
Astrogliosis in the GFAP-CreERT2:Rosa26iDTR Mouse Model Does Not Exacerbate Retinal Microglia Activation or Müller Cell Gliosis under Hypoxic Conditions

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Diphtheria Toxin-Mediated Astrocyte Activation in the GFAP-CreERT2:Rosa26iDTR Mouse
2.3. Induction of Hypoxia-Induced Retinopathy
2.4. Tissue Collection
2.5. Immunofluorescent Staining
2.6. Tissue Imaging and Quantification
2.7. Protein Quantification
2.8. Statistical Analysis
3. Results
3.1. Diphtheria Toxin Administration in GFAP-CreERT2:Rosa26iDTR Mice Is Associated with Reactive Astrogliosis and Vimentin Overexpression
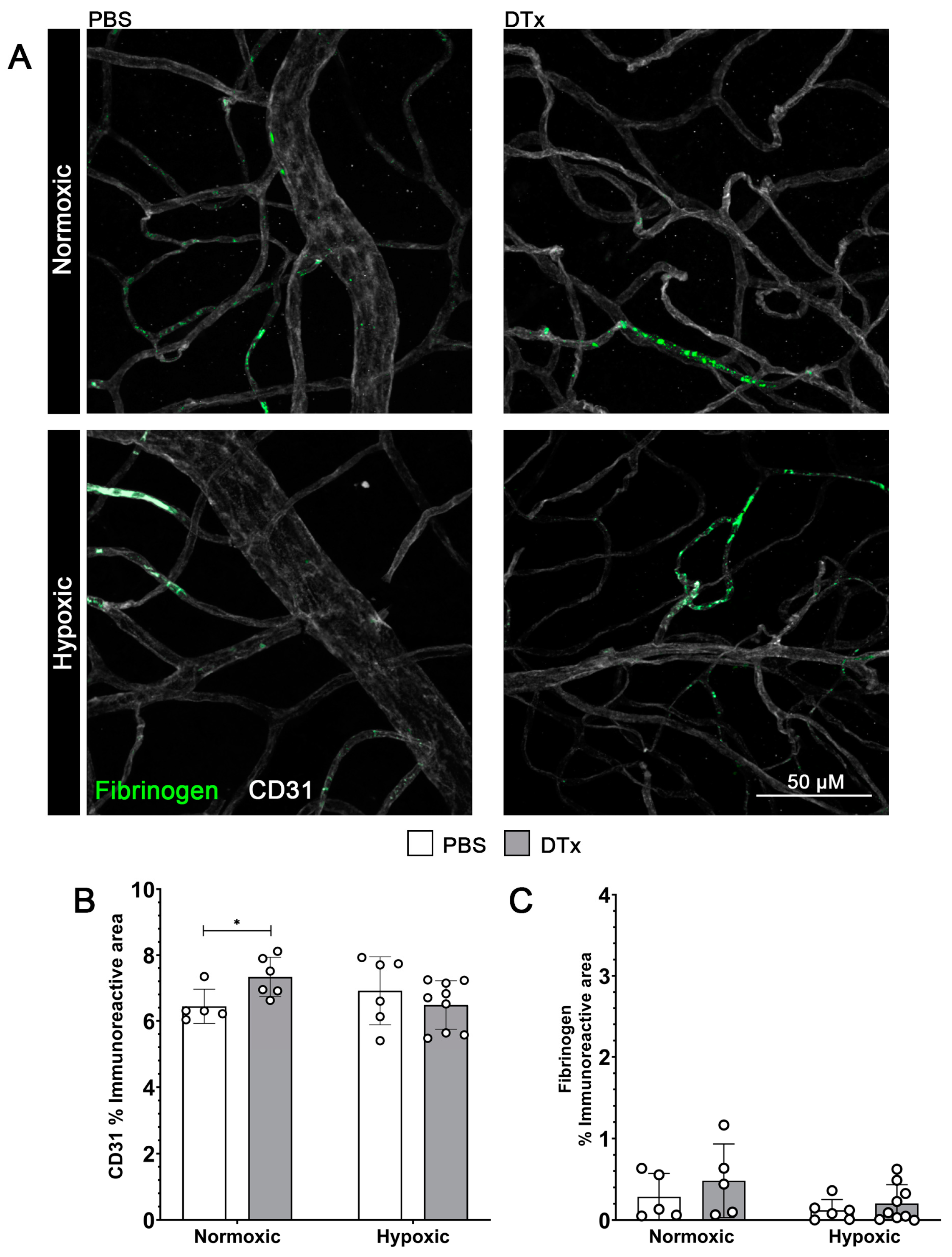
3.2. Reactive Astrogliosis in the GFAP-CreERT2:Rosa26iDTR Mouse Does Not Affect Vascular Integrity
3.3. Hypoxia Does Not Alter Neuronal Densities in the GFAP-CreERT2:Rosa26iDTR Mouse Model
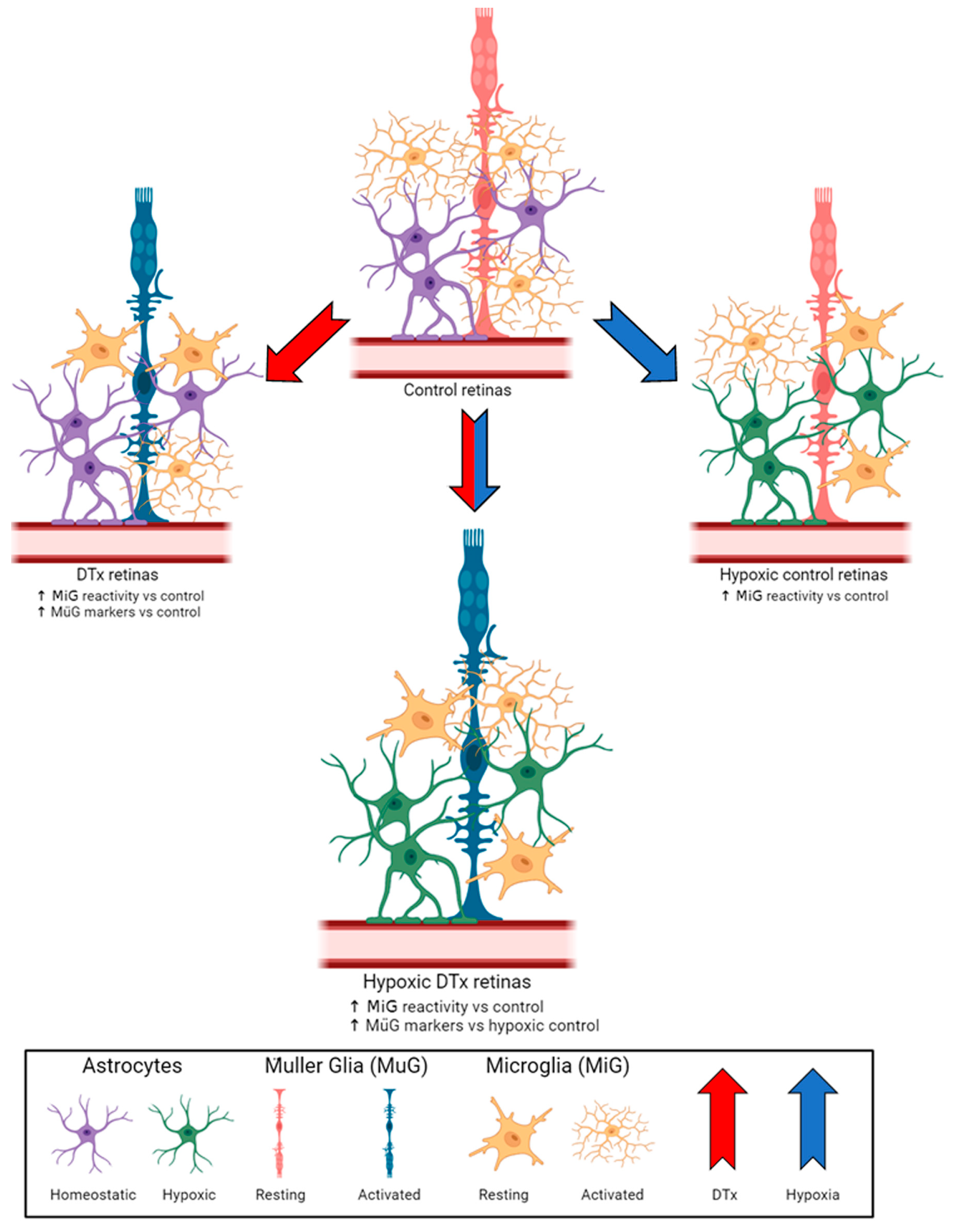
3.4. Retinal Microglial Cells in DTx-Treated Mice Show Evidence of Morphological Activation Independent of Hypoxic Conditions
3.5. DTx-Induced Retinal Pathology Is Not Explained by the Expression of Standard Inflammatory and Anti-Inflammatory Mediators
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sapkota, A.; Halder, S.K.; Milner, R. Hypoxia-induced vascular remodeling responses in the brain are much more robust than other organs. Microvasc. Res. 2023, 148, 104517. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Tian, M.; Yang, G.; Tan, Q.; Chen, Y.; Li, G.; Zhang, Q.; Li, Y.; Wan, P.; Wu, J. Hypoxia signaling in human health and diseases: Implications and prospects for therapeutics. Signal Transduct. Target. Ther. 2022, 7, 218. [Google Scholar] [CrossRef]
- Shinojima, A.; Lee, D.; Tsubota, K.; Negishi, K.; Kurihara, T. Retinal Diseases Regulated by Hypoxia-Basic and Clinical Perspectives: A Comprehensive Review. J. Clin. Med. 2021, 10, 5496. [Google Scholar] [CrossRef] [PubMed]
- Atef, M.M.; Shafik, N.M.; Hafez, Y.M.; Watany, M.M.; Selim, A.; Shafik, H.M.; El-Deeb, O.S. The evolving role of long noncoding RNA HIF1A-AS2 in diabetic retinopathy: A cross-link axis between hypoxia, oxidative stress and angiogenesis via MAPK/VEGF-dependent pathway. Redox Rep. 2022, 27, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Majidova, S.R. Evaluation of Hypoxia and Microcirculation Factors in the Progression of Diabetic Retinopathy. Investig. Opthalmology Vis. Sci. 2024, 65, 35. [Google Scholar] [CrossRef]
- Janáky, M.; Grósz, A.; Tóth, E.; Benedek, K.; Benedek, G. Hypobaric hypoxia reduces the amplitude of oscillatory potentials in the human ERG. Doc. Ophthalmol. 2007, 114, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kinuthia, U.M.; Wolf, A.; Langmann, T. Microglia and Inflammatory Responses in Diabetic Retinopathy. Front. Immunol. 2020, 11, 564077. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-Y.; Tang, S.-C.; Lee, J.-E.; Chen, Y.-R.; Chen, Y.-T.; Chen, K.-W.; Hsieh, S.-T.; Wang, K.-C. Recombinant soluble form of receptor for advanced glycation end products ameliorates microcirculation impairment and neuroinflammation after subarachnoid hemorrhage. Neurotherapeutics 2024, 21, e00312. [Google Scholar] [CrossRef] [PubMed]
- Butturini, E.; Boriero, D.; de Prati, A.C.; Mariotto, S. STAT1 drives M1 microglia activation and neuroinflammation under hypoxia. Arch. Biochem. Biophys. 2019, 669, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Huang, W.; Zhang, Z.; Yu, L.; Xie, C.; Zhu, D.; Peng, Z.; Chen, J. Hypoxia-inducible factor-1α upregulation in microglia following hypoxia protects against ischemia-induced cerebral infarction. Neuroreport 2014, 25, 1122–1128. [Google Scholar] [CrossRef]
- Halder, S.K.; Milner, R. The impact of chronic mild hypoxia on cerebrovascular remodelling; uncoupling of angiogenesis and vascular breakdown. Fluids Barriers CNS 2021, 18, 50. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, A.S.; Yan, Z.; Dixit, K.; Johnson, J.R.; Bouhaddou, M.; Meyer-Franke, A.; Shin, M.-G.; Yong, Y.; Agrawal, A.; MacDonald, E.; et al. Defining blood-induced microglia functions in neurodegeneration through multiomic profiling. Nat. Immunol. 2023, 24, 1173–1187. [Google Scholar] [CrossRef] [PubMed]
- Neal, M.; Luo, J.; Harischandra, D.S.; Gordon, R.; Sarkar, S.; Jin, H.; Anantharam, V.; Désaubry, L.; Kanthasamy, A.; Kanthasamy, A. Prokineticin-2 promotes chemotaxis and alternative A2 reactivity of astrocytes. Glia 2018, 66, 2137–2157. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.Y.; Lee, A.G.; Culbertson, C.; Sun, G.; Talati, R.K.; Manley, N.C.; Li, X.; Zhao, H.; Lyons, D.M.; Zhou, Q.-Y.; et al. Prokineticin 2 is an endangering mediator of cerebral ischemic injury. Proc. Natl. Acad. Sci. USA 2012, 109, 5475–5480. [Google Scholar] [CrossRef] [PubMed]
- Shukla, A.; Bhardwaj, U.; Apoorva; Seth, P.; Singh, S.K. Hypoxia-Induced miR-101 Impairs Endothelial Barrier Integrity Through Altering VE-Cadherin and Claudin-5. Mol. Neurobiol. 2024, 61, 1807–1817. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.K.; Milner, R. Mild hypoxia triggers transient blood-brain barrier disruption: A fundamental protective role for microglia. Acta Neuropathol. Commun. 2020, 8, 175. [Google Scholar] [CrossRef] [PubMed]
- Forrester, J.V.; Kuffova, L.; Delibegovic, M. The Role of Inflammation in Diabetic Retinopathy. Front. Immunol. 2020, 11, 583687. [Google Scholar] [CrossRef]
- Cardona, A.E.; Pioro, E.P.; Sasse, M.E.; Kostenko, V.; Cardona, S.M.; Dijkstra, I.M.; Huang, D.; Kidd, G.; Dombrowski, S.; Dutta, R.; et al. Control of microglial neurotoxicity by the fractalkine receptor. Nat. Neurosci. 2006, 9, 917–924. [Google Scholar] [CrossRef]
- Cardona, S.M.; Mendiola, A.S.; Yang, Y.-C.; Adkins, S.L.; Torres, V.; Cardona, A.E. Disruption of Fractalkine Signaling Leads to Microglial Activation and Neuronal Damage in the Diabetic Retina. ASN Neuro 2015, 7, 1759091415608204. [Google Scholar] [CrossRef]
- Church, K.A.; Rodriguez, D.; Mendiola, A.S.; Vanegas, D.; Gutierrez, I.L.; Tamayo, I.; Amadu, A.; Velazquez, P.; Cardona, S.M.; Gyoneva, S.; et al. Pharmacological depletion of microglia alleviates neuronal and vascular damage in the diabetic CX3CR1-WT retina but not in CX3CR1-KO or hCX3CR1(I249/M280)-expressing retina. Front. Immunol. 2023, 14, 1130735. [Google Scholar] [CrossRef]
- Church, K.A.; Rodriguez, D.; Vanegas, D.; Gutierrez, I.L.; Cardona, S.M.; Madrigal, J.L.M.; Kaur, T.; Cardona, A.E. Models of microglia depletion and replenishment elicit protective effects to alleviate vascular and neuronal damage in the diabetic murine retina. J. Neuroinflamm. 2022, 19, 300. [Google Scholar] [CrossRef]
- Rodriguez, D.; Church, K.A.; Smith, C.T.; Vanegas, D.; Cardona, S.M.; Muzzio, I.A.; Nash, K.R.; Cardona, A.E. Therapeutic Delivery of Soluble Fractalkine Ameliorates Vascular Dysfunction in the Diabetic Retina. Int. J. Mol. Sci. 2024, 25, 1727. [Google Scholar] [CrossRef]
- Pawelec, P.; Ziemka-Nalecz, M.; Sypecka, J.; Zalewska, T. The Impact of the CX3CL1/CX3CR1 Axis in Neurological Disorders. Cells 2020, 9, 2277. [Google Scholar] [CrossRef] [PubMed]
- Buch, T.; Heppner, F.L.; Tertilt, C.; Heinen, T.J.A.J.; Kremer, M.; Wunderlich, F.T.; Jung, S.; Waisman, A. A Cre-inducible diphtheria toxin receptor mediates cell lineage ablation after toxin administration. Nat. Methods 2005, 2, 419–426. [Google Scholar] [CrossRef]
- Chun, H.; Im, H.; Kang, Y.J.; Kim, Y.; Shin, J.H.; Won, W.; Lim, J.; Ju, Y.; Park, Y.M.; Kim, S.; et al. Severe reactive astrocytes precipitate pathological hallmarks of Alzheimer's disease via H2O2− production. Nat. Neurosci. 2020, 23, 1555–1566. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Tanaka, J.; Toku, K.; Tateishi, N.; Suzuki, Y.; Matsuda, S.; Sakanaka, M.; Maeda, N. Effects of GM-CSF and ordinary supplements on the ramification of microglia in culture: A morphometrical study. Glia 1996, 18, 269–281. [Google Scholar] [CrossRef]
- Sethi, C.S.; Lewis, G.P.; Fisher, S.K.; Leitner, W.P.; Mann, D.L.; Luthert, P.J.; Charteris, D.G. Glial remodeling and neural plasticity in human retinal detachment with proliferative vitreoretinopathy. Investig. Ophthalmol. Vis. Sci. 2005, 46, 329–342. [Google Scholar] [CrossRef]
- Kugler, E.C.; Greenwood, J.; MacDonald, R.B. The "Neuro-Glial-Vascular" Unit: The Role of Glia in Neurovascular Unit Formation and Dysfunction. Front. Cell Dev. Biol. 2021, 9, 732820. [Google Scholar] [CrossRef]
- Amor, S.; Puentes, F.; Baker, D.; Van Der Valk, P. Inflammation in neurodegenerative diseases. Immunology 2010, 129, 154–169. [Google Scholar] [CrossRef]
- Veje, M.; Griška, V.; Pakalnienė, J.; Mickienė, A.; Bremell, D.; Zetterberg, H.; Blennow, K.; Lindquist, L.; Studahl, M. Serum and cerebrospinal fluid brain damage markers neurofilament light and glial fibrillary acidic protein correlate with tick-borne encephalitis disease severity-a multicentre study on Lithuanian and Swedish patients. Eur. J. Neurol. 2023, 30, 3182–3189. [Google Scholar] [CrossRef]
- Tomita, Y.; Qiu, C.; Bull, E.; Allen, W.; Kotoda, Y.; Talukdar, S.; Smith, L.E.H.; Fu, Z. Müller glial responses compensate for degenerating photoreceptors in retinitis pigmentosa. Exp. Mol. Med. 2021, 53, 1748–1758. [Google Scholar] [CrossRef] [PubMed]
- Newman, E.A. Propagation of intercellular calcium waves in retinal astrocytes and Müller cells. J. Neurosci. 2001, 21, 2215–2223. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.E.; Wesolowski, E.; McLellan, A.; Kostyk, S.K.; D'amato, R.; Sullivan, R.; D'Amore, P.A. Oxygen-induced retinopathy in the mouse. Investig. Ophthalmol. Vis. Sci. 1994, 35, 101–111. [Google Scholar]
- I Dorrell, M.; Aguilar, E.; Friedlander, M. Retinal vascular development is mediated by endothelial filopodia, a preexisting astrocytic template and specific R-cadherin adhesion. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3500–3510. [Google Scholar]
- Ferreira, T.A.; Blackman, A.V.; Oyrer, J.; Jayabal, S.; Chung, A.J.; Watt, A.J.; Sjöström, P.J.; Van Meyel, D.J. Neuronal morphometry directly from bitmap images. Nat. Methods 2014, 11, 982–984. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization From M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef] [PubMed]
- Imtiyaz, H.Z.; Williams, E.P.; Hickey, M.M.; Patel, S.A.; Durham, A.C.; Yuan, L.-J.; Hammond, R.; Gimotty, P.A.; Keith, B.; Simon, M.C. Hypoxia-inducible factor 2alpha regulates macrophage function in mouse models of acute and tumor inflammation. J. Clin. Investig. 2010, 120, 2699–2714. [Google Scholar] [CrossRef] [PubMed]
- Takeda, N.; O'Dea, E.L.; Doedens, A.; Kim, J.-W.; Weidemann, A.; Stockmann, C.; Asagiri, M.; Simon, M.C.; Hoffmann, A.; Johnson, R.S. Differential activation and antagonistic function of HIF-α isoforms in macrophages are essential for NO homeostasis. Genes Dev. 2010, 24, 491–501. [Google Scholar] [CrossRef]
- Ravenna, L.; Salvatori, L.; Russo, M.A. HIF3α: The little we know. FEBS J. 2016, 283, 993–1003. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, Y.; Xu, H.; Zhang, Z.; Wang, X.; Yuan, L.; Lenahan, C.; Zhang, C.; Jiang, J.; Fang, C.; et al. CCL17 exerts neuroprotection through activation of CCR4/mTORC2 axis in microglia after subarachnoid haemorrhage in rats. Stroke Vasc. Neurol. 2022, 8, 4–16. [Google Scholar] [CrossRef]
- Shi, Z.; Jiang, X.; Geng, Y.; Yue, X.; Gao, J.; Cheng, X.; Zhao, M.; Zhu, L. Expression profile of cytokines and chemokines in a mouse high-altitude cerebral edema model. Int. J. Immunopathol. Pharmacol. 2023, 37, 3946320231177189. [Google Scholar] [CrossRef]
- Shukla, U.V.; Tripathy, K. Diabetic Retinopathy. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary | ||||||
|---|---|---|---|---|---|---|
| Target/Cell | Host | RRID | Dilution | |||
| ionized calcium binding adaptor molecule-1 (Iba1)/Microglia | Rabbit | AB_839504 | 1:4000 | |||
| neuronal nuclei (NeuN)/RGC | Mouse | AB_2298772 | 1:4000 | |||
| glial fibrillary acidic protein (GFAP)/Astrocytes | Rat | AB_2532994 | 1:4000 | |||
| platelet endothelial cell adhesion molecule (PECAM-1/CD31)/Endothelial cells | Rat | AB_393571 | 1:500 | |||
| fibrinogen | Rabbit | AB_578481 | 1:2000 | |||
| Vimentin/Müller glia | Chicken | AB_2216267 | 1:1000 | |||
| S100β | Rabbit | AB_956280 | 1:1000 | |||
| Secondary | ||||||
| Target | Host | RRID | Dilution | |||
| anti-rabbit 488 | Donkey | AB_2313584 | 1:1000 | |||
| anti-rabbit Cy3 | Goat | AB_2338006 | 1:1000 | |||
| anti-rat Cy5 | Donkey | AB_2340694 | 1:1000 | |||
| anti-rat Cy3 | Goat | AB_2338394 | 1:1000 | |||
| anti-mouse Cy3 | Goat | AB_2338709 | 1:1000 | |||
| anti-chicken Biotin | Donkey | AB_2340363 | 1:1000 | |||
| Streptavidin Cy3 | N/A | AB_2337244 | 1:1000 | |||
| Protein | Values ± SD 1 | Two-Way ANOVA p Value | |||||
|---|---|---|---|---|---|---|---|
| PBS Nor | PBS Hyp | DTx Nor | DTx Hyp | PBS Nor vs. Hyp | DTx Nor vs. Hyp | ||
| IL-1β | Brain | 5.6 ± 1.67 | 5.18 ± 0.77 | 4.58 ± 1.22 | 5.04 ± 0.13 | 0.9289 | 0.9107 |
| Serum | 18.87 ± 3.67 | 19.44 ± 1.92 | 18.89 ± 4.47 | 20.42 ± 1.02 | 0.991 | 0.8604 | |
| IL-2 | Brain | 5.25 ± 1.76 | 4.67 ± 0.53 | 4.04 ± 1.31 | 4.71 ± 0.32 | 0.8532 | 0.7881 |
| Serum | 19.89 ± 9.09 | 15.39 ± 3.31 | 22.79 ± 16.46 | 16.21 ± 0.59 | 0.8777 | 0.7002 | |
| IL-6 | Brain | 4.28 ± 1.67 | 4.78 ± 0.9 | 3.57 ± 2.06 | 4.41 ± 0.88 | 0.9479 | 0.8027 |
| Serum | 46.42 ± 60.26 | 19.8 ± 8.19 | 20.52 ± 6.66 | 22.34 ± 4.35 | 0.5333 | 0.9997 | |
| IFN-γ | Brain | 5.72 ± 2.49 | 5.21 ± 0.7 | 3.86 ± 1.94 | 5.15 ± 0.25 | 0.9593 | 0.6021 |
| Serum | 53.25 ± 55 | 19.88 ± 11.8 | 29.05 ± 17.71 | 22.65 ± 3.09 | 0.3149 | 0.9856 | |
| TNF-α | Brain | 14.27 ± 6.94 | 13.17 ± 1.38 | 9.95 ± 4.64 | 13.15 ± 1.32 | 0.9762 | 0.647 |
| Serum | 87.91 ± 102.76 | 44.44 ± 17.32 | 63.75 ± 55.98 | 48.97 ± 9.74 | 0.6604 | 0.9786 | |
| IL-4 | Brain | 3.48 ± 2.22 | 3.05 ± 0.82 | 2.13 ± 1.16 | 3.23 ± 0.84 | 0.9599 | 0.6046 |
| Serum | 36.48 ± 30.1 | 13.59 ± 6.54 | 24.27 ± 14.83 | 16.08 ± 3.37 | 0.193 | 0.8739 | |
| IL-5 | Brain | 3.52 ± 1.38 | 2.92 ± 0.33 | 2.48 ± 1.01 | 2.97 ± 0.24 | 0.7045 | 0.8145 |
| Serum | 18.88 ± 4.84 | 16.1 ± 5.19 | 17.64 ± 5.76 | 16.66 ± 3.43 | 0.8048 | 0.9886 | |
| IL-10 | Brain | 13.3 ± 5.71 | 12.68 ± 1.62 | 10.1 ± 4 | 12.32 ± 0.57 | 0.9924 | 0.7648 |
| Serum | 86.63 ± 86.11 | 54.46 ± 24.69 | 49.28 ± 35.74 | 57.15 ± 11.76 | 0.7251 | 0.9939 | |
| CCL-17 | Brain | 2.71 ± 4.29 | 2.32 ± 2.08 | 2.51 ± 2.21 | 5.1 ± 3.35 | 0.9971 | 0.5689 |
| Serum | 42.31 ± 34.18 | 48.4 ± 30.63 | 53.75 ± 55.28 | 83.72 ± 24.76 | 0.9968 | 0.7165 | |
| GM-CSF | Brain | 13.1 ± 4.2 | 11.99 ± 1.63 | 10.33 ± 3.53 | 11.79 ± 0.61 | 0.9278 | 0.8535 |
| Serum | 43.33 ± 26.98 | 50.74 ± 6.91 | 58.37 ± 57.07 | 49.5 ± 7.46 | 0.9826 | 0.9709 | |
| IL-18 | Brain | 22.56 ± 16.66 | 40 ± 19.72 | 34.94 ± 19.27 | 36.12 ± 19.49 | 0.4801 | 0.9996 |
| Serum | 23.17 ± 26.86 | 16.82 ± 8.5 | 41.82 ± 50.3 | 16.98 ± 6.05 | 0.9915 | 0.6493 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rorex, C.; Cardona, S.M.; Church, K.A.; Rodriguez, D.; Vanegas, D.; Saldivar, R.; Faz, B.; Cardona, A.E. Astrogliosis in the GFAP-CreERT2:Rosa26iDTR Mouse Model Does Not Exacerbate Retinal Microglia Activation or Müller Cell Gliosis under Hypoxic Conditions. Biomolecules 2024, 14, 567. https://doi.org/10.3390/biom14050567
Rorex C, Cardona SM, Church KA, Rodriguez D, Vanegas D, Saldivar R, Faz B, Cardona AE. Astrogliosis in the GFAP-CreERT2:Rosa26iDTR Mouse Model Does Not Exacerbate Retinal Microglia Activation or Müller Cell Gliosis under Hypoxic Conditions. Biomolecules. 2024; 14(5):567. https://doi.org/10.3390/biom14050567
Chicago/Turabian StyleRorex, Colin, Sandra M. Cardona, Kaira A. Church, Derek Rodriguez, Difernando Vanegas, Reina Saldivar, Brianna Faz, and Astrid E. Cardona. 2024. "Astrogliosis in the GFAP-CreERT2:Rosa26iDTR Mouse Model Does Not Exacerbate Retinal Microglia Activation or Müller Cell Gliosis under Hypoxic Conditions" Biomolecules 14, no. 5: 567. https://doi.org/10.3390/biom14050567