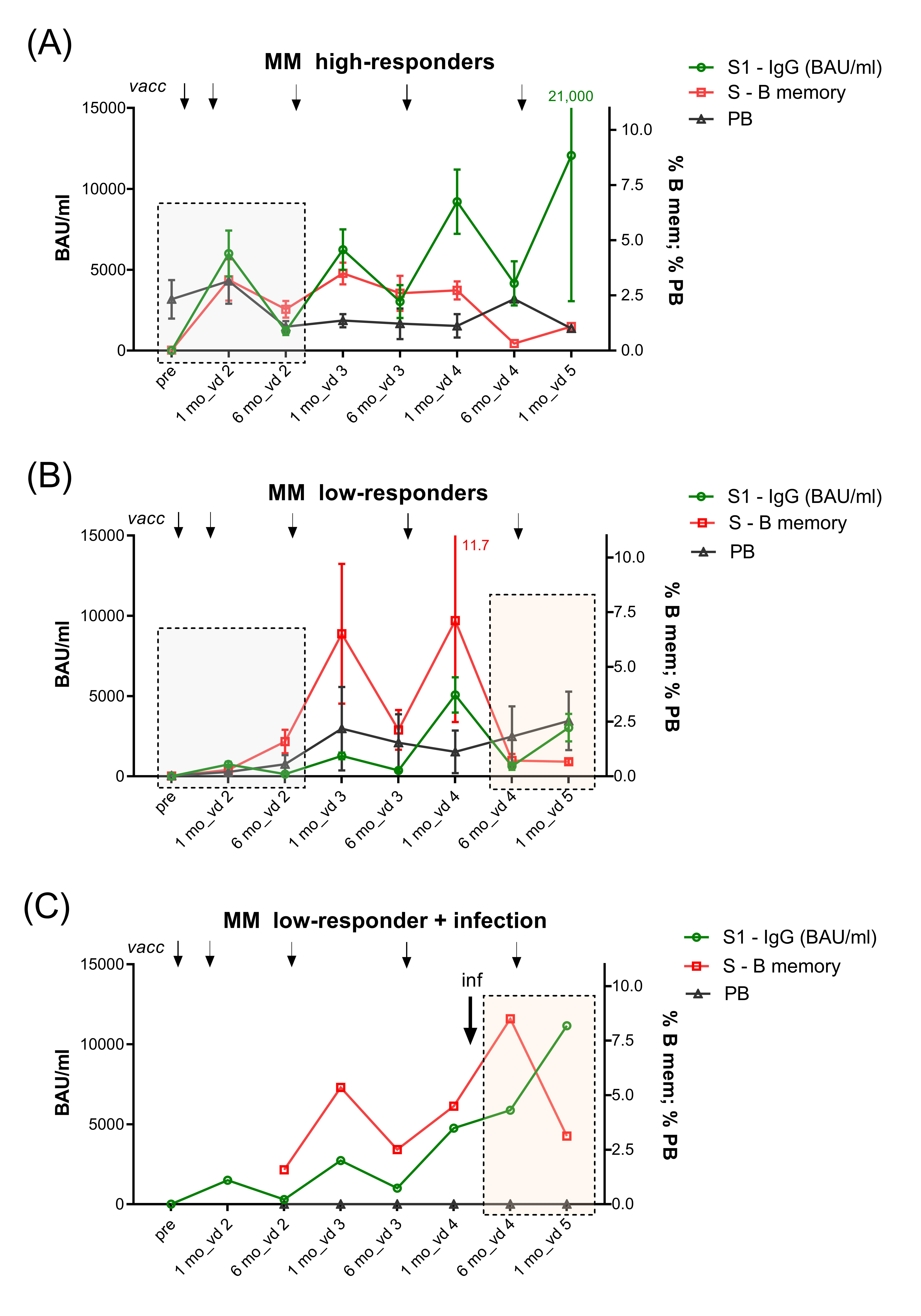
3.5. Lymphocyte Distributions and SARS-CoV-2 S-Protein-Specific Memory B Cells in MM Ab High and Low Responders
3.5.1. Leukocyte and Lymphocyte Counts
3.5.2. CD3+ T Cells, CD19+ B Cells, and NK T Cells
3.5.3. SARS-CoV-2 Spike (S) Protein-Specific Memory B Cells
3.6. SARS-CoV-2-Specific Cytokines Are Induced in MM Patients after Third and Fourth Vaccine Dose
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bird, S.A.; Boyd, K. Multiple myeloma: An overview of management. Palliat. Care Soc. Pract. 2019, 13, 1178224219868235. [Google Scholar] [CrossRef]
- Vijenthira, A.; Gong, I.Y.; Fox, T.A.; Booth, S.; Cook, G.; Fattizzo, B.; Martín-Moro, F.; Razanamahery, J.; Riches, J.C.; Zwicker, J.; et al. Outcomes of patients with hematologic malignancies and COVID-19: A systematic review and meta-analysis of 3377 patients. Blood 2020, 136, 2881–2892. [Google Scholar] [CrossRef] [PubMed]
- Becerril-Gaitan, A.; Vaca-Cartagena, B.F.; Ferrigno, A.S.; Mesa-Chavez, F.; Barrientos-Gutiérrez, T.; Tagliamento, M.; Lambertini, M.; Villarreal-Garza, C. Immunogenicity and risk of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) infection after Coronavirus Disease 2019 (COVID-19) vaccination in patients with cancer: A systematic review and meta-analysis. Eur. J. Cancer 2022, 160, 243–260. [Google Scholar] [CrossRef]
- Wagner, A.; Garner-Spitzer, E.; Schotta, A.M.; Orola, M.; Wessely, A.; Zwazl, I.; Ohradanova-Repic, A.; Weseslindtner, L.; Tajti, G.; Gebetsberger, L.; et al. SARS-CoV-2-mRNA Booster Vaccination Reverses Non-Responsiveness and Early Antibody Waning in Immunocompromised Patients-A Phase Four Study Comparing Immune Responses in Patients with Solid Cancers, Multiple Myeloma and Inflammatory Bowel Disease. Front. Immunol. 2022, 13, 889138. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Gavriatopoulou, M.; Ntanasis-Stathopoulos, I.; Briasoulis, A.; Gumeni, S.; Malandrakis, P.; Fotiou, D.; Papanagnou, E.D.; Migkou, M.; Theodorakakou, F.; et al. The neutralizing antibody response post COVID-19 vaccination in patients with myeloma is highly dependent on the type of anti-myeloma treatment. Blood Cancer J. 2021, 11, 138. [Google Scholar] [CrossRef] [PubMed]
- Van Oekelen, O.; Gleason, C.R.; Agte, S.; Srivastava, K.; Beach, K.F.; Aleman, A.; Kappes, K.; PVI/Seronet Team; Mouhieddine, T.H.; Wang, B.; et al. Highly variable SARS-CoV-2 spike antibody responses to two doses of COVID-19 RNA vaccination in patients with multiple myeloma. Cancer Cell 2021, 39, 1028–1030. [Google Scholar] [CrossRef]
- Stampfer, S.D.; Goldwater, M.S.; Jew, S.; Bujarski, S.; Regidor, B.; Daniely, D.; Chen, H.; Xu, N.; Li, M.; Green, T.; et al. Response to mRNA vaccination for COVID-19 among patients with multiple myeloma. Leukemia 2021, 35, 3534–3541. [Google Scholar] [CrossRef] [PubMed]
- Garner-Spitzer, E.; Wagner, A.; Gudipati, V.; Schoetta, A.M.; Orola-Taus, M.; Kundi, M.; Kunert, R.; Mayrhofer, P.; Huppa, J.B.; Stockinger, H.; et al. Lower magnitude and faster waning of antibody responses to SARS-CoV-2 vaccination in anti-TNF-alpha-treated IBD patients are linked to lack of activation and expansion of cTfh1 cells and impaired B memory cell formation. EBioMedicine 2023, 96, 104788. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, A.; Avet-Loiseau, H.; Oliva, S.; Lokhorst, H.M.; Goldschmidt, H.; Rosinol, L.; Richardson, P.; Caltagirone, S.; Lahuerta, J.J.; Facon, T.; et al. Revised International Staging System for Multiple Myeloma: A Report from International Myeloma Working Group. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2015, 33, 2863–2869. [Google Scholar] [CrossRef]
- Gattinger, P.; Tulaeva, I.; Borochova, K.; Kratzer, B.; Trapin, D.; Kropfmuller, A.; Pickl, W.F.; Valenta, R. Omicron: A SARS-CoV-2 variant of real concern. Allergy 2022, 77, 1616–1620. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef] [PubMed]
- Austrian Federal Ministry of Social Affairs, Health, Care and Consumer Protection. COVID-19-Impfungen, Ergänzung zum Impfplan Österreich 2022, Version 1.1, 28 November 2022. Available online: https://www.paediatrie.at/media/com_acymailing/upload/impfplan_2022.pdf (accessed on 23 April 2024).
- Amman, F.; Markt, R.; Endler, L.; Hupfauf, S.; Agerer, B.; Schedl, A.; Richter, L.; Zechmeister, M.; Bicher, M.; Heiler, G.; et al. Viral variant-resolved wastewater surveillance of SARS-CoV-2 at national scale. Nat. Biotechnol. 2022, 40, 1814–1822. [Google Scholar] [CrossRef] [PubMed]
- Hofsink, Q.; Haggenburg, S.; Lissenberg-Witte, B.I.; Broers, A.E.C.; van Doesum, J.A.; van Binnendijk, R.S.; den Hartog, G.; Bhoekhan, M.S.; Haverkate, N.J.E.; van Meerloo, J.; et al. Fourth mRNA COVID-19 vaccination in immunocompromised patients with haematological malignancies (COBRA KAI): A cohort study. EClinicalMedicine 2023, 61, 102040. [Google Scholar] [CrossRef]
- Azeem, M.I.; Nooka, A.K.; Shanmugasundaram, U.; Cheedarla, N.; Potdar, S.; Manalo, R.J.; Moreno, A.; Switchenko, J.M.; Cheedarla, S.; Doxie, D.B.; et al. Impaired SARS-CoV-2 Variant Neutralization and CD8+ T-cell Responses Following 3 Doses of mRNA Vaccines in Myeloma: Correlation with Breakthrough Infections. Blood Cancer Discov. 2023, 4, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Enssle, J.C.; Campe, J.; Buchel, S.; Moter, A.; See, F.; Griessbaum, K.; Rieger, M.A.; Wolf, S.; Ballo, O.; Steffen, B.; et al. Enhanced but variant-dependent serological and cellular immune responses to third-dose BNT162b2 vaccination in patients with multiple myeloma. Cancer Cell 2022, 40, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Aleman, A.; van Kesteren, M.; Zajdman, A.K.; Srivastava, K.; Cognigni, C.; Mischka, J.; Chen, L.Y.; Upadhyaya, B.; Serebryakova, K.; Nardulli, J.R.; et al. Cellular mechanisms associated with sub-optimal immune responses to SARS-CoV-2 bivalent booster vaccination in patients with Multiple Myeloma. EBioMedicine 2023, 98, 104886. [Google Scholar] [CrossRef] [PubMed]
- Van de Donk, N.; Usmani, S.Z. CD38 Antibodies in Multiple Myeloma: Mechanisms of Action and Modes of Resistance. Front. Immunol. 2018, 9, 2134. [Google Scholar] [CrossRef] [PubMed]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; Verbist, B.; Bald, J.; Plesner, T.; Syed, K.; Liu, K.; van de Donk, N.W.; Weiss, B.M.; et al. Daratumumab depletes CD38+ immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple myeloma. Blood 2016, 128, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Casneuf, T.; Xu, X.S.; Adams, H.C., 3rd; Axel, A.E.; Chiu, C.; Khan, I.; Ahmadi, T.; Yan, X.; Lonial, S.; Plesner, T.; et al. Effects of daratumumab on natural killer cells and impact on clinical outcomes in relapsed or refractory multiple myeloma. Blood Adv. 2017, 1, 2105–2114. [Google Scholar] [CrossRef]
- Verhoeven, D.; Grinwis, L.; Marsman, C.; Jansen, M.H.; Consortium, T.B.; Van Leeuwen, E.M.; Kuijpers, T.W. B-cell targeting with anti-CD38 daratumumab: Implications for differentiation and memory responses. Life Sci. Alliance 2023, 6. [Google Scholar] [CrossRef]
- Rooney, A.; Bivona, C.; Liu, B.; Streeter, D.; Gong, H.; Khan, Q. Risk of SARS-CoV-2 Breakthrough Infection in Vaccinated Cancer Patients: A Retrospective Cohort Study. J. Hematol. Oncol. 2022, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Goldblatt, M.J.; Casado Sastre, C.M.; Briggs, S.; Lindner, R. Isolation, Loneliness and Aloneness in the Age of COVID-19: Reflections on Clinical Experiences. Br. J. Psychother. 2022, 38, 738–753. [Google Scholar] [CrossRef] [PubMed]
- Lasrado, N.; Barouch, D.H. SARS-CoV-2 Hybrid Immunity: The Best of Both Worlds. J. Infect. Dis. 2023, 228, 1311–1313. [Google Scholar] [CrossRef]
- Bobrovitz, N.; Ware, H.; Ma, X.; Li, Z.; Hosseini, R.; Cao, C.; Selemon, A.; Whelan, M.; Premji, Z.; Issa, H.; et al. Protective effectiveness of previous SARS-CoV-2 infection and hybrid immunity against the omicron variant and severe disease: A systematic review and meta-regression. Lancet. Infect. Dis. 2023, 23, 556–567. [Google Scholar] [CrossRef]
- Enssle, J.C.; Campe, J.; Schwenger, A.; Wiercinska, E.; Hellstern, H.; Durrwald, R.; Rieger, M.A.; Wolf, S.; Ballo, O.; Steffen, B.; et al. Severe impairment of T-cell responses to BNT162b2 immunization in patients with multiple myeloma. Blood 2022, 139, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Bange, E.M.; Han, N.A.; Wileyto, P.; Kim, J.Y.; Gouma, S.; Robinson, J.; Greenplate, A.R.; Hwee, M.A.; Porterfield, F.; Owoyemi, O.; et al. CD8(+) T cells contribute to survival in patients with COVID-19 and hematologic cancer. Nat. Biotechnol. 2021, 27, 1280–1289. [Google Scholar] [CrossRef]
- Keppler-Hafkemeyer, A.; Greil, C.; Wratil, P.R.; Shoumariyeh, K.; Stern, M.; Hafkemeyer, A.; Ashok, D.; Hollaus, A.; Lupoli, G.; Priller, A.; et al. Potent high-avidity neutralizing antibodies and T cell responses after COVID-19 vaccination in individuals with B cell lymphoma and multiple myeloma. Nat. Cancer 2023, 4, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Enssle, J.C.; Campe, J.; Moter, A.; Voit, I.; Gessner, A.; Yu, W.; Wolf, S.; Steffen, B.; Serve, H.; Bremm, M.; et al. Cytokine-responsive T- and NK-cells portray SARS-CoV-2 vaccine-responders and infection in multiple myeloma patients. Leukemia 2024, 38, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.R.; Gao, Y.; Wu, J.; Ribeiro, O.; Chen, P.; Bergman, P.; Blennow, O.; Hansson, L.; Mielke, S.; Nowak, P.; et al. Memory T cells effectively recognize the SARS-CoV-2 hypermutated BA.2.86 variant. Cell Host Microbe 2024, 32, 156–161.e153. [Google Scholar] [CrossRef] [PubMed]
- Sigal, A. Milder disease with Omicron: Is it the virus or the pre-existing immunity? Nat. Rev. Immunol. 2022, 22, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Mieth, K.; Bennett, C.; Miller, C.; Anderson, L.D., Jr.; Chen, M.; Cao, J. Clinical Features and Risk Stratification of Multiple Myeloma Patients with COVID-19. Cancers 2023, 15, 3598. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Musto, P.; Engelhardt, M.; Delforge, M.; Cook, G.; Gay, F.; Van de Donk, N.W.; Ntanasis-Stathopoulos, I.; Vangsted, A.J.; Driessen, C.; et al. Management of patients with multiple myeloma and COVID-19 in the post pandemic era: A consensus paper from the European Myeloma Network (EMN). Leukemia 2023, 37, 1175–1185. [Google Scholar] [CrossRef]
- Wagner, A.; Garner-Spitzer, E.; Jasinska, J.; Kollaritsch, H.; Stiasny, K.; Kundi, M.; Wiedermann, U. Age-related differences in humoral and cellular immune responses after primary immunisation: Indications for stratified vaccination schedules. Sci. Rep. 2018, 8, 9825. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.P.; Liu, S.L. Challenges and Prospects in Developing Future SARS-CoV-2 Vaccines: Overcoming Original Antigenic Sin and Inducing Broadly Neutralizing Antibodies. J. Immunol. 2023, 211, 1459–1467. [Google Scholar] [CrossRef] [PubMed]
- Gattinger, P.; Kratzer, B.; Sehgal, A.; Ohradanova-Repic, A.; Gebetsberger, L.; Tajti, G.; Focke-Tejkl, M.; Schaar, M.; Fuhrmann, V.; Petrowitsch, L.; et al. Vaccine based on recombinant fusion protein combining HBV PreS with SARS-CoV-2 wild-type- and Omicron-derived RBD strongly induces Omicron-neutralizing antibodies in a murine model. Vaccines 2024, 24, 12. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Value | MM (n = 47) | Controls (n = 38) | |
|---|---|---|---|
| Gender (female), n (%) | 0.668 | 20 (43%) | 18 (47%) |
| Age (years), mean (SD) | <0.001 | 65.2 (9.1) | 54.8 (13.8) |
| Age < 60 years, n (%) | 12 (25.5) | 21 (55.2) | |
| BMI (kg/m2), mean (SD) | 0.599 | 25.9 (4.3) | 25.4 (4.4) |
| BMI > 30, n (%) | 0.601 | 12 (26%) | 7 (18%) |
| Time since diagnosis (years), mean (SD) | 7.6 (6) | n. a. | |
| MGUS | SCT w/o IT | SCT with IT * | MM Progressed | No SCT, IT or Other Therapy | Total n | |
|---|---|---|---|---|---|---|
| Group number | G 1 | G 2 | G 3 | G 4 | G 5 | |
| n per group | 7 | 11 | 12 | 12 | 4 | 46 |
| SCT | 0 | 11 | 12 | 10 | 0 | 33 |
| >2 y since SCT at inclusion | 9 | 8 | 6 | 23 | ||
| >2 y since SCT at 6 mo after vd3 | 10 | 10 | 10 | 30 | ||
| Treatment in G4 (MM Progressed) | No SCT + dara mono | SCT + dara or + dara & dexa | SCT + dara (of double or triple) | No SCT + PI | SCT + PI | |
| n per group | 1 | 5 | 4 | 1 | 1 | 12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, A.; Garner-Spitzer, E.; Auer, C.; Gattinger, P.; Zwazl, I.; Platzer, R.; Orola-Taus, M.; Pichler, P.; Amman, F.; Bergthaler, A.; et al. Breakthrough Infections in SARS-CoV-2-Vaccinated Multiple Myeloma Patients Improve Cross-Protection against Omicron Variants. Vaccines 2024, 12, 518. https://doi.org/10.3390/vaccines12050518
Wagner A, Garner-Spitzer E, Auer C, Gattinger P, Zwazl I, Platzer R, Orola-Taus M, Pichler P, Amman F, Bergthaler A, et al. Breakthrough Infections in SARS-CoV-2-Vaccinated Multiple Myeloma Patients Improve Cross-Protection against Omicron Variants. Vaccines. 2024; 12(5):518. https://doi.org/10.3390/vaccines12050518
Chicago/Turabian StyleWagner, Angelika, Erika Garner-Spitzer, Claudia Auer, Pia Gattinger, Ines Zwazl, René Platzer, Maria Orola-Taus, Peter Pichler, Fabian Amman, Andreas Bergthaler, and et al. 2024. "Breakthrough Infections in SARS-CoV-2-Vaccinated Multiple Myeloma Patients Improve Cross-Protection against Omicron Variants" Vaccines 12, no. 5: 518. https://doi.org/10.3390/vaccines12050518