
Alterations in the Anatomy and Ultrastructure of Leaf Blade in Norway Maple (Acer platanoides L.) Growing on Mining Sludge: Prospects of Using This Tree Species for Phytoremediation
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Microscopic Analysis
2.2.1. Sampling
2.2.2. Transmission Electron Microscopy
2.3. Measurements
2.4. Statistics
3. Results
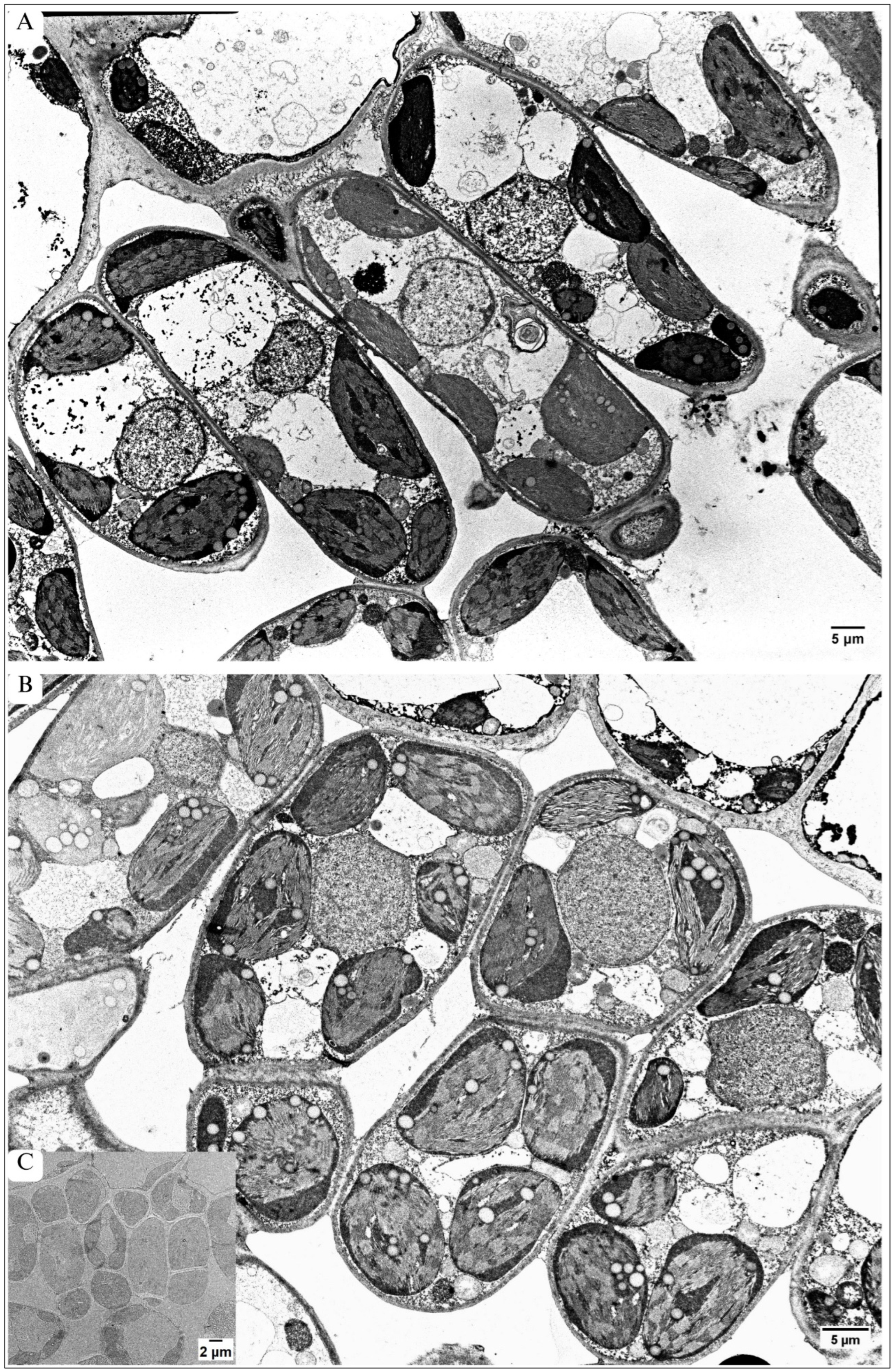
3.1. General Tree Morphology and Alterations in the Entire Leaf Blade Architecture
3.2. Alterations in the Architecture of Upper Epidermis
3.2.1. Ultrastructure of Cell Wall
3.3. Alterations in Architecture of Palisade Parenchyma
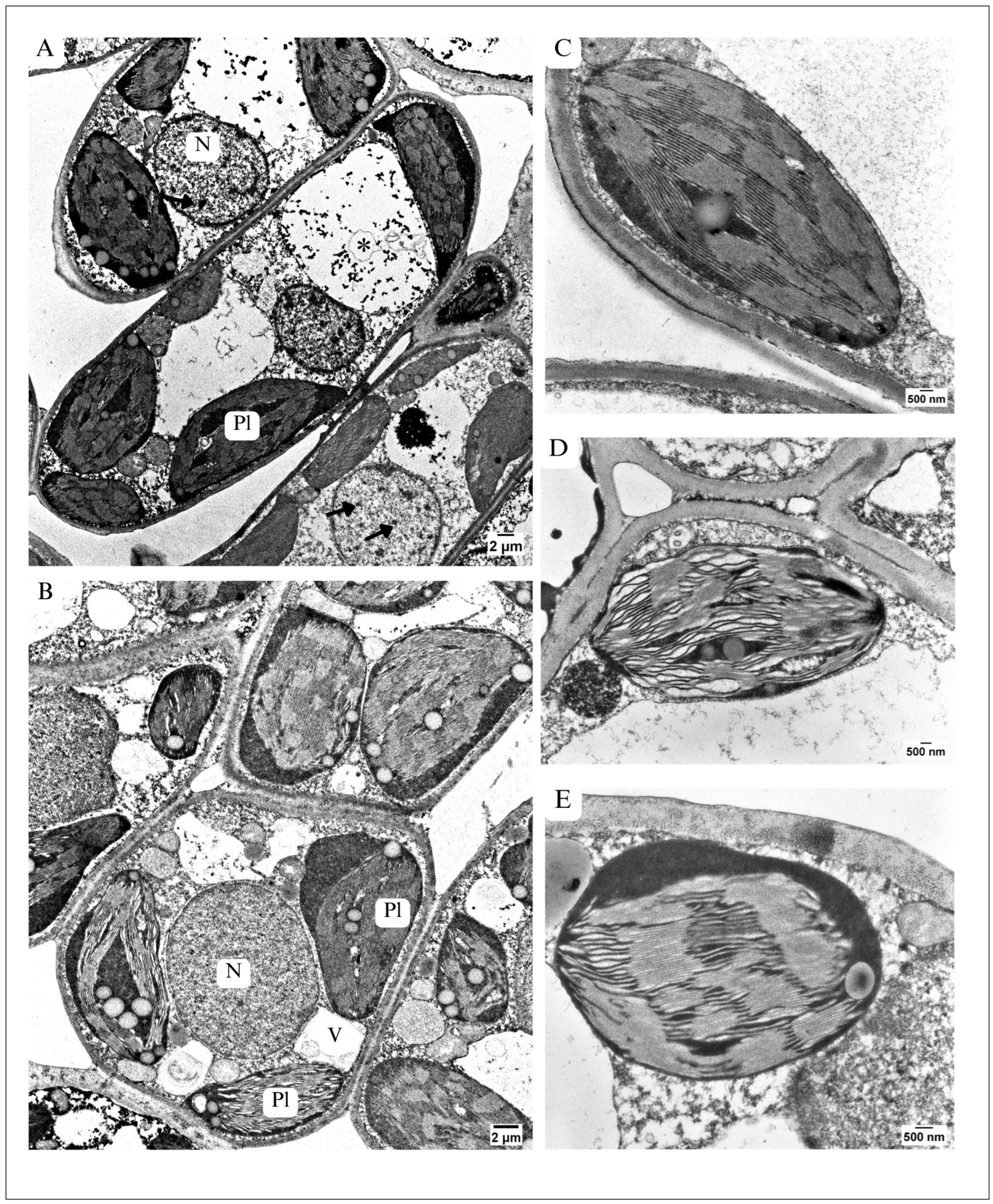
3.3.1. The Most Altered Organelles of Palisade Parenchyma Cells
Chloroplasts Ultrastructure
Ultrastructure of Nucleus
3.4. Alterations in Architecture of Spongy Parenchyma
3.5. Alterations in Architecture of Lower Epidermis
4. Discussion
4.1. Alterations in the Leaf Anatomy
4.2. Alterations at the Cellular Level
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CW | cell wall |
| IS | intercellular space |
| Maple C | Norway maple tree growing in control conditions |
| Maple S | Norway maple tree growing in mining sludge |
| TE | toxic and potentially toxic (toxic in excess) elements |
| TEM | transmission electron microscope |
References
- Drzewiecka, K.; Piechalak, A.; Goliński, P.; Gąsecka, M.; Magdziak, Z.; Szostek, M.; Budzyńska, S.; Niedzielski, P.; Mleczek, M. Differences of Acer platanoides L. and Tilia cordata Mill. Response patterns/survival strategies during cultivation in extremely polluted mining sludge—A pot trial. Chemosphere 2019, 229, 589–601. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Bin, J.; Hu, Y.; Liu, R.; Sun, W. A review on in situ phytoremediation of mine tailings. Chemosphere 2017, 184, 594–600. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Ji, B.; Khoso, S.A.; Tang, H.; Liu, R.; Wang, L.; Hu, Y. An extensive review on restoration technologies for mining tailings. Environ. Sci. Pollut. Res. Int. 2018, 25, 33911–33925. [Google Scholar] [CrossRef] [PubMed]
- Rasafi, T.E.; Haouas, A.; Tallou, A.; Chakouri, M.; Aallam, Y.; El Moukhtari, A.; Hamamouch, N.; Hamdali, H.; Oukarroum, A.; Farissi, M.; et al. Recent progress on emerging technologies for trace elements-contaminated soil remediation. Chemosphere 2023, 341, 140121. [Google Scholar] [CrossRef] [PubMed]
- König, L.A.; Medina-Vega, J.A.; Longo, R.M.; Zuidema, P.A.; Jakovac, C.C. Restoration success in former Amazonian mines is driven by soil amendment and forest proximity. Phil. Trans. R. Soc. B 2022, 378, 20210086. [Google Scholar] [CrossRef]
- Pietrzykowski, M. Tree species selection and reaction to mine soil reconstructed at reforested post-mine sites: Central and eastern European experiences. Ecol. Eng. 2019, 142S, 100012. [Google Scholar] [CrossRef]
- Ashraf, S.; Ali, Q.; Zahir, Z.A.; Ashraf, S.; Asghar, H.N. Phytoremediation: Environmentally sustainable way for reclamation of heavy metal polluted soils. Ecotoxicol. Environ. Saf. 2019, 174, 714–727. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Castro, I.; Molina, L.; Prieto-Fernández, M.Á.; Segura, A. Past, present and future trends in the remediation of heavy-metal contaminated soil—Remediation techniques applied in real soil-contamination events. Heliyon 2023, 9, e16692. [Google Scholar] [CrossRef] [PubMed]
- Mocek-Płóciniak, A.; Mencel, J.; Zakrzewski, W.; Roszkowski, S. Phytoremediation as an Effective Remedy for Removing Trace Elements from Ecosystems. Plants 2023, 12, 1653. [Google Scholar] [CrossRef]
- Bhat, S.A.; Bashir, O.; Ul Haq, S.A.; Amin, T.; Rafiq, A.; Ali, M.; Américo-Pinheiro, J.H.P.; Sher, F. Phytoremediation of heavy metals in soil and water: An eco-friendly, sustainable and multidisciplinary approach. Chemosphere 2022, 303, 134788. [Google Scholar] [CrossRef]
- Günthardt-Goerg, M.; Vollenweider, P. Phytoremediation Using Native Vegetation to Stabilize Heavy Metal Contaminants at Polluted Sites. Earth Environ. 2022, 11. Available online: https://researchoutreach.org/articles/phytoremediation-using-native-vegetation-stabilise-heavy-metal-contaminants-polluted-sites/ (accessed on 29 June 2023).
- Pulford, I.D.; Watson, C. Phytoremediation of heavy metal-contaminated land by trees—A review. Environ. Int. 2003, 29, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Mleczek, M.; Goliński, P.; Krzesłowska, M.; Gąsecka, M.; Magdziak, Z.; Rutkowski, P.; Budzyńska, S.; Waliszewska, B.; Kozubik, T.; Karolewski, Z.; et al. Phytoextraction of potentially toxic elements by six tree species growing on hazardous mining sludge. Environ. Sci. Pollut. Res. Int. 2017, 24, 22183–22195. [Google Scholar] [CrossRef]
- Günthardt-Goerg, M.; Vollenweider, P.; Schulin, R. Metal Accumulation and Biomass Production in Young Afforestations Established on Soil Contaminated by Heavy Metals. Plants 2022, 11, 523. [Google Scholar] [CrossRef]
- Madejón, P.; Domínguez, M.T.; Madejón, E.; Cabrera, F.; Maranón, T.; Murillo, J.M. Soil-plant relationships and contamination by trace elements: A review of twenty years of experimentation and monitoring after the Aznalcóllar (SW Spain) mine accident. Sci. Total Environ. 2018, 625, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Günthardt-Goerg, M.; Vollenweider, P. Linking stress with macroscopic and microscopic leaf response in trees: New diagnostic perspectives. Environ. Pollut. 2007, 147, 467–488. [Google Scholar] [CrossRef] [PubMed]
- Mitrović, M.; Pavlović, P.; Djurdjević, L.; Gajić, G.; Kostić, O.; Bojivić, S. Differences in Norway maple leaf morphology and anatomy among polluted (Belgrade City Parks) and unpolluted (Maljen MT.) Landscapes. Ecologia 2006, 25, 126–137. [Google Scholar]
- Vollenweider, P.; Cosio, C.; Günthardt-Goerg, M.S.; Keller, K. Localization and effects of cadmium in leaves of a cadmium-tolerant willow (Salix viminalis L.). Part II Microlocalization and cellular effects of cadmium. Environ. Exp. Bot. 2006, 58, 25–40. [Google Scholar] [CrossRef]
- Hermle, S.; Vollenweider, P.; Günthardt-Goerg, M.S.; McQuattie, C.J.; Matyssek, R. Leaf responsiveness of Populus tremula and Salix viminalis to soil contaminated with heavy metals and acidic rainwater. Tree Physiol. 2007, 27, 1517–1531. [Google Scholar] [CrossRef]
- Yadav, V.; Arif, N.; Kováč, J.; Singh, V.P.; Tripathi, D.K.; Chauhan, D.K.; Vaculík, M. Structural modifications of plant organs and tissues by metals and metalloids in the environment: A review. Plant Physiol. Biochem. 2021, 159, 100–112. [Google Scholar] [CrossRef]
- Guo, Z.; Gao, Y.; Yuan, X.; Yuan, M.; Huang, L.; Wang, S.; Liu, C.; Duan, C. Effects of Heavy Metals on Stomata in Plants: A Review. Int. J. Mol. Sci. 2023, 24, 9302. [Google Scholar] [CrossRef]
- Wójcik, M.; Tukiendorf, A. Accumulation and tolerance of lead in two contrasting ecotypes of Dianthus carthusianorum. Phytochemistry 2014, 100, 60–65. [Google Scholar] [CrossRef]
- Wierzbicka, M.; Panufnik, D. The adaptation of Silene vulgaris to growth on a calamine waste heap (S. Poland). Environ. Pollut. 1998, 101, 415–426. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kamińska, I.; Górecka, M.; Bederska-Błaszczyk, M. Evaluation of heavy metal-induced responses in Silene vulgaris ecotypes. Protoplasma 2019, 256, 1279–1297. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Rożańska, E.; Hanus-Fajerska, E.; Koszelnik-Leszek, A. Structural, physiological and genetic diversification of Silene vulgaris ecotypes from heavy metal-contaminated areas and their synchronous in vitro cultivation. Planta 2019, 249, 1761–1778. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Muszyńska, E.; Labudda, M. Structural Adaptation and Physiological Mechanisms in the Leaves of Anthyllis vulneraria L. from Metallicolous and Non-Metallicolous Populations. Plants 2020, 9, 662. [Google Scholar] [CrossRef]
- André, O.; Vollenweider, P.; Günthardt-Goerg, M.S. Foliage response to heavy metal contamination in Sycamore Maple (Acer pseudoplatanus L.). For. Snow Landsc. Res. 2006, 80, 275–288. [Google Scholar]
- Siemianowski, O.; Barabasz, A.; Weremczuk, A.; Ruszczyńska, S.; Bulska, E.; Williams, L.E.; Antosiewicz, D.M. Development of Zn-related necrosis in tobacco is enhanced by expressing AtHMA4 and depends on the apoplastic Zn levels. Plant Cell Environ. 2013, 36, 1093–1104. [Google Scholar] [CrossRef]
- Weremczuk, A.; Ruszczyńska, A.; Bulska, E.; Antosiewicz, D.M. NO-Dependent programmed cell death is involved in the formation of Zn-related lesions in tobacco leaves. Metallomics 2017, 9, 924. [Google Scholar] [CrossRef]
- Weremczuk, A.; Papierniak, A.; Kozak, K.; Willats, W.G.T.; Antosiewicz, D.M. Contribution of NtZIP1-like, NtZIP11 and a WAK-pectin based mechanism to the formation of Zn-related lesions in tobacco leaves. Environ. Exp. Bot. 2020, 176, 104074. [Google Scholar] [CrossRef]
- Kozak, K.; Antosiewicz, D.M. Tobacco as an efficient metal accumulator. Biometals 2023, 36, 351–370. [Google Scholar] [CrossRef] [PubMed]
- Bouazizi, H.; Jouili, H.; Geitmann, A.; El Ferjani, E. Structural changes of cell wall and lignifying enzymes modulations in bean roots in response to copper stress. Biol. Trace Elem. Res. 2010, 136, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Barceló, J.; Vázquez, M.D.; Poschenrieder, C. Structural and ultrastructural disorders in cadmium-treated bush bean plants (Phaseolus vulgaris). New Phytol. 1988, 108, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.M.; Chen, Z.; Xu, F.; Huang, L. Physiological and Cellular Ultrastructural Responses of Sesuvium portulacastrum under Cd Stress Grown Hydroponically. Plants 2023, 12, 3381. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Luo, L. Subcellular Distribution and Chemical Forms of Pb in Corn: Strategies Underlying Tolerance in Pb Stress. J. Agric. Food Chem. 2018, 66, 6675–6682. [Google Scholar] [CrossRef] [PubMed]
- Ďúranová, H.; Šimora, V.; Ďurišová, L.; Olexiková, L.; Kovár, M.; Požgajová, M. Modifications in Ultrastructural Characteristics and Redox Status of Plants under Environmental Stress: A Review. Plants 2023, 12, 1666. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Zang, J.; Shao, T.; Jiang, Q.; Li, Y.; Zhang, W.; Liu, M. Cadmium distribution and transformation in leaf cells involved in detoxification and tolerance in barley. Ecotoxicol. Environ. Saf. 2023, 249, 114391. [Google Scholar] [CrossRef] [PubMed]
- de Rigo, D.; Caudullo, G.; Houston Durrant, T.; San-Miguel-Ayanz, J. The European Atlas of Forest Tree Species: Modelling, data and information on forest tree species. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Kosiorek, M.; Modrzewska, B.; Wyszkowski, M. Levels of selected trace elements in Scots pine (Pinus sylvestris L.), silver birch (Betula pendula L.), and Norway maple (Acer platanoides L.) in an urbanized environment. Environ. Monit. Assess. 2016, 188, 598. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Biogeochemia Pierwiastków Śladowych. [Biogeochemistry of Trace Elements]; Polish Scientific Publishers: Warsaw, Poland, 1999. (In Polish) [Google Scholar]
- Siebielec, G.; Smreczak, B.; Klimkowicz-Pawlas, A.; Maliszewska-Kordybach, B.; Terelak, H.; Koza, P.; Hryńczuk, B.; Łysiak, M.; Miturski, T.; Gałązka, R.; et al. Monitoring Chemizmu gleb Ornych w Polsce w Latach 2010–2012. [The Monitoring of Chemism of Arable Soil in Poland in 2010–2012]; IUNG-PIB: Puławy, Poland; Inspekcja Ochrony Środowiska: Warszawa, Poland, 2012; pp. 94–101. (In Polish) [Google Scholar]
- Pallardy, S.G. Physiology of Woody Plants, 3rd ed.; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2008; pp. 39–86. [Google Scholar]
- Krzesłowska, M.; Woźny, A. Lead uptake, localization and changes in cell ultrastructure of Funaria hygrometrica protonemata. Biol. Plant 1996, 38, 253–259. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Ejaz, U.; Khan, S.M.; Khalid, N.; Ahmad, Z.; Jehangir, S.; Fatima Rizvi, Z.; Lho, L.H.; Han, H.; Raposo, A. Detoxifying the heavy metals: A multipronged study of tolerance strategies against heavy metals toxicity in plants. Front. Plant. Sci. 2023, 14, 1154571. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; de Sáe Melo Marques, T.C.L.L.; de Oliveira Gonçalves Nogueira, M.; de Castro, E.M.; Soares, A.M. Ecophysiological and anatomical changes due to uptake and accumulation of heavy metal in Brachiaria decumbens. Sci. Agric. 2011, 68, 566–573. [Google Scholar] [CrossRef]
- Pereira, M.P.; de Almeida Rodrigues, L.C.; Corrȇa, F.F.; de Castro, E.M.; Ribeiro, V.E.; Pereira, F.J. Cadmium tolerance in Schinus molle trees is modulated by enhanced leaf anatomy and photosynthesis. Trees 2016, 30, 807–814. [Google Scholar] [CrossRef]
- Liza, S.J.; Shethi, K.J.; Rashid, P. Effects of cadmium on the anatomical structures of vegetative organs of chickpea (Cicer arientinum L.). Dhaka Univ. J. Biol. Sci. 2020, 1, 45–52. [Google Scholar]
- Vaculík, M.; Mrázová, A.; Lux, A. Antimony (SbIII) reduces growth, declines photosynthesis, and modifies leaf tissue anatomy in sunflower (Helianthus annuus L.). Environ. Sci. Pollut. Res. Int. 2015, 22, 18699–18706. [Google Scholar] [CrossRef] [PubMed]
- de Freitas-Silva, L.; de Araújo, T.O.; da Silva, L.C.; de Oliveira, J.A.; de Araujo, J.M. Arsenic accumulation in Brassicaceae seedlings and its effects on growth and plant anatomy. Ecotoxicol. Environ. Saf. 2016, 124, 1–9. [Google Scholar] [CrossRef]
- Pita-Barbosa, A.; Williams, T.C.R.; Loureiro, M.E. Effects of short-term arsenic exposure in Arabidopsis thaliana: Tolerance versus toxicity responses. Biologia Plantarum 2019, 63, 45–53. [Google Scholar] [CrossRef]
- Souza, V.L.; Almeida, A.A.F.; Lima, S.G.; Cascardo, J.C.D.M.; Silva, D.D.C.; Mangabeira, P.A.; Gomes, F.P. Morphophysiological responses and programmed cell death induced by cadmium in Genipa americana L. (Rubiaceae). Biometals 2011, 24, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Barcelo, J.; Poschenrieder, C.; Andreu, I.; Gunse, B. Cadmium induced decrease of water stress resistance in bush bean plants (Phaseolus vulgaris L. cv. Contender) I. Effects of Cd on water potential, relative water content, and cell wall elasticity. J. Plant Physiol. 1986, 125, 17–25. [Google Scholar] [CrossRef]
- Luković, J.; Merkulov, L.; Pajević, S.; Zorić, L.; Nikolić, N.; Borisěv, M.; Karanović, D. Quantitative assessment of effects of cadmium on the histological structure of poplar and willow leaves. Water Air Soil Pol. 2012, 223, 2979–2993. [Google Scholar] [CrossRef]
- Da Silva Paiva, W.; de Araujo, M.A.; Rosalem, P.F.; Ferreira, T.C.; Bomfim, N.C.P.; Gonçalves, R.A.; Martins, A.R.; de Camargos, L.S. Lead effect on the morphophysiology of leaves and roots of Peltophorum dubium (Spreng.) Taub., a potential phytostabilizer. Int. J. Environ. Sci. Technol. 2024, 29, 2735–2746. [Google Scholar] [CrossRef]
- Silva, J.G.; Gomes, M.P.; Pereira, E.G.; Bicalho, E.; Garcia, Q.S. Initial growth of Peltophorum dubium is affected by nitrogen source and manganese concentration. J. Soil Sci. Plant Nutr. 2022, 22, 201–211. [Google Scholar] [CrossRef]
- Gerber, D.; Bechara, F.C.; Topanotti, L.R.; Nicoletti, M.F.; Azevedo, J.C. Evaluating tree survival and modeling initial growth for Atlantic Forest restoration. Cienc. Rural 2023, 53, 2023. [Google Scholar] [CrossRef]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Adv. Bot. 2014, 2014, 208747. [Google Scholar] [CrossRef]
- Čiamporová, M.; Nadubinská, M.; Banásová, V.; Ďurišová, E.; Zelinová, V.; Horak, O.; Gruber, D.; Lichtscheidl, I.K. Structural traits of leaf epidermis correspond to metal tolerance in Rumex acetosella populations growing on metal-contaminated soils. Protoplasma 2021, 258, 1277–1290. [Google Scholar] [CrossRef]
- Probst, A.; Liu, H.; Fanjul, M.; Liao, B.; Hollande, E. Response of Vicia faba L. to metal toxicity on mine tailing substrate: Geochemical and morphological changes in leaf and root. Environ. Exp. Bot. 2009, 66, 297–308. [Google Scholar] [CrossRef]
- Sitko, K.; Opała-Owczarek, M.; Jemioła, G.; Gieroń, Z.; Szopiński, M.; Owczarek, P.; Rudnicka, M.; Małkowski, E. Effect of Drought and Heavy Metal Contamination on Growth and Photosynthesis of Silver Birch Trees Growing on Post-Industrial Heaps. Cells 2022, 11, 53. [Google Scholar] [CrossRef] [PubMed]
- Rucińska-Sobkowiak, R. Water relations in plants subjected to heavy metal stresses. Acta Physiol. Plant 2016, 38, 257. [Google Scholar] [CrossRef]
- Licaj, I.; Germinario, C.; Di Meo, M.C.; Varricchio, E.; Rocco, M. The physiology and anatomy study in leaves of Saragolla and Svevo wheat cultivars under polyethylene glycol-simulated drought stress. Funct. Plant Biol. 2024, 51, FP23151. [Google Scholar] [CrossRef]
- Barceló, J.; Poschenrieder, C. Plant water relations as affected by heavy metal stress: A review. J. Plant Nut. 1990, 13, 1–37. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Timmers, A.C.J.; Mleczek, M.; Niedzielski, P.; Rabęda, I.; Woźny, A.; Goliński, P. Alterations of root architecture and cell wall modifications in Tilia cordata Miller (Linden) growing on mining sludge. Environ. Pollut. 2019, 248, 247–259. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Photosynthetic and anatomic responses of peanut leaves to cadmium stress. Photosynthetica 2008, 46, 627–630. [Google Scholar] [CrossRef]
- Araujo, M.A.; Leite, M.C.M.; Camargos, L.S.; Martins, A.R. Tolerance evaluation and morphophysiological responses of Astronium graveolens, a native Brazilian Cerrado, to addition of lead in soil. Ecotoxicol. Environ. Saf. 2020, 195, 110524. [Google Scholar] [CrossRef] [PubMed]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2011, 33, 35–51. [Google Scholar] [CrossRef]
- Meychik, N.; Nikolaeva, Y.; Kushunina, M. The significance of ion-exchange properties of plant root cell walls for nutrient and water uptake by plants. Plant Physiol. Biochem. 2021, 166, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Szymanski, D.B.; Cosgrove, D.J. Dynamic Coordination of Cytoskeletal and Cell Wall Systems during Plant Cell Morphogenesis. Curr. Biol. 2009, 19, R800–R811. [Google Scholar] [CrossRef]
- Guo, K.; Huang, C.; Miao, Y.; Cosgrove, D.J.; Hsia, K.J. Leaf morphogenesis: The multifaceted roles of mechanics. Mol. Plant 2022, 15, 1098–1119. [Google Scholar] [CrossRef]
- Gzyl, J.; Chmielowska-Bąk, J.; Przymusinski, R.; Gwozdz, E. Cadmium affects microtubule organization and posttranslational modifications of tubulin in seedlings of soybean (Glycine max L.). Front. Plant Sci. 2015, 6, 937. [Google Scholar] [CrossRef]
- Horiunova, L.; Krasylenko, Y.A.; Yemets, A.I.; Blume, Y.B. Involvement of plant cytoskeleton in cellular mechanisms of metal toxicity. Tsitol. Genet. 2016, 50, 47–59. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Goliński, P.; Szostek, M.; Mocek-Płóciniak, A.; Drzewiecka, K.; Piechalak, A.; Ilek, A.; Neumann, U.; Timmers, A.C.J.; Budzyńska, S.; et al. Morphology and Physiology of Plants Growing on Highly Polluted Mining Wastes. In Phytoremediation for Environmental Sustainability; Prasad, R., Ed.; Springer Nature: Singapore, 2021; pp. 113–149. [Google Scholar]
- Białońska, D.; Zobel, A.M.; Kuraś, M.; Tykarska, T.; Sawicka-Kapusta, K. Phenolic Compounds and Cell Structure in Bilberry Leaves Affected by Emissions from a Zn–Pb Smelter. Water Air Soil Pollut. 2007, 181, 123–133. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kral, A. Ecotype-Specific Pathways of Reactive Oxygen Species Deactivation in Facultative Metallophyte Silene vulgaris (Moench) Garcke Treated with Heavy Metals. Antioxidants 2020, 9, 102. [Google Scholar] [CrossRef]
- Krzesłowska, M.; Lenartowska, M.; Samardakiewicz, S.; Bilski, H.; Woźny, A. Lead deposited in the cell wall of Funaria hygrometrica protonemata is not stable—A remobilization can occur. Environ. Pollut. 2010, 158, 325–338. [Google Scholar] [CrossRef]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and Biological Functions in Plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Pelloux, J.; Rusterucci, C.; Mellerowicz, E.J. New insight into pectin methylesterase structure and function. Trends Plant Sci. 2007, 12, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Barceló, J.; Poschenrieder, C. Structural and ultrastructural changes in heavy metal exposed plants. In Heavy Metal Stress in Plants; Prasad, M.N.V., Hagemeyer, J., Eds.; Springer: Berlin/Heidelberg, Germany, 1999; pp. 183–207. [Google Scholar]
- Zechmann, B. Ultrastructure of plastids serves as reliable abiotic and biotic stress marker. PLoS ONE 2019, 4, e0214811. [Google Scholar] [CrossRef]
- Tan, S.; Sha, Y.; Sun, L.; Li, Z. Abiotic Stress-Induced Leaf Senescence: Regulatory Mechanisms and Application. Int. J. Mol. Sci. 2023, 24, 11996. [Google Scholar] [CrossRef]
- de Freitas, T.A.; França, M.G.; de Almeida, A.A.; de Oliveira, S.J.; de Jesus, R.M.; Souza, V.L.; Dos Santos Silva, J.V.; Mangabeira, P.A. Morphology, ultrastructure and mineral uptake is affected by copper toxicity in young plants of Inga subnuda subs. luschnathiana (Benth.) T.D. Penn. Environ. Sci. Pollut. Res. Int. 2015, 22, 15479–15494. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Wang, J.; Wu, C.; Li, S.; Hartley, W.; Wu, H.; Zhu, F.; Cui, M. Physiological response of Polygonum perfoliatum L. following exposure to elevated manganese concentrations. Environ. Sci. Pollut. Res. Int. 2018, 25, 132–140. [Google Scholar] [CrossRef]
- Madejón, E.; de Mora, A.P.; Felipe, E.; Burgos, P.; Cabrera, F. Soil amendments reduce trace element solubility in a contaminated soil and allow regrowth of natural vegetation. Environ. Pollut. 2006, 139, 40–52. [Google Scholar] [CrossRef]
- Sarathchandra, S.S.; Rengel, Z.; Solaiman, Z.M. A Review on Remediation of Iron Ore Mine Tailings via Organic Amendments Coupled with Phytoremediation. Plants 2023, 12, 1871. [Google Scholar] [CrossRef]
- Su, R.; Ou, Q.; Wang, H.; Dai, X.; Chen, Y.; Luo, Y.; Yao, H.; Ouyang, D.; Li, Z.; Wang, Z. Organic-inorganic composite modifiers enhance restoration potential of Nerium oleander L. to lead-zinc tailing: Application of phytoremediation. J. Environ. Sci. Pollut. Res. 2023, 30, 56569–56579. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement [µm/µm2] | Maple C | Significance Level for the t-Test | Maple S |
|---|---|---|---|
| Total thickness of leaf blade | 73.07 ± 14.69 | ns | 73.56 ± 7.85 |
| Width of upper epidermis | 9.81 ± 2.23 | *** | 15.13 ± 1.63 |
| Width of palisade parenchyma | 22.11 ± 3.45 | *** | 17.90 ± 2.41 |
| Width of spongy parenchyma | 31.66 ± 6.87 | ns | 31.72 ± 2.9 |
| Width of lower epidermis | 9.48 ± 2.14 | ns | 8.82 ± 0.92 |
| Area of cell in palisade parenchyma | 152.66 ± 52.13 | *** | 54.86 ± 20.84 |
| Measurement [μm] | Maple C | Significance Level for the t-Test | Maple S |
|---|---|---|---|
| Peripheral CW of upper epidermis with cuticle | 1.9 ± 0.32 | *** | 1.75 ± 0.44 |
| CW of palisade parenchyma | 0.35 ± 0.13 | ns | 0.37 ± 0.09 |
| CW of spongy parenchyma | 0.44 ± 0.11 | ns | 0.41 ± 0.1 |
| Peripheral CW of lower epidermis with cuticle | 1.21 ± 0.22 | ns | 1.18 ± 0.25 |
| Leaf Architecture Trait | Maple C | Maple S |
|---|---|---|
| General plant shape | Well-developed plant: many side branches, numerous leaves, large leaf area | Disturbed plant development: only few side branches, low number of leaves, markedly lower leaf area |
| Leaf mesophylls organization | Loose | High compactness |
| Upper epidermis | Oval and rectangular shape of cells Mean width of this tissue about 9.81 µm | Oval, rectangular and triangle shape of cells Significantly larger: 15.13 µm (36%) mean width of this tissue |
| Palisade mesophyll | 1 layer of cells, Elongated, oval-shaped cells Large IS Mean width of this tissue 22.11 µm | 1–3 layers of cells; Cell size markedly (about 60%) smaller than in control Round-shaped cells Small IS Significantly lower mean width of this tissue by 19%— 17.90 µm |
| Spongy mesophyll | Large IS | Small IS |
| Lower epidermis | Rectangular shape of cells—the longer axis arranged parallel to the leaf section | Rectangular shape of cell—the longer axis arranged perpendicular to the leaf section; occurrence of triangular cells |
| CWs of upper epidermis | Peripheral CW
| Peripheral CW
|
| CW of lower epidermis | Peripheral CW
| Peripheral CW
|
| Chloroplasts | Elliptical-shaped Thylakoid membranes run straight, regularly Not very numerous plastoglobuli | More rounded shape than in control, Irregular running of thylakoid system, swollen and wavy thylakoids Numerous, large plastoglobuli |
| Nucleus | Nucleoplasm not highly condensed, heterochromatin easy to distinguish | Nucleoplasm condensed, heterochromatin indistinguishable |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzesłowska, M.; Mleczek, M.; Luboński, A.; Weręża, K.; Woźny, A.; Goliński, P.; Samardakiewicz, S. Alterations in the Anatomy and Ultrastructure of Leaf Blade in Norway Maple (Acer platanoides L.) Growing on Mining Sludge: Prospects of Using This Tree Species for Phytoremediation. Plants 2024, 13, 1295. https://doi.org/10.3390/plants13101295
Krzesłowska M, Mleczek M, Luboński A, Weręża K, Woźny A, Goliński P, Samardakiewicz S. Alterations in the Anatomy and Ultrastructure of Leaf Blade in Norway Maple (Acer platanoides L.) Growing on Mining Sludge: Prospects of Using This Tree Species for Phytoremediation. Plants. 2024; 13(10):1295. https://doi.org/10.3390/plants13101295
Chicago/Turabian StyleKrzesłowska, Magdalena, Mirosław Mleczek, Aleksander Luboński, Karolina Weręża, Adam Woźny, Piotr Goliński, and Sławomir Samardakiewicz. 2024. "Alterations in the Anatomy and Ultrastructure of Leaf Blade in Norway Maple (Acer platanoides L.) Growing on Mining Sludge: Prospects of Using This Tree Species for Phytoremediation" Plants 13, no. 10: 1295. https://doi.org/10.3390/plants13101295