
Pharmacodynamic Evaluation and Mechanism of Ginseng Polysaccharide against Nephrotoxicity Induced by Hexavalent Chromium
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Experimental Design
2.3. Collection of Blood and Kidney Tissue
2.4. Pharmacodynamic Study of Ginseng Polysaccharide on Nephrotoxicity Induced by Hexavalent Chromium in Mice
2.4.1. Monitoring of General Signs in Mice
2.4.2. Measurement of Kidney Index
2.4.3. Detection of Renal Function Biomarkers
2.4.4. Renal Histopathological Examination
2.5. Study of the Mechanism of Ginseng Polysaccharide in Nephrotoxicity Induced by Hexavalent Chromium in Mice
2.5.1. Detection of Oxidative Stress Markers in Renal Tissue
2.5.2. Detection of Apoptosis Biomarkers
2.5.3. Detection of Cell Membrane Damage Index
2.6. Non-Targeted Lipidomics Analysis
2.6.1. Preparation of Lipid Samples
2.6.2. UPLC-Q-Orbitrap-HRMS Analysis
2.7. Statistical Analysis
3. Results
3.1. Effect of Ginseng Polysaccharide on Hexavalent Chromium-Induced Nephrotoxicity in Mice
3.1.1. Effect of Ginseng Polysaccharide on the Physical Signs of Mice
3.1.2. Effect of Ginseng Polysaccharide on Kidney Index
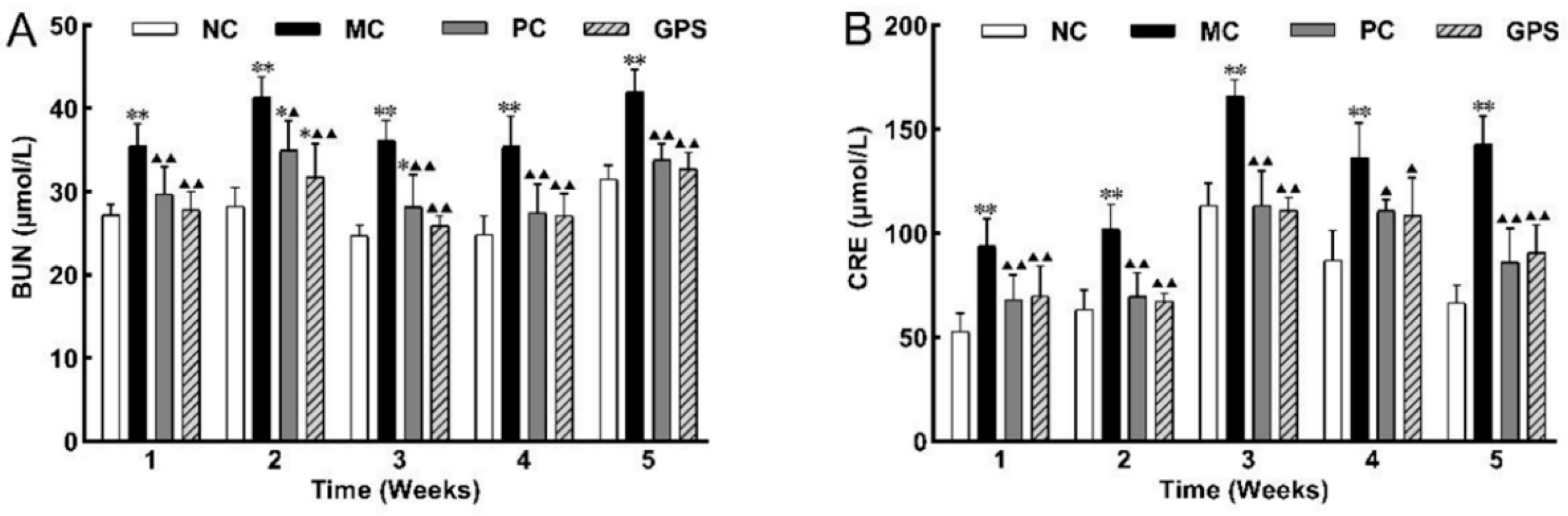
3.1.3. Effect of Ginseng Polysaccharide on Renal Function Biomarkers
3.1.4. Effect of Ginseng Polysaccharide on Renal Histopathology
3.2. Mechanism of Ginseng Polysaccharide against Hexavalent Chromium-Induced Nephrotoxicity in Mice
3.2.1. Effect of Ginseng Polysaccharide on Oxidative Stress Index
3.2.2. Effect of Ginseng Polysaccharide on Apoptosis Index
3.2.3. Effect of Ginseng Polysaccharide on Cell Membrane Damage Index
3.3. Multivariate Analysis and Structural Identification of Differential Lipids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shekhawat, K.; Chatterjee, S.; Joshi, B. Chromium Toxicity and its Health Hazards. Int. J. Adv. Res. 2015, 3, 167–172. [Google Scholar]
- Wang, Z.J.L. The Toxicity of Different Valence Chromium and Effect on Human Body. Environ. Exploit. 1997, 3, 48–50. [Google Scholar]
- Hossini, H.; Shafie, B.; Niri, A.D.; Nazari, M.; Esfahlan, A.J.; Ahmadpour, M.; Nazmara, Z.; Ahmadimanesh, M.; Makhdoumi, P.; Mirzaei, N.; et al. A comprehensive review on human health effects of chromium: Insights on induced toxicity. Environ. Sci. Pollut. Res. 2022, 29, 70686–70705. [Google Scholar] [CrossRef] [PubMed]
- Borowska, S.; Brzoska, M.M. Metals in cosmetics: Implications for human health. J. Appl. Toxicol. 2015, 35, 551–572. [Google Scholar] [CrossRef] [PubMed]
- Ming, W.E.Z. Research Overview on Hepatotoxicity and the Related Mechanism of Hexavalent Chromium Compounds. Asian J. Ecotoxicol. 2019, 14, 63–70. [Google Scholar]
- Xiong, Y.; Liu, Y.; Fu, J.; Zhang, W.; Zhen, G.; Bi, W.; Lu, X.; Chen, X. Experimental study on toxicity of potassium dichromate to kidney of Kunming mice. Occup. Health 2016, 32, 2572–2574. [Google Scholar] [CrossRef]
- Bagchi, D.; Stohs, S.J.; Downs, B.W.; Bagchi, M.; Preuss, H.G. Cytotoxicity and oxidative mechanisms of different forms of chromium. Toxicology 2002, 180, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Avila-Rojas, S.H.; Aparicio-Trejo, O.E.; Briones-Herrera, A.; Medina-Campos, O.N.; Reyes-Fermín, L.M.; Martínez-Klimova, E.; León-Contreras, J.C.; Hernández-Pando, R.; Tapia, E.; Pedraza-Chaverri, J. Alterations in mitochondrial homeostasis in a potassium dichromate model of acute kidney injury and their mitigation by curcumin. Food Chem. Toxicol. 2020, 145, 111774. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, A. Research progress of antagonists of nephrotoxicity induced by arsenic, cadmium and chromium. Chin. J. Endem. 2020, 39, 73–78. [Google Scholar]
- Liu, H.; Lu, X.; Hu, Y.; Fan, X. Chemical constituents of Panax ginseng and Panax notoginseng explain why they differ in therapeutic efficacy. Pharmacol. Res. 2020, 161, 105263. [Google Scholar] [CrossRef]
- Avila-Rojas, S.H.; Tapia, E.; Briones-Herrera, A.; Aparicio-Trejo, O.E.; León-Contreras, J.C.; Hernández-Pando, R.; Pedraza-Chaverri, J. Curcumin prevents potassium dichromate (K2Cr2O7)-induced renal hypoxia. Food Chem. Toxicol. 2018, 121, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, R.; Salama, A.; Mansour, D.; Hassan, A. Renoprotective Effect of Lactoferrin against Chromium-Induced Acute Kidney Injury in Rats: Involvement of IL-18 and IGF-1 Inhibition. PLoS ONE 2016, 11, e0151486. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Li, S.; Li, J.; Lv, Y.; Wang, X.; Wu, P.; Yang, Q.; Tang, Y.; Liu, Y.; Zhang, Z. Hexavalent chromium induces renal apoptosis and autophagy via disordering the balance of mitochondrial dynamics in rats. Ecotoxicol. Environ. Saf. 2020, 204, 111061. [Google Scholar] [CrossRef] [PubMed]
- El-Demerdash, F.M.; El-Sayed, R.A.; Abdel-Daim, M.M. Rosmarinus officinalis essential oil modulates renal toxicity and oxidative stress induced by potassium dichromate in rats. J. Trace Elem. Med. Biol. 2021, 67, 126791. [Google Scholar] [CrossRef] [PubMed]
- Cuevas-Magaña, M.Y.; Vega-García, C.C.; León-Contreras, J.C.; Hernández-Pando, R.; Zazueta, C.; García-Niño, W.R. Ellagic acid ameliorates hexavalent chromium-induced renal toxicity by attenuating oxidative stress, suppressing TNF-α and protecting mitochondria. Toxicol. Appl. Pharmacol. 2022, 454, 116242. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.S.; Wong, J.H.D.; Md Shah, M.N.; Vijayananthan, A.; Jalalonmuhali, M.; Ng, K.H. Comparison of Shear Wave Elastography and Conventional Ultrasound in Assessing Kidney Function as Measured Using 51Cr-ethylenediaminetetraacetic Acid and 99Tc-Dimercaptosuccinic Acid. Ultrasound Med. Biol. 2019, 45, 1417–1426. [Google Scholar] [CrossRef]
- Soudani, N.; Sefi, M.; Bouaziz, H.; Chtourou, Y.; Boudawara, T.; Zeghal, N. Nephrotoxicity induced by chromium (VI) in adult rats and their progeny. Hum. Exp. Toxicol. 2010, 30, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Imre, B.; García, L.; Puglia, D.; Vilaplana, F. Reactive Compatibilization of Plant Polysaccharides and Biobased Polymers: Review on Current Strategies, Expectations and Reality. Carbohydr. Polym. 2018, 209, 20–37. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Jin, Y.P.; Yao, C.L.; Wang, Y.P. Research achievements on structures and activities of polvsaccharides from Panax ginseng. China J. Chin. Mater. Medica 2014, 39, 4709–4715. [Google Scholar]
- Wang, K.; Zhang, H.; Han, Q.; Lan, J.; Chen, G.; Cao, G.; Yang, C. Effects of astragalus and ginseng polysaccharides on growth performance, immune function and intestinal barrier in weaned piglets challenged with lipopolysaccharide. J. Anim. Physiol. Anim. Nutr. 2019, 104, 1096–1105. [Google Scholar] [CrossRef]
- Abeer, M.E.-M.; Maysara, M.S.; Ayman, M. The effect of potassium dichromate on convoluted tubules of the kidney of adult male albino rats and the possible protective role of ginseng. Egypt. J. Histol. 2015, 38, 157–167. [Google Scholar] [CrossRef]
- Lester McCully, C.M.; Rodgers, L.T.; Garica, R.C.; Thomas, M.L.; Peer, C.J.; Figg, W.D.; Barnard, D.E.; Warren, K.E. Flow Rate and Apparent Volume of Cerebrospinal Fluid in Rhesus Macaques (Macaca mulatta) Based on the Pharmacokinetics of Intrathecally Administered Inulin. Comp. Med. 2020, 70, 526–531. [Google Scholar] [CrossRef]
- Liu, T.; Li, B.; Zhou, X.; Chen, H. A Study on the Time–Effect and Dose–Effect Relationships of Polysaccharide from Opuntia dillenii against Cadmium-Induced Liver Injury in Mice. Foods 2022, 11, 1340. [Google Scholar] [CrossRef]
- Yi, M.; Yi, H.; Wu, L. Accumulation and Toxicity of Hexavalent Chromium in Mice. Asian J. Ecotoxicol. 2017, 12, 259–265. [Google Scholar]
- Liu, Y.; Sui, D.; Fu, W.; Sun, L.; Li, Y.; Yu, P.; Yu, X.; Zhou, Y.; Xu, H. Protective effects of polysaccharides from Panax ginseng on acute gastric ulcers induced by ethanol in rats. Food Funct. 2021, 12, 2741–2749. [Google Scholar] [CrossRef]
- Li, W.; Wang, J.-Q.; Zhou, Y.-D.; Hou, J.-G.; Liu, Y.; Wang, Y.-P.; Gong, X.-J.; Lin, X.-H.; Jiang, S.; Wang, Z. Rare Ginsenoside 20(R)-Rg3 Inhibits D-Galactose-Induced Liver and Kidney Injury by Regulating Oxidative Stress-Induced Apoptosis. Am. J. Chin. Med. 2020, 48, 1141–1157. [Google Scholar] [CrossRef]
- Huang, Q.; Zhang, S.P.; Shi, Z.L. Role of ginseng polysaccharides in renal fibrosis via cAMP/PKA/CREB signaling pathway in diabetic nephropathy. Chin. Pharmacol. Bull. 2018, 34, 695–701. [Google Scholar]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxidative Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Feng, X.; Zeng, M.; Guan, L.; Hu, Q.; Zhong, C. Hexavalent chromium induces energy metabolism disturbance and p53-dependent cell cycle arrest via reactive oxygen species in L-02 hepatocytes. Mol. Cell. Biochem. 2012, 371, 65–76. [Google Scholar] [CrossRef]
- Marouani, N.; Tebourbi, O.; Mokni, M.; Yacoubi, M.T.; Sakly, M.; Benkhalifa, M.; Rhouma, K.B. Hexavalent Chromium-Induced Apoptosis in Rat Uterus: Involvement of Oxidative Stress. Arch. Environ. Occup. Health 2015, 70, 189–195. [Google Scholar] [CrossRef]
- Miao, L.; St. Clair, D.K. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef]
- Jin, D.; Zhang, Y.; Zhang, Y.; Duan, L.; Zhou, R.; Duan, Y.; Sun, Y.; Lian, F.; Tong, X. Panax Ginseng cA Mey. as Medicine: The Potential Use of Panax Ginseng c.A. Mey. as a Remedy for Kidney Protection from a Pharmacological Perspective. Front. Pharmacol. 2021, 12, 734151. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-T.; Zhen, J.; Pang, B.; Gu, J.-N.; Wu, S.-S. Ginsenoside Rg1 ameliorates oxidative stress and myocardial apoptosis in streptozotocin-induced diabetic rats. J. Zhejiang Univ. Sci. B 2015, 16, 344–354. [Google Scholar] [CrossRef]
- Yin, M.; Jiang, N.; Guo, L.; Ni, Z.; Al-Brakati, A.Y.; Othman, M.S.; Abdel Moneim, A.E.; Kassab, R.B. Oleuropein suppresses oxidative, inflammatory, and apoptotic responses following glycerol-induced acute kidney injury in rats. Life Sci. 2019, 232, 116634. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, X.; Ge, X.; Zhang, F. Puerarin attenuates neurological deficits via Bcl-2/Bax/cleaved caspase-3 and Sirt3/SOD2 apoptotic pathways in subarachnoid hemorrhage mice. Biomed. Pharmacother. 2018, 109, 726–733. [Google Scholar] [CrossRef] [PubMed]
- Willis, S.; Day, C.L.; Hinds, M.G.; Huang, D.C.S. The Bcl-2-regulated apoptotic pathway. J. Cell Sci. 2003, 116, 4053–4056. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Huang, X.; Zheng, L.; Zheng, F.; Dong, J.; Chen, F.; Zeng, W. Biosensors for Caspase-3: From chemical methodologies to biomedical applications. Talanta 2022, 240, 123198. [Google Scholar] [CrossRef]
- Zhong, X.; de Cássia da Silveira e Sá, R.; Zhong, C. Mitochondrial Biogenesis in Response to Chromium (VI) Toxicity in Human Liver Cells. Int. J. Mol. Sci. 2017, 18, 1877. [Google Scholar] [CrossRef]
- Wang, X.D.; Su, G.Y.; Zhao, C.; Qu, F.Z.; Wang, P.; Zhao, Y.Q. Anticancer activity and potential mechanisms of 1C, a ginseng saponin derivative, on prostate cancer cells. J. Ginseng Res. 2018, 42, 133–143. [Google Scholar] [CrossRef]
- Zhu, J.-D.; Wang, J.-J.; Zhang, X.-H.; Yu, Y.; Kang, Z.-S. Panax ginseng extract attenuates neuronal injury and cognitive deficits in rats with vascular dementia induced by chronic cerebral hypoperfusion. Neural Regen. Res. 2018, 13, 664–672. [Google Scholar] [CrossRef]
- Li, X.T.; Chen, R.; Jin, L.M.; Chen, H.Y. Regulation on Energy Metabolism and Protection on Mitochondria of Panax Ginseng Polysaccharide. Am. J. Chin. Med. 2009, 37, 1139–1152. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Baek, S.-H.; Kang, K.S.; Shin, M.-S. Characterization of macrophage activation after treatment with polysaccharides from ginseng according to heat processing. Appl. Biol. Chem. 2023, 66, 15. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.-S.; Hwang, S.-H.; Yoon, T.-J.; Kim, S.H.; Shin, K.-S. Polysaccharides from ginseng leaves inhibit tumor metastasis via macrophage and NK cell activation. Int. J. Biol. Macromol. 2017, 103, 1327–1333. [Google Scholar] [CrossRef]
- An, C.; Jiao, B.; Du, H.; Tran, M.; Song, B.; Wang, P.; Zhou, D.; Wang, Y. Jumonji domain-containing protein-3 (JMJD3) promotes myeloid fibroblast activation and macrophage polarization in kidney fibrosis. Br. J. Pharmacol. 2023, 180, 2250–2265. [Google Scholar] [CrossRef] [PubMed]
- An, C.; Jiao, B.; Du, H.; Tran, M.; Zhou, D.; Wang, Y. Myeloid PTEN deficiency aggravates renal inflammation and fibrosis in angiotensin II-induced hypertension. J. Cell. Physiol. 2021, 237, 983–991. [Google Scholar] [CrossRef] [PubMed]
- Jiao, B.; An, C.; Du, H.; Tran, M.; Wang, P.; Zhou, D.; Wang, Y. STAT6 Deficiency Attenuates Myeloid Fibroblast Activation and Macrophage Polarization in Experimental Folic Acid Nephropathy. Cells 2021, 10, 3057. [Google Scholar] [CrossRef] [PubMed]
- Jiao, B.; An, C.; Tran, M.; Du, H.; Wang, P.; Zhou, D.; Wang, Y. Pharmacological Inhibition of STAT6 Ameliorates Myeloid Fibroblast Activation and Alternative Macrophage Polarization in Renal Fibrosis. Front. Immunol. 2021, 12, 735014. [Google Scholar] [CrossRef]
- Meier, S.; Andersen, T.C.; Lind-Larsen, K.; Svardal, A.; Holmsen, H. Effects of alkylphenols on glycerophospholipids and cholesterol in liver and brain from female Atlantic cod (Gadus morhua). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 145, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Solís-Calero, C.; Ortega-Castro, J.; Frau, J.; Muñoz, F. Nonenzymatic Reactions above Phospholipid Surfaces of Biological Membranes: Reactivity of Phospholipids and Their Oxidation Derivatives. Oxidative Med. Cell. Longev. 2015, 2015, 319505. [Google Scholar] [CrossRef]
- Zalewski, A.; Macphee, C. Role of lipoprotein-associated phospholipase A2 in atherosclerosis: Biology, epidemiology, and possible therapeutic target. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 923–931. [Google Scholar] [CrossRef]
- Ilincheta de Boschero, M.G.; López, G.H.; Castagnet, P.I.; Giusto, N.M. Differential incorporation of precursor moieties into cerebral cortex and cerebellum glycerophospholipids during aging. Neurochem. Res. 2000, 25, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Eto, M.; Shindou, H.; Koeberle, A.; Harayama, T.; Yanagida, K.; Shimizu, T. Lysophosphatidylcholine Acyltransferase 3 Is the Key Enzyme for Incorporating Arachidonic Acid into Glycerophospholipids during Adipocyte Differentiation. Int. J. Mol. Sci. 2012, 13, 16267–16280. [Google Scholar] [CrossRef] [PubMed]
- Furnkranz, A.; Leitinger, N. Regulation of inflammatory responses by oxidized phospholipids: Structure-function relationships. Curr. Pharm. Des. 2004, 10, 915–921. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Administration time (Week) | |||||||||||
| 0 | 1 | 2 | 3 | 4 | 5 | |||||||
| Experiment time (Day) | ||||||||||||
| 1–10 | 11–16 | 17 | 11–23 | 24 | 11–30 | 31 | 11–37 | 38 | 11–37 | 37–44 | 45 | |
| NC | ○ | ○○ | Sacrifice (Renal Function) | ○○ | Sacrifice (Renal Function) | ○○ | Sacrifice (Renal Function) | ○○ | Sacrifice (Renal Function) | ○ | Stop/Continuous treatment | Sacrifice (Renal Function) |
| MC |  | ●○ | ●○ | ●○ | ●○ | ● | ||||||
| PC | ●□ | ●□ | ●□ | ●□ | ● | |||||||
| GPS | ●■ | ●■ | ●■ | ●■ | ● | |||||||
| Group | Administration Time (Days) | ||
|---|---|---|---|
| 1–10 | 11–30 | 31 | |
| NC | ○ | ○○ | Sacrifice (Renal Function) |
| MC | ● | ●○ | |
| PC | ●□ | ||
| GPS1 | ●■ | ||
| GPS2 | ●■ | ||
| GPS3 | ●■ | ||
| GPS4 | ●■ | ||
| GPS5 | ●■ | ||
| Condition | Positive Ion Mode | Negative Ion Mode |
|---|---|---|
| Heating temperature (°C) | 300 | 300 |
| Sheath gas velocity (arb) | 45 | 45 |
| Auxiliary gas velocity (arb) | 15 | 15 |
| Purge gas flow rate (kv) | 1 | 1 |
| Spray voltage (°C) | 3 | 2.5 |
| Ion transport tube temperature (°C) | 350 | 350 |
| S-lens voltage (%) | 50 | 50 |
| Mass scanning range of primary mass spectrometry (Da) | 200–1800 | 250–1800 |
| Group | 1 Week | 2 Weeks | 3 Weeks | 4 Weeks | 5 Weeks |
|---|---|---|---|---|---|
| NC | 0.53 ± 0.055 | 0.540 ± 0.015 | 0.548 ± 0.025 | 0.513 ± 0.034 | 0.516±0.016 |
| MC | 0.62 ± 0.048 * | 0.600 ± 0.022 ** | 0.603 ± 0.055 * | 0.576 ± 0.050 * | 0.581 ± 0.061 * |
| PC | 0.58 ± 0.050 | 0.554 ± 0.025 ▲ | 0.553 ± 0.031 ▲ | 0.519 ± 0.072 ▲ | 0.541 ± 0.041 |
| GPS | 0.60 ± 0.043 * | 0.569 ± 0.037 ▲ | 0.553 ± 0.021 ▲ | 0.530 ± 0.028 | 0.521 ± 0.041 ▲ |
| Group | Renal Index | Group | Renal Index | Group | Renal Index |
|---|---|---|---|---|---|
| NC | 0.58 ± 0.037 | MC | 0.63 ± 0.055 ** | PC | 0.59 ± 0.051 ▲▲ |
| GPS 1 | 0.60 ± 0.070 *▲ | GPS 2 | 0.61 ± 0.057 *▲ | GPS 3 | 0.58 ± 0.046 ▲▲ |
| GPS 4 | 0.59 ± 0.051 ▲▲ | GPS 5 | 0.60 ± 0.0370 *▲ |
| No. | Name | EIS Mode | Measured m/z | Retention Time (min) | Category | Mass Accuracy (ppm) | MS/MS Fragments | MC | GPS |
|---|---|---|---|---|---|---|---|---|---|
| 1 | PE(0:0/18:0) | − | 481.3156 | 3.559 | Glycerophospholipids | −2 | 78.9576; 283.2636 | ↑ | ↓ |
| 2 | Coenzyme Q9 | + | 794.6198 | 17.446 | Pregnenolone lipids | −1 | 81.0703; 197.0802 | ↓ | ↑ |
| 3 | PC (15:0/22:6(4Z,7Z,10Z,13Z,16Z,19Z)) | − | 791.5456 | 13.411 | Glycerophospholipids | −1 | 78.9575; 283.2654; 327.2320 | ↓ | ↑ |
| 4 | PE (O- 16:0/22:5(4Z,7Z,10Z,13Z,16Z)) | + | 751.5505 | 14.227 | Glycerophospholipids | −1 | 361.2726; 392.2928 | ↓ | ↑ |
| 5 | Cer (d18:2/16:0) | − | 535.4953 | 12.486 | sphingolipid | −2 | 280.2637; 534.4876 | ↑ | ↓ |
| 6 | PC (O- 18:1(11Z)/0:0) | + | 504.3477 | 2.629 | Glycerophospholipids | −2 | 86.0967; 104.1071 | ↑ | ↓ |
| 7 | PE (P- 16:0/0:0) | − | 437.2894 | 2.590 | Glycerophospholipids | −2 | 78.9576; 140.0104; 196.0369; 239.2372; 436.2820 | ↓ | ↑ |
| 8 | Cer (d18:2/23:0) | − | 633.6046 | 16.115 | sphingolipid | −2 | 280.2637; 534.4876 | ↓ | ↑ |
| 9 | Palmitoylcarnitine | + | 399.3334 | 1.943 | fatty acids | −3 | 393.2962 | ↓ | ↑ |
| 10 | Coenzyme Q8 | + | 726.5566 | 16.373 | Pregnenolone lipids | −2 | 86.0968; 664.4657; 723.5400 | ↓ | ↑ |
| 11 | PC (O- 16:1(11Z)/0:0) | + | 479.3362 | 1.706 | Glycerophospholipids | −2 | 415.2201; 433.2311 | ↑ | ↓ |
| 12 | PC (16:0/20:5(5Z,8Z,11Z,14Z,17Z)) | − | 779.5438 | 10.39 | Glycerophospholipids | −3 | 78.9576; 303.2322 | ↑ | ↓ |
| 13 | PC (P- 18:1(9Z)/0:0) | + | 505.3516 | 1.889 | Glycerophospholipids | −3 | 86.0968; 104.1072 | ↑ | ↓ |
| 14 | PE (20:5(5Z,8Z,11Z,14Z,17Z)/22:5 (7Z,10Z,13Z,16Z,19Z)) | + | 811.5120 | 11.019 | Glycerophospholipids | −3 | 812.5469 | ↑ | ↓ |
| 15 | SM (d16:1/17:0) | + | 688.5497 | 10.968 | sphingolipid | −3 | 184.0728 | ↓ | ↑ |
| 16 | PC (P- 16:0/15:1(9Z)) | + | 701.5345 | 14.205 | Glycerophospholipids | −2 | 184.0727 | ↓ | ↑ |
| 17 | TG(18:3(9Z,12Z,15Z)/20:4(5Z,8Z,11Z,1 4Z)/20:4 (5Z,8Z,11Z,14Z)) | + | 924.7156 | 18.717 | glycerolipid | −5 | 601.5157 | ↓ | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jing, B.; Wei, M.; Chen, H.; Xie, W.; An, S.; Li, J.; Wang, S.; Zhou, X. Pharmacodynamic Evaluation and Mechanism of Ginseng Polysaccharide against Nephrotoxicity Induced by Hexavalent Chromium. Nutrients 2024, 16, 1416. https://doi.org/10.3390/nu16101416
Jing B, Wei M, Chen H, Xie W, An S, Li J, Wang S, Zhou X. Pharmacodynamic Evaluation and Mechanism of Ginseng Polysaccharide against Nephrotoxicity Induced by Hexavalent Chromium. Nutrients. 2024; 16(10):1416. https://doi.org/10.3390/nu16101416
Chicago/Turabian StyleJing, Baitong, Mengyao Wei, Huaguo Chen, Wen Xie, Silan An, Jiawen Li, Shenglin Wang, and Xin Zhou. 2024. "Pharmacodynamic Evaluation and Mechanism of Ginseng Polysaccharide against Nephrotoxicity Induced by Hexavalent Chromium" Nutrients 16, no. 10: 1416. https://doi.org/10.3390/nu16101416