
Effects of Wind Exposure and Deficit Irrigation on Vegetative Growth, Yield Components and Berry Composition of Malbec and Cabernet Sauvignon

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
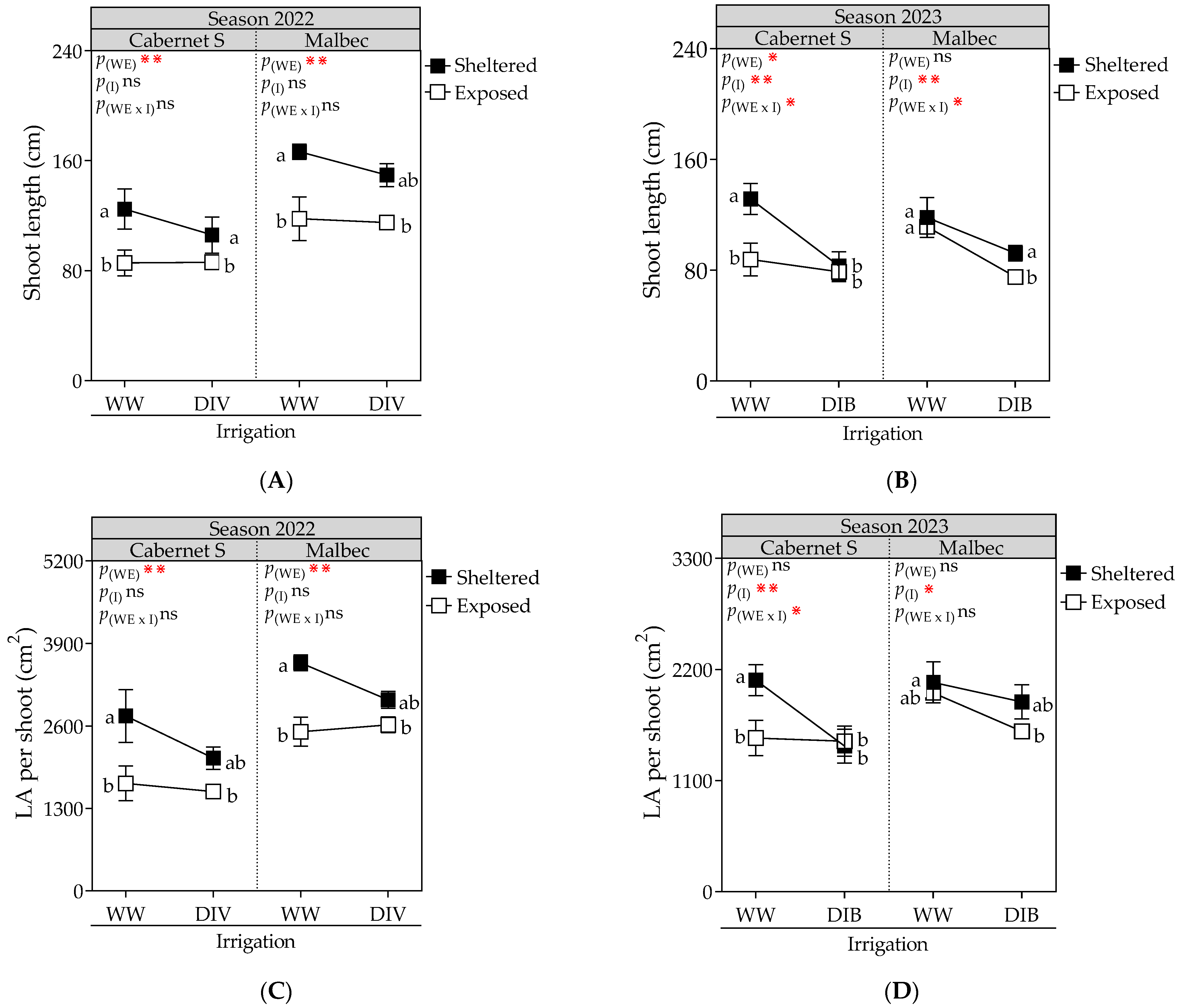
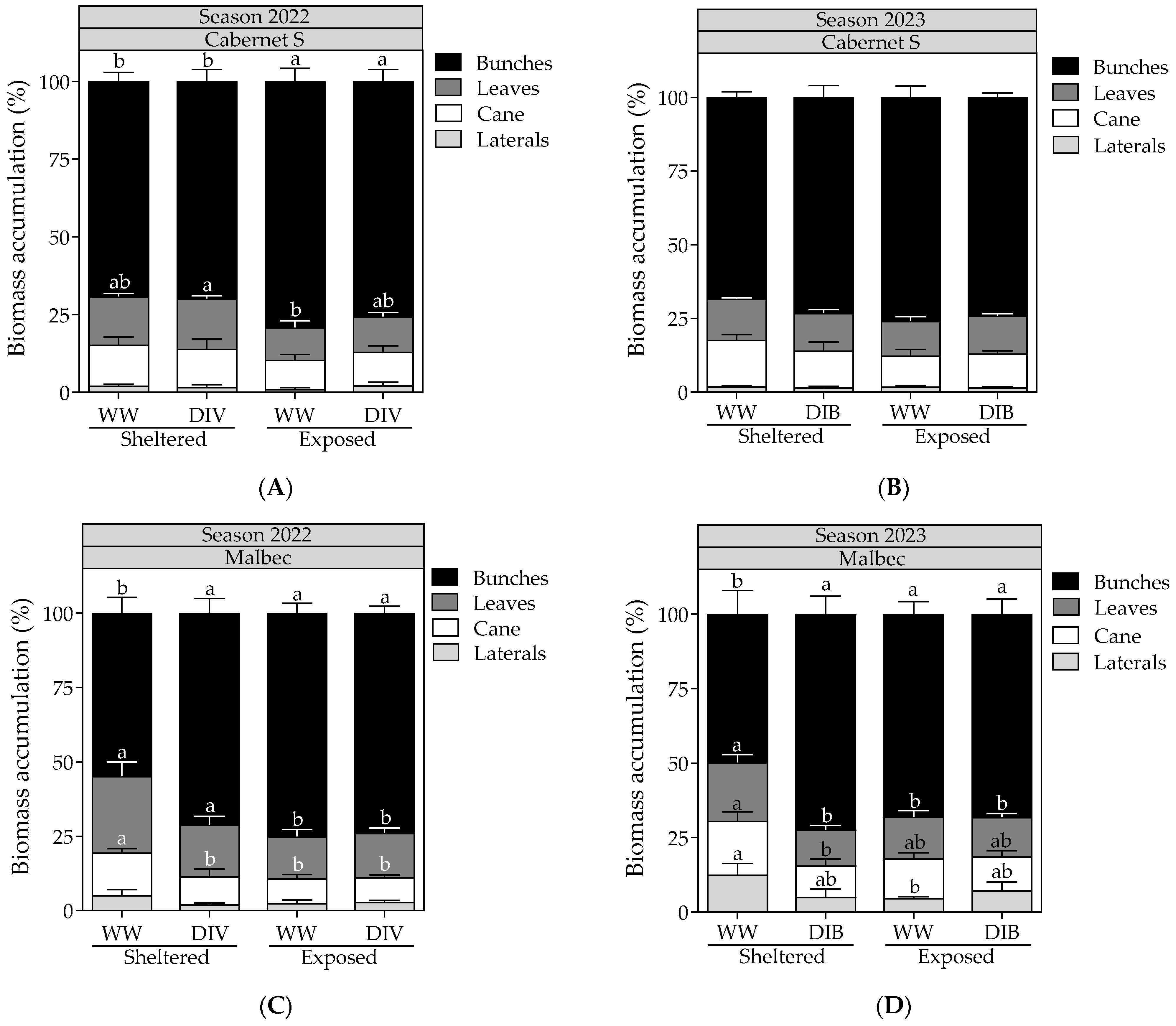
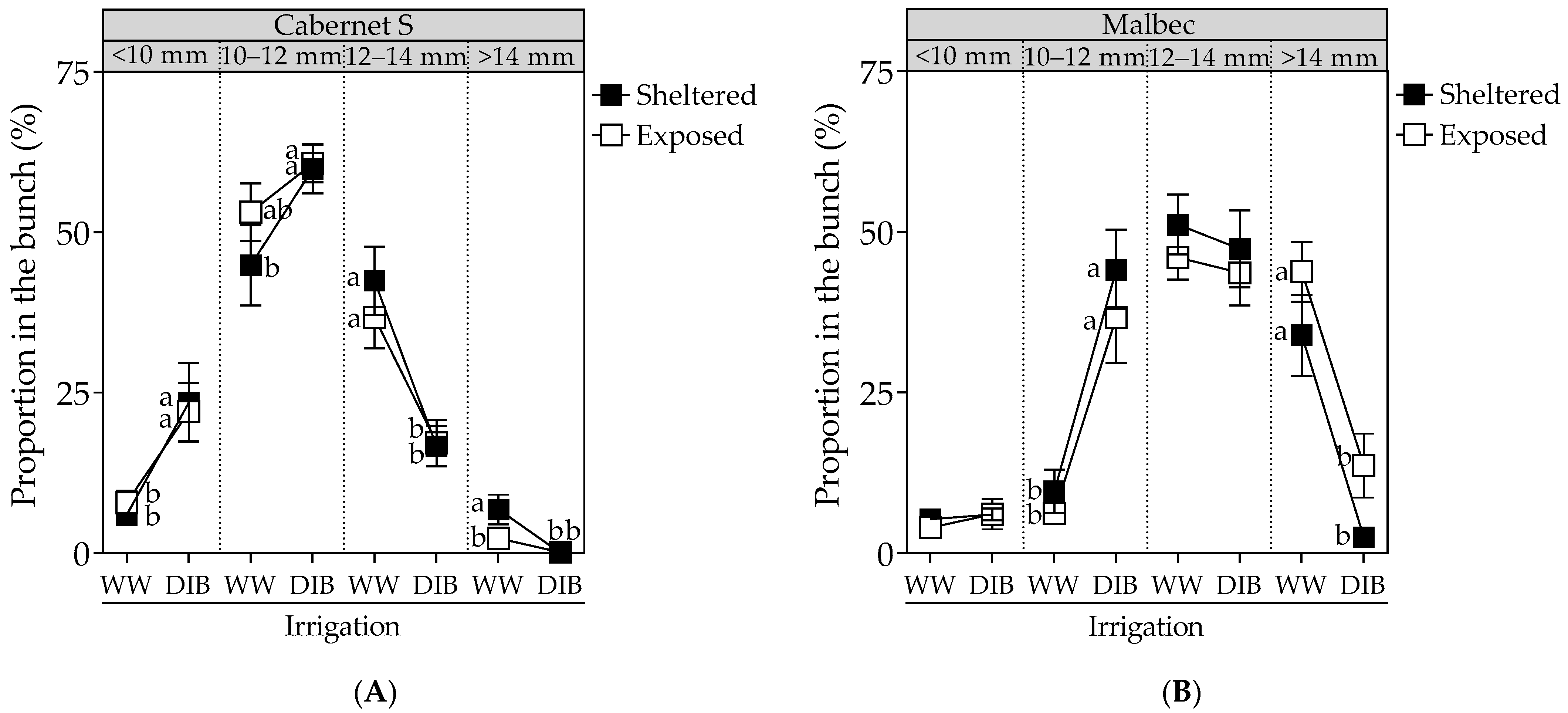
2.1. Vegetative Growth, Stomatal Conductance, Biomass Partitioning, Fruit Yield and Proportion of Berry Sizes
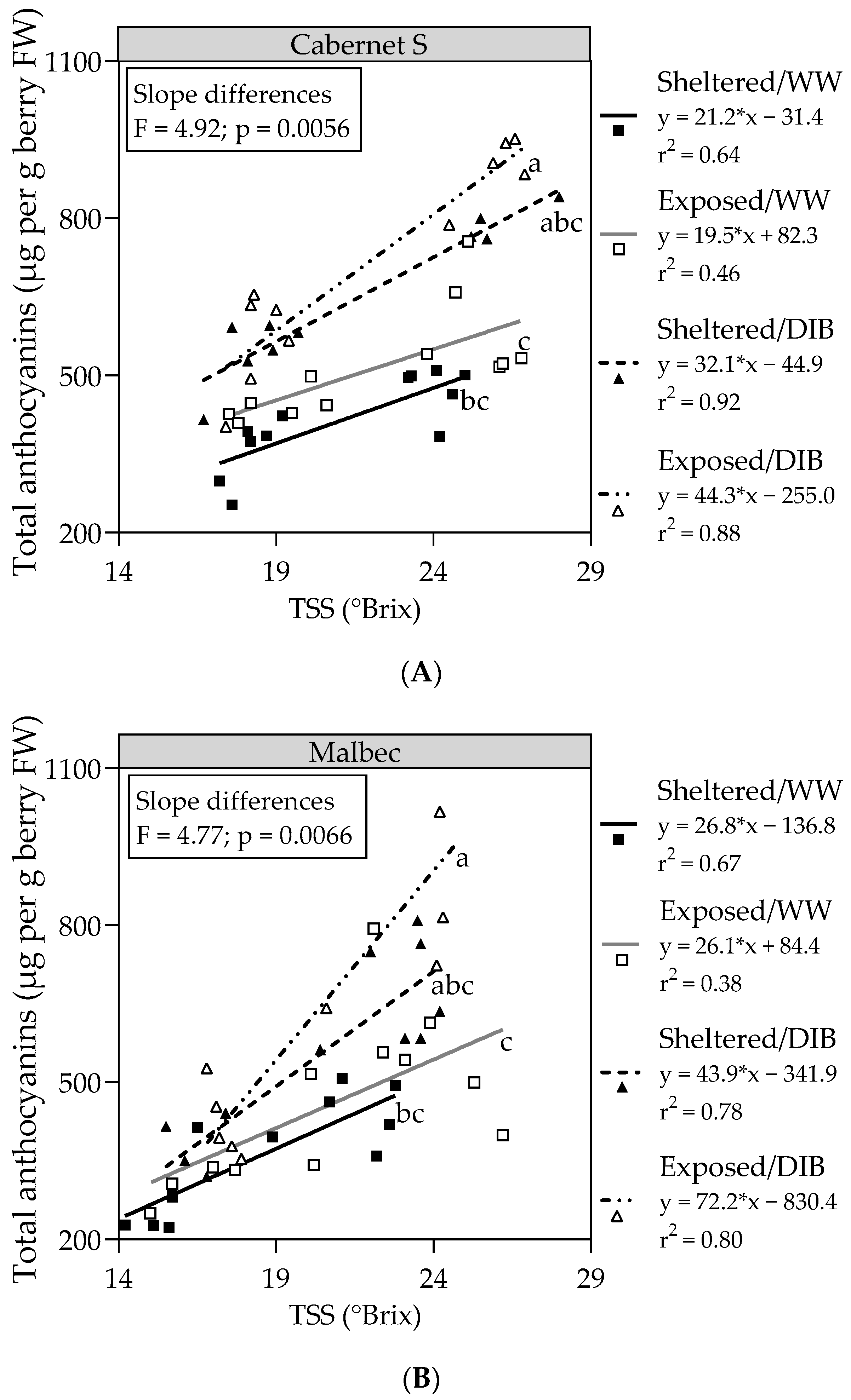
2.2. Berry Skin Total Phenolics and Relationship between Total Anthocyanins and Total Soluble Solids
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Design
4.2. Vegetative Growth and Physiological Parameters
4.3. Berry Sampling, Total Soluble Solids, Skin Phenolic Contents, Yield Component and Biomass Accumulation
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- OIV. Annual Assessment of the World Vine and Wine Sector in 2022; International Organisation of Vine and Wine (OIV): Dijon, France, 2022. [Google Scholar]
- Cabré, F.; Nuñez, M. Impacts of climate change on viticulture in Argentina. Reg. Environ. Change 2020, 20, 12. [Google Scholar] [CrossRef]
- INV. Informe Anual de Superficie 2022; Instituto Nacional de Vitivinicultura (INV): Mendoza, Argentina, 2023. [Google Scholar]
- Villarreal, P.; Romagnoli, S.; Llorente, A. Pautas Tecnológicas: Vid Para Vinificar; Ediciones INTA: Buenos Aires, Argentina, 2007. [Google Scholar]
- Palese, C.; Lässig, J.L.; Cogliati, M.G.; Bastanski, M.A. Wind regime and wind power in North Patagonia, Argentina. Wind. Eng. 2000, 24, 361–377. [Google Scholar] [CrossRef]
- De Langre, E. Effects of wind on plants. Annu. Rev. Fluid Mech. 2008, 40, 141–168. [Google Scholar] [CrossRef]
- Biddington, N.L. The effects of mechanically-induced stress in plants—A review. Plant Growth Regul. 1986, 4, 103–123. [Google Scholar] [CrossRef]
- Jaffe, M.J. Thigmomorphogenesis: The response of plant growth and development to mechanical stimulation: With special reference to Bryonia dioica. Planta 1973, 114, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Dry, P.R.; Reed, S.; Potter, G. The effect of wind on the performance of Cabernet Franc grapevines. Acta Hortic. 1989, 240, 143–146. [Google Scholar] [CrossRef]
- Gardiner, B.; Berry, P.; Moulia, B. Review: Wind impacts on plant growth, mechanics and damage. Plant Sci. 2016, 245, 94–118. [Google Scholar] [CrossRef] [PubMed]
- Cleugh, H.A.; Miller, J.M.; Böhm, M. Direct mechanical effects of wind on crops. Agrofor. Syst. 1998, 41, 85–112. [Google Scholar] [CrossRef]
- Campbell-Clause, J.M. Stomatal response of grapevines to wind. Aust. J. Exp. Agric. 1998, 38, 77–82. [Google Scholar] [CrossRef]
- Freeman, B.M.; Kliewer, W.M.; Stern, P. Influence of windbreaks and climatic region on diurnal fluctuation of leaf water potential, stomatal conductance, and leaf temperature of grapevines. Am. J. Enol. Vitic. 1982, 33, 233–236. [Google Scholar] [CrossRef]
- Pienaar, J.W. The Effect of Wind on the Performance of the Grapevine; University of Stellenbosch: Stellenbosch, South Africa, 2005. [Google Scholar]
- Bettiga, L.J.; Dokoozlian, N.K.; Williams, L.E. Response of Chardonnay grapevines to windbreaks in the Salinas Valley. Am. J. Enol. Vitic. 1997, 48, 374. [Google Scholar]
- Kobriger, J.M.; Kliewer, W.M.W.M.; Lagier, S.T.T.; Gamay, N.; Seedless, T.; Kobringer, J.M.; Kliewer, W.M.W.M.; Lagier, S.T.T. Effects of wind on water relations of several grapevine cultivars. Management 1984, 35, 164–169. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Cheynier, V.; Moutounet, M.; Sarni-Manchado, P. Les composés phénoliques. In Oenologie, Fondements Scientifiques et Technologiques; Flanzy, C., Ed.; Lavoisier Tec et Doc: Paris, France, 1998; pp. 124–164. [Google Scholar]
- Keller, M. Deficit irrigation and vine mineral nutrition. Am. J. Enol. Vitic. 2005, 3, 267–283. [Google Scholar] [CrossRef]
- Teixeira, A.; Eiras-Dias, J.; Castellarin, S.D.; Gerós, H. Berry phenolics of grapevine under challenging environments. Int. J. Mol. Sci. 2013, 14, 18711–18739. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Abscisic acid’s role in the modulation of compounds that contribute to wine quality. Plants 2021, 10, 938. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.; Berli, F.J.; Fontana, A.; Piccoli, P.; Bottini, R. Malbec grape (Vitis vinifera L.) responses to the environment: Berry phenolics as influenced by solar UV-B, water deficit and sprayed abscisic acid. Plant Physiol. Biochem. 2016, 109, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Laitinen, R.A.E.; Nikoloski, Z. Genetic basis of plasticity in plants. J. Exp. Bot. 2019, 70, 739–745. [Google Scholar] [CrossRef]
- Roig-Puscama, F.; Berli, F.; Roig, F.A.; Tomazello-Filho, M.; Mastrantonio, L.; Piccoli, P. Wood hydrosystem of three cultivars of Vitis vinifera L. is modified in response to contrasting soils. Plant Soil 2021, 463, 573–588. [Google Scholar] [CrossRef]
- Dal Santo, S.; Zenoni, S.; Sandri, M.; De Lorenzis, G.; Magris, G.; De Paoli, E.; Di Gaspero, G.; Del Fabbro, C.; Morgante, M.; Brancadoro, L. Grapevine field experiments reveal the contribution of genotype, the influence of environment and the effect of their interaction (G × E) on the berry transcriptome. Plant J. 2018, 93, 1143–1159. [Google Scholar] [CrossRef]
- Köse, B.; Çelik, H. Phenological changes of shoot carbohydrates and plant growth characteristics in Vitis labrusca L. grape. Sci. Papers. Ser. B Hortic. 2017, 61, 257–268. [Google Scholar]
- Poni, S.; Lakso, A.; Turner, J.; Melious, R. The effects of pre-and post-veraison water stress on growth and physiology of potted Pinot Noir grapevines at varying crop levels. Vitis 1993, 32, 207–214. [Google Scholar]
- Chaves, M.; Zarrouk, O.; Francisco, R.; Costa, J.; Santos, T.; Regalado, A.; Rodrigues, M.; Lopes, C. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef]
- Matthews, M.A.; Anderson, M.M. Fruit ripening in Vitis vinifera L.: Responses to seasonal water deficits. Am. J. Enol. Vitic. 1988, 39, 313–320. [Google Scholar] [CrossRef]
- Matthews, M.A.; Anderson, M.M. Reproductive development in grape (Vitis vinifera L.): Responses to seasonal water deficits. Am. J. Enol. Vitic. 1989, 40, 52–60. [Google Scholar] [CrossRef]
- Keller, M.; Romero, P.; Gohil, H.; Smithyman, R.P.; Riley, W.R.; Casassa, L.F.; Harbertson, J.F. Deficit irrigation alters grapevine growth, physiology, and fruit microclimate. Am. J. Enol. Vitic. 2016, 67, 426–435. [Google Scholar] [CrossRef]
- Casassa, L.F.; Keller, M.; Harbertson, J.F. Regulated deficit irrigation alters anthocyanins, tannins and sensory properties of Cabernet Sauvignon grapes and wines. Molecules 2015, 20, 7820–7844. [Google Scholar] [CrossRef]
- Becker, N.; Zimmermann, H. Influence de divers apports d’eau sur des vignes en pots, sur la maturation des sarments, le développement des baies et la qualité des vins. Bulletin de l’OIV 1984, 57, 573–683. [Google Scholar]
- Coombe, B. Distribution of solutes within the developing grape berry in relation to its morphology. Am. J. Enol. Vitic. 1987, 38, 120–127. [Google Scholar] [CrossRef]
- Ojeda, H.; Andary, C.; Kraeva, E.; Carbonneau, A.; Deloire, A. Influence of pre-and postveraison water deficit on synthesis and concentration of skin phenolic compounds during berry growth of Vitis vinifera cv. Shiraz. Am. J. Enol. Vitic. 2002, 53, 261–267. [Google Scholar]
- Seymour, G.B.; Taylor, J.E.; Tucker, G.A. Biochemistry of Fruit Ripening; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Berli, F.J.; Alonso, R.; Beltrano, J.; Bottini, R. High-altitude solar UV-B and abscisic acid sprays increase grape berry antioxidant capacity. Am. J. Enol. Vitic. 2015, 66, 65–72. [Google Scholar] [CrossRef]
- Berli, F.J.; Alonso, R.; Bressan-Smith, R.; Bottini, R. UV-B impairs growth and gas exchange in grapevines grown in high altitude. Physiol. Plant. 2013, 149, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Berli, F.J.; Fanzone, M.; Piccoli, P.; Bottini, R. Solar UV-B and ABA are involved in phenol metabolism of Vitis vinifera L. increasing biosynthesis of berry skin polyphenols. J. Agric. Food Chem. 2011, 59, 4874–4884. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, R.; Muñoz, F.; Bottini, R.; Piccoli, P.; Berli, F.J. Effects of Wind Exposure and Deficit Irrigation on Vegetative Growth, Yield Components and Berry Composition of Malbec and Cabernet Sauvignon. Plants 2024, 13, 1292. https://doi.org/10.3390/plants13101292
Alonso R, Muñoz F, Bottini R, Piccoli P, Berli FJ. Effects of Wind Exposure and Deficit Irrigation on Vegetative Growth, Yield Components and Berry Composition of Malbec and Cabernet Sauvignon. Plants. 2024; 13(10):1292. https://doi.org/10.3390/plants13101292
Chicago/Turabian StyleAlonso, Rodrigo, Flavio Muñoz, Rubén Bottini, Patricia Piccoli, and Federico J. Berli. 2024. "Effects of Wind Exposure and Deficit Irrigation on Vegetative Growth, Yield Components and Berry Composition of Malbec and Cabernet Sauvignon" Plants 13, no. 10: 1292. https://doi.org/10.3390/plants13101292