1. Introduction
Among vegetables, peppers contribute significantly to the human diet due to their high content of antioxidants, vitamins (A, C, and E, mainly), minerals, polyphenols, and pigments. However, peppers have a comparatively short storage and shelf life than other vegetables, such as root vegetables. Therefore, developing methods to preserve the nutritional content and extend the shelf life of peppers could provide significant benefits for both the food industry and the consumers. Multiple ancient civilizations have used pickling as one of the oldest preservation methods for various food products, including vegetables and fruits [
1]. Although, traditionally, fermented pickles were homemade products obtained by spontaneous fermentation, they are now evolving as a select entrée to improve quality and safety [
2]. It is well known that using a preselected mixed starter culture for lactic acid (LA) fermentation leads to superior quality in different fermented vegetables or fruits [
3].
Functional foods are defined as foods or ingredients with additional physiological benefits and nutritional value. In recent years, there has been an increase in interest in probiotics, and a significant body of research now recognises probiotic products and fermented foods as potential functional foods that may contribute to improved gut health, prevention, and the treatment of diseases.
Lactic acid bacteria (LAB) have traditionally been the main probiotics used in food processing as starter cultures, pharmaceuticals, and biological control agents. Currently, more than 62 different genera of LAB are widely used in commercial products as a safe fermentation culture. However, it is necessary to address tools of different complexity for the isolation, molecular characterization, identification, and evaluation of the probiotic properties of LAB before they can be considered [
4].
Since probiotics are expected to provide beneficial health effects to the host, they must show resistance to the acidic conditions of the stomach and the presence of bile salts and pancreatin in the small intestine. It is also essential that probiotic strains have good patterns of susceptibility to antibiotics and cell surface properties, such as auto-aggregation, cell surface hydrophobicity, auto-aggregation, and co-aggregation, to facilitate LAB colonization in the intestinal wall. In addition, functional attributes like antimicrobial and antimutagenic activities are desirable for probiotic strains to be effective [
5].
The search for new probiotic strains with unique properties continues to be an area of great interest despite the availability of numerous well-characterised probiotic strains worldwide. In this sense, there is a growing interest in exploring the probiotic and biotechnological potential of microorganisms already adapted to the food matrices in which they are found naturally. This is especially relevant in the case of bacteria isolated from red and green peppers (Capsicum annum L.), which have the advantage of being already adapted to this specific microbiome. By evaluating the functional and technological capabilities of regional pepper isolates, we identified new strains with unique properties useful in various applications, including the food and pharmaceutical industries.
The primary focus of this study was to examine the beneficial properties of lactic acid bacteria extracted from peppers in the northern region of Argentina. Given this vegetable’s crucial role in the local economy, exploring ways to increase its market value is imperative. Our main goal was to isolate and identify strains of lactic acid bacteria that can potentially promote human health by conducting extensive investigations of their functional and technological properties for the development of innovative and sustainable practices in regional agriculture.
4. Discussion
Historically, pickling is one of the oldest preservation processes of several foodstuffs, such as vegetables, fruits, fish, and meat. Pickling imparts unique and desirable changes in flavour, texture, and colour that take place over time in fermented pickles. In many cultures, pickles have been associated with many nutritional benefits over time. In particular, LAB microorganisms contribute to this process and are increasingly linked to consumer health benefits. In fact,
Lp. plantarum,
Lv. brevis,
Lactobacillus acidophilus,
Limosilactobacillus fermentum;
Leuconostoc fallax, and
Ln. mesenteroides are the most studied [
25].
Lactic acid bacteria dominate the fermented vegetable microbial community, providing several health-related properties. In concordance with our results, García-Burgos et al. (2020) stated that lactobacilli are the most prevalent microorganisms in the fermentation of fruit and vegetable juices and that species such as
Lp. plantarum,
Lactobacillus bavaricus,
Lactobacillus xylosus,
Lactobacillus bifidus, and
Lv. brevis are frequently found [
26]. In the same way, other authors reported that
Lp. plantarum,
Lv. brevis,
Ln. mesenteroides,
P. pentosaceus,
Limosilactobacillus fermentum, and
Lactococcus lactis were the main microorganisms isolated from vegetables [
27,
28].
It is important to note that the genus previously called Lactobacillus is one of the most widely used bacterial genera as probiotics, and its use as microbial food supplements has obtained the status of “Generally Recognized as Safe” (GRAS). Lactobacilli are found in the gastrointestinal tract of humans and animals, in plant- or animal-based fermented products, and in most commercially available fermented foods. In this study, bacterial isolates of peppers were evaluated for their stress tolerance, safety, technology, and probiotic properties.
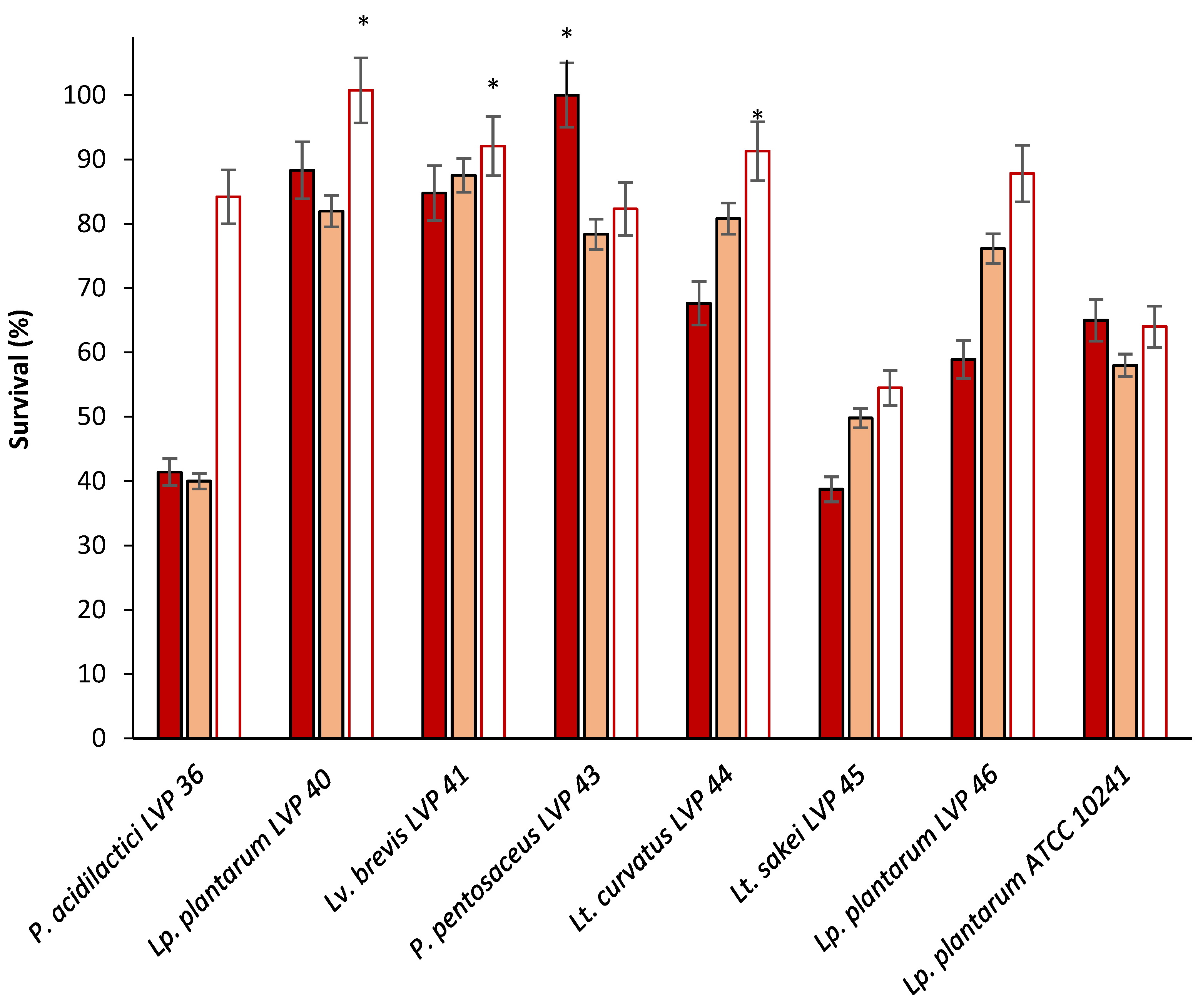
Resilience to the challenging conditions of the gastrointestinal tract stands as a primary factor restricting the application of microorganisms as live probiotic agents. The capability to endure acidity and bile salts is widely acknowledged as crucial for the survival of LAB in the gut. Specifically, the ability to survive at a pH of 3.0 is deemed as the optimal acid tolerance for probiotic strains [
29]. Consequently, this study assessed isolates for their capacity to withstand a pH of 3.0 and bile. Seven isolates were resistant to exposure to a pH of 3.0 and then were assessed for their bile tolerance. Only five strains supported the bile and were identified as lactobacilli and pediococci. This fact is in concordance with the results found in
Lacticaseibacillus rhamnosus,
Lactobacillus gasseri, and
Limosilactobacillus, which exhibited a bile resistance of 3.0 g/L [
30,
31].
The adhesion capacity of LAB isolates that had been able to survive the conditions of the digestive system was evaluated. Hydrophobicity properties are strain-specific, in concordance with an earlier report [
32]. So, the high hydrophobicity found in
Lactobacillus strains agrees with that observed in
Lactobacillus gasseri and
Lacticaseibacillus rhamnosus, and the mean values depended on the origins and genera [
31]. De Souza et al. (2019) also confirmed this by describing different strains of
Lacticaseibacillus casei with hydrophobicity values ranging from 9.66 to 69.36% and, for
Lm. fermentum strains, from 0.30 to 68.81% [
33], and they stated that bacteria with a higher hydrophobicity can adhere better to epithelial cells and significantly influence the microbial composition in the intestine.
In addition, after 24 h of incubation, all LAB strains demonstrated self-aggregation percentages higher than 70%, except for
Lv. brevis. In contrast, the auto-aggregation abilities of
Lactobacillus gasseri,
Lacticaseibacillus rhamnosus, and
Limosilactobacillus reuteri were in a variable range between 5.8 and 28.5%, depending on the incubation time and strain [
31]. Haemolytic effects were not observed in the tested isolates, and our results agreed with those reported by Damaceno et al. (2021) [
34].
On the other hand, evaluating the antibiotic resistance of potential probiotic microorganisms is vital to limit the antibiotic resistance gene transfer between the typical microbiota and pathogens [
35]. In the present study, all the strains were resistant to vancomycin, ciprofloxacin, and norfloxacin. In accordance with the present results, lactobacilli were previously reported as vancomycin- and ciprofloxacin-resistant [
29,
31]. In most
Lactobacillus species, vancomycin resistance genes are chromosomally coded; therefore, there is no risk of transfer as in the case of plasmids [
31]. Other authors noticed a variable resistance to gentamicin [
36,
37]. Nevertheless, in this work, neither of the strains was resistant to gentamicin.
The new isolates also showed a high tolerance to 2.5% of NaCl and a low tolerance to 5% of NaCl, while another study reported that several probiotics could tolerate 6.5% NaCl [
38]. The stability of pickles during the fermentation process and storage is due to their content of LAB and its viability and metabolic activity (acidifying activity). The suitability and good selectivity of LAB starter cultures for large-scale production are essential for keeping the fermenting microflora stable for a long time [
39].
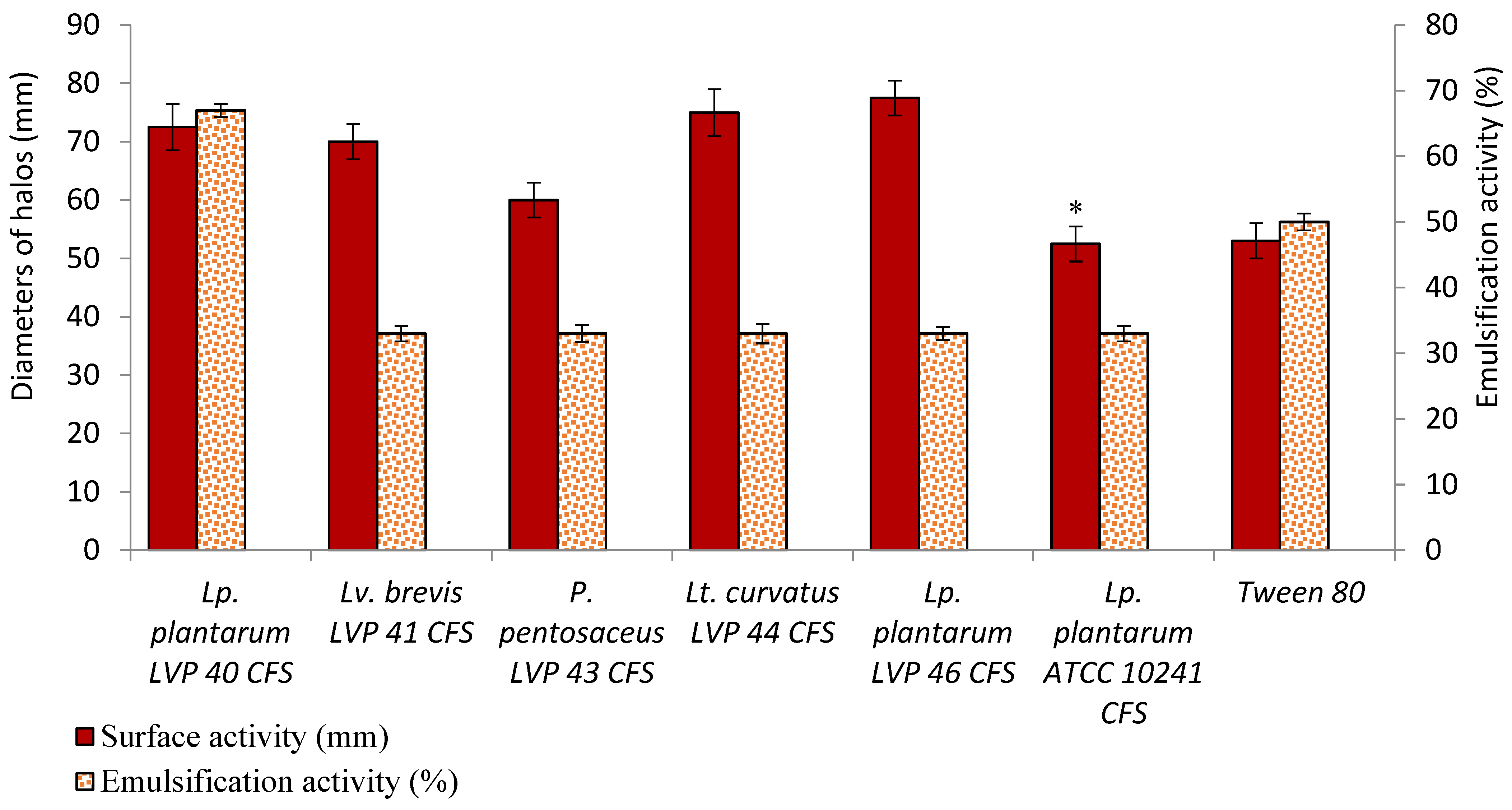
The bacterial surfactant property has also been investigated in the present work. Biosurfactants are predominantly synthesised by bacteria and fungi and can reduce interfacial and surface tension between two immiscible liquids. These compounds exhibit various properties such as detergency, wettability, and foaming, making them suitable for biomedical and industrial applications [
40].
LAB biosurfactants have shown promise as anti-adhesive agents to prevent the adhesion of pathogens to the host epithelium and solid surfaces, including biomedical instruments [
41,
42]. Exploring bacterial supernatants’ surface and interfacial activities, such as those produced by
Lactobacillus strains, is essential for identifying new strategies to inhibit microbial adherence and control biofilm-forming pathogens [
20,
43]. Previous studies have identified glycoproteinaceous biosurfactants, such as surlactin, in various
Lactobacillus species [
17,
44,
45,
46]. The findings found in this work are consistent with the results obtained by Verni et al. (2022), which revealed an emulsifying activity of the
Lactobacillus paracasei subsp.
paracasei CE75 supernatant similar to the
Lactobacillus crispatus BC1′ biosurfactant [
20,
47].
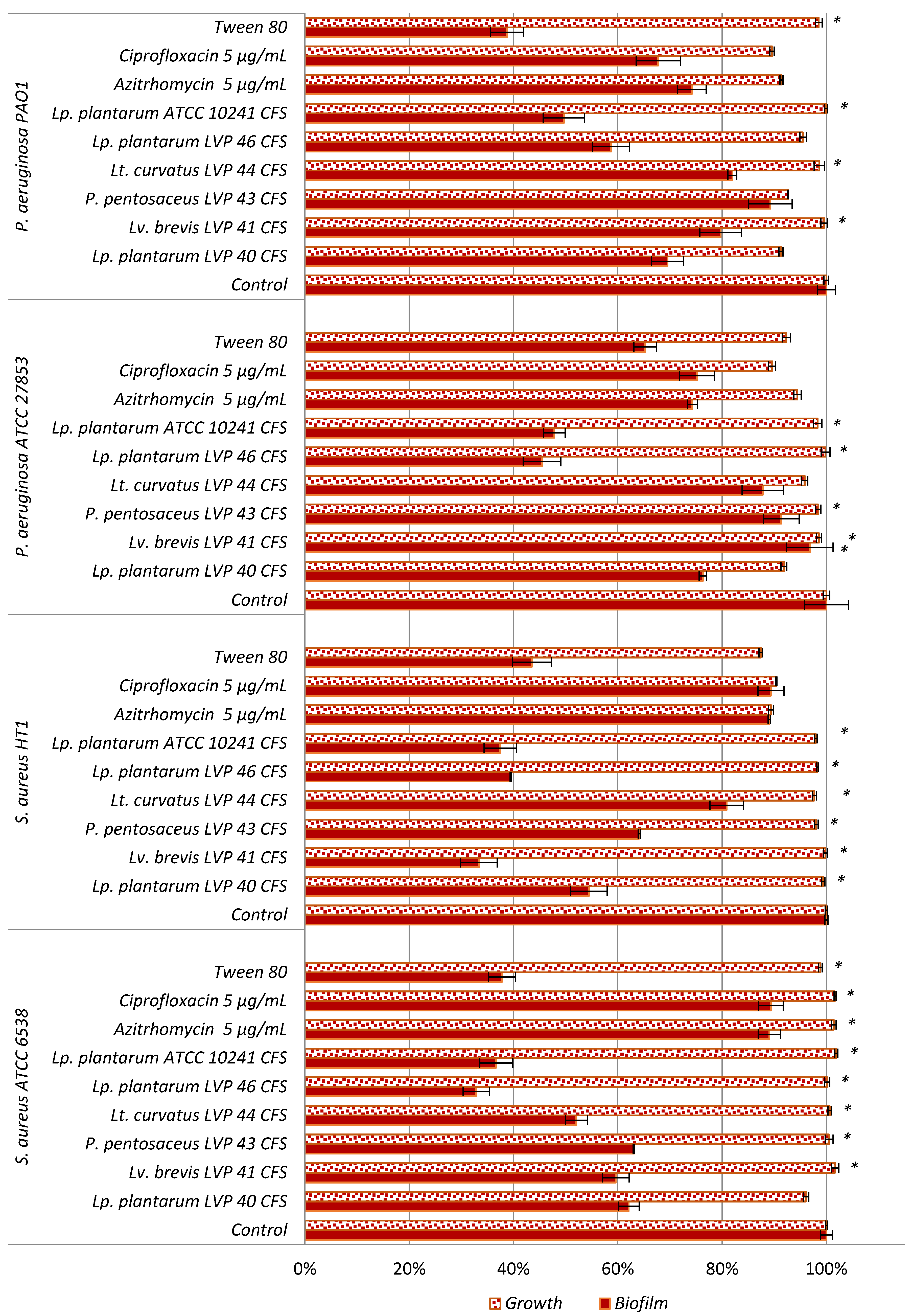
Biofilm formation is initiated by the adhesion of individual bacteria to a surface. Surface-sensing creates bacteria’s awareness of their adhering state on the surface. It is essential to initiate the phenotypic and genotypic changes that characterise the transition from initial bacterial adhesion to a biofilm. This first stage is controlled by complex combinations of the physicochemical interactions between the cell membrane and the material surface [
48,
49]. In fact, the observed effects on the biofilm biomass after only one hour of incubation did not depend on bacterial growth inhibition but rather on non-stick effects, and these results agree with previous works that demonstrated antibiofilm effects exclusively due to
Lactobacillus anti-adhesive properties [
20,
43,
50,
51]. Indeed, Gudiña et al. (2010) observed an anti-adhesive activity of the crude biosurfactant isolated from
L. paracasei subsp.
paracasei A20 higher against
S. aureus (76.80%) than against
P. aeruginosa (21.20%) at 50 mg/mL after four hours of treatment [
50]. Meanwhile, Verni et al. (2022) demonstrated a potent inhibition of
P. aeruginosa HT5 biofilm (72.01%) at one hour linked to the anti-adhesiveness effects of the
L. paracasei biosurfactant [
20]. Our results indicate that BAL supernatants are promising sources of biosurfactants with anti-adhesive properties. Specially,
Lp. plantarum LVP46′s supernatant exerts a surface activity that correlates with strong antibiofilm effects against pathogenic bacteria Gram (+) and (−).
It is important to highlight that the antibiofilm and antipathogenic effects observed in LAB supernatants were consistent with previous studies [
23,
52,
53,
54]. These results are significant since biofilm formation, governed by quorum sensing (QS), constitutes a significant problem for the safety of several food products. Probiotics have multiple properties, and, although evidence is scarce, their involvement in the regulation of QS may bring new solutions in several areas, including food preservation. Several species of lactic acid bacteria (
Lp. plantarum,
Lm. fermentum,
Lactobacillus acidophilus,
Ls. casei, Lv. brevis, Lactobacillus reuteri, and
Lt. curvatus) have already been reported at least once as quorum-quenching (QQ) agents [
23,
52,
54,
55]. For instance, a
Lp. plantarum strain reduced the expression of some genes involved in the biofilm formation of
S. aureus [
56],
Lv. brevis 3M004 inhibited biofilm formation of
P. aeruginosa [
57], and
Lp. plantarum PA 100 inhibited the
N-acyl-homoserine lactone (QS autoinducer) activity of
P. aeruginosa by blocking their synthesis [
53]. Recently, Diaz et al. demonstrated that
Ls. casei probiotic strains inhibited the biofilm formation and critical virulence factors of
S. aureus and
P. aeruginosa [
23,
54].
Co-aggregation between pathogens and probiotic strains as well as auto-aggregation are regarded as one of the critical indicators for using probiotics [
58]. The co-aggregation against
S. aureus and
P. aeruginosa was good and strain-dependent. A previous study found that
Enterococcus faecium exhibited a significant co-aggregation effect with
Salmonella enterica and
S. aureus [
32].
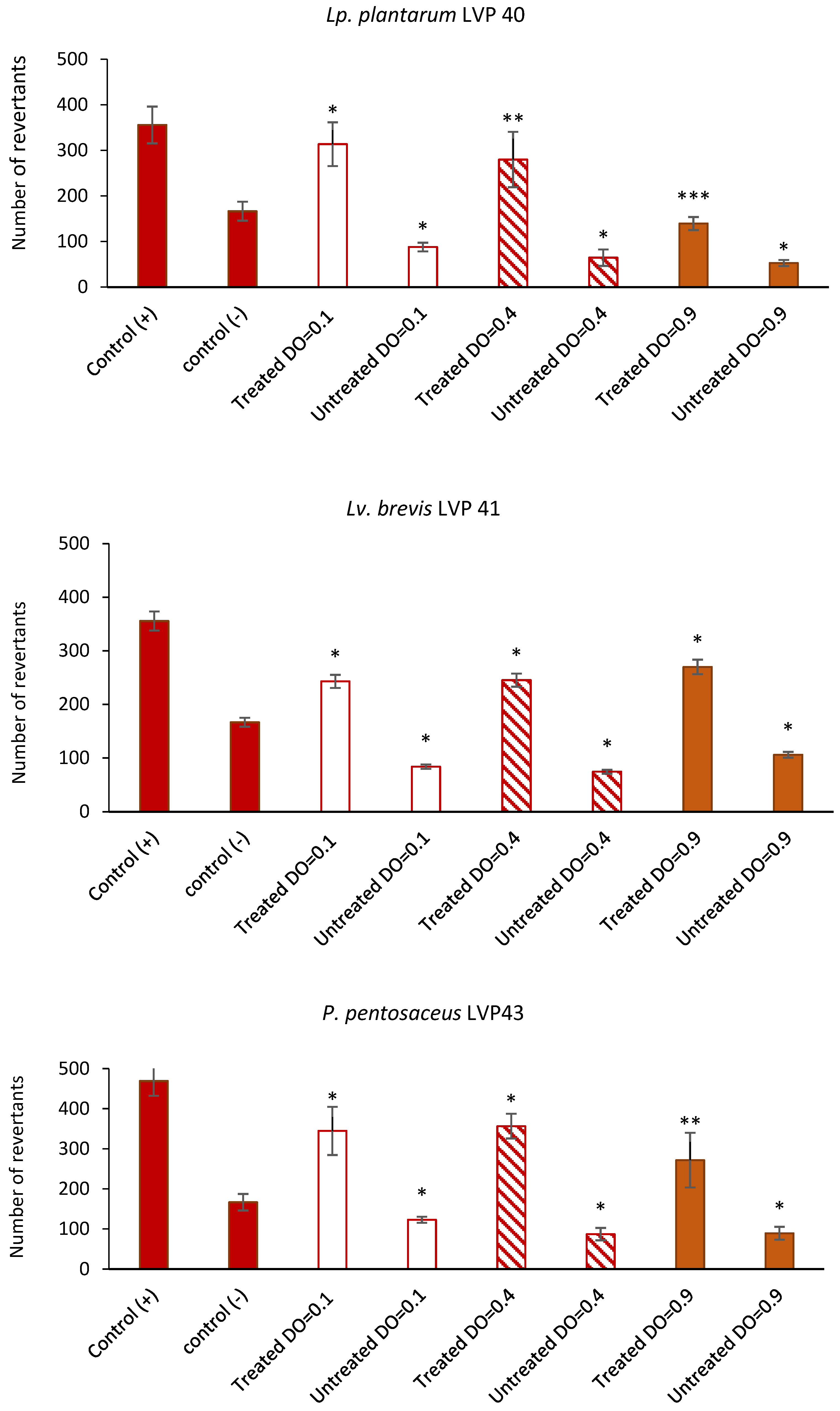
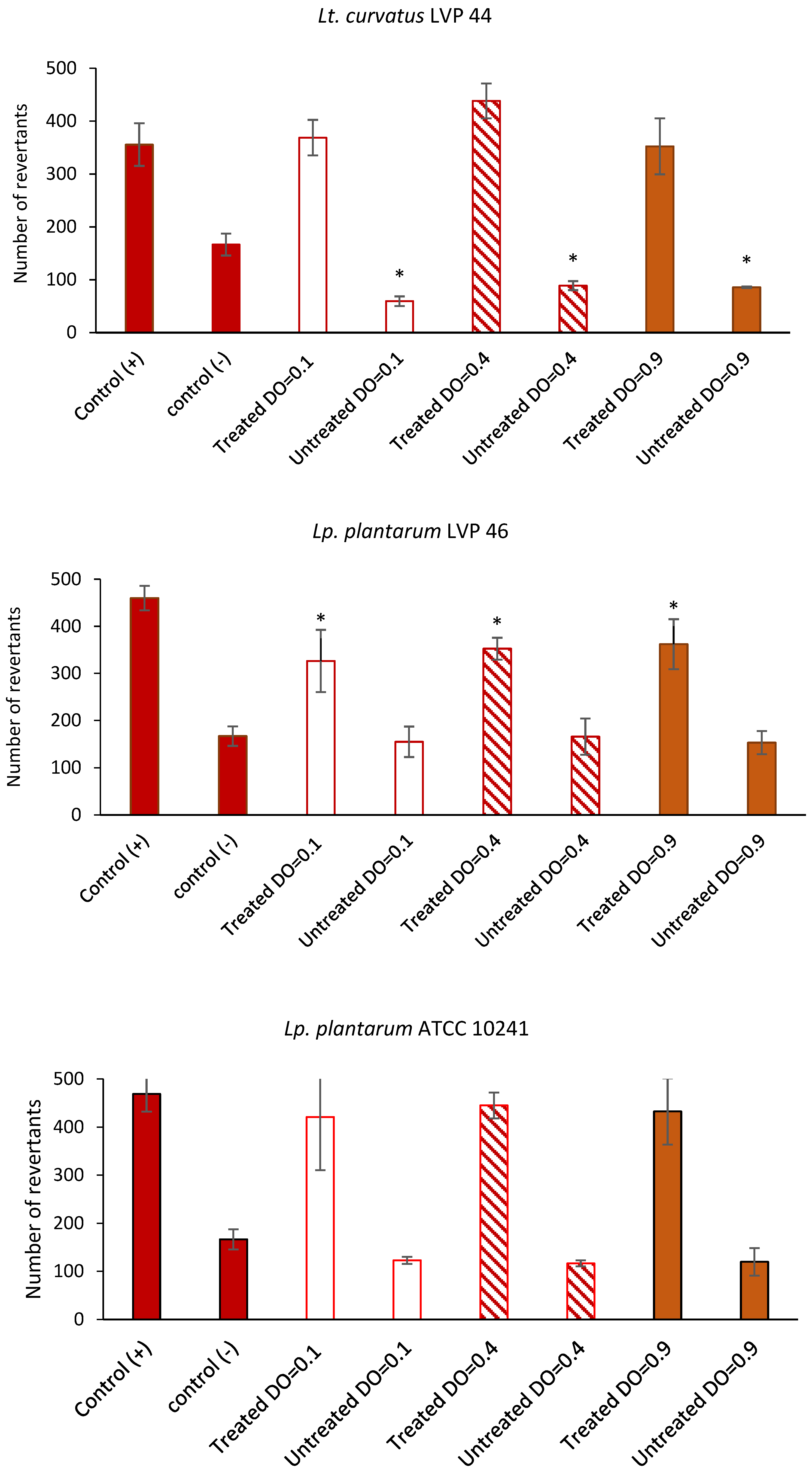
Finally, several studies suggest that fermented pickles containing probiotic strains may have a protective effect against colorectal cancer cells by potentially mitigating proliferative and mutagenic activity, suppressing the activity of the enzymes involved in the production of mutagens, carcinogens, and tumour promoters [
59]. According to the present results, previous articles have reported antimutagenic properties of lactobacilli, with this effect being strain-dependent [
5,
23]. Nonetheless, additional research is imperative to validate the antimutagenic effects of fermented pickles that contain naturally occurring probiotics.
5. Conclusions
In summary, this study assessed the functional and technological properties of 15 strains of lactic acid bacteria isolated from bell peppers, identifying them phenotypically and genotypically. The results highlight that Lp. plantarum LVP 40, Lv. brevis LVP 41, P. pentosaceus LVP 43, Lt. curvatus LVP 44, and Lp. plantarum LVP 46 exhibit notable resistance to low pH values, bile salts, and in vitro gastrointestinal conditions. These strains also display surface properties, such as auto-aggregation, hydrophobicity, co-aggregation, and anti-adhesive capacity against pathogens. Furthermore, they demonstrate superior biosurfactant and antimutagenic properties compared to the reference strain.
The technological properties, including osmotic tolerance, acidifying capacity, and compatibility between these strains, suggest the potential of these bacteria to be used in mixed starter cultures for the production of fermented foods. Overall, these characteristics indicate that the five selected strains have the potential to be promising probiotics with functional merits for application in the fermentation of bell peppers. However, to qualify as probiotics, these strains require further studies.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}