
Microbe-Derived Antioxidants Protect IPEC-1 Cells from H2O2-Induced Oxidative Stress, Inflammation and Tight Junction Protein Disruption via Activating the Nrf2 Pathway to Inhibit the ROS/NLRP3/IL-1β Signaling Pathway
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture and Processing
2.3. Cell Viability Assay
2.4. Measurement of Intracellular ROS
2.5. Determination of Antioxidant Enzyme Activity
2.6. Inflammatory Cytokine Detection
2.7. Reverese Transcription-Quantitative Real-Time Polymerase Chain Reaction (RT-qPCR)
2.8. Western Blot Analysis
2.9. RNA Silencing
2.10. Statistical Analysis
3. Results
3.1. MAs Demonstrate Anti-Inflammatory Properties in H2O2-Induced IPEC-1 Cells
3.2. MAs Mitigate the Inflammatory Effects Induced by H2O2 by Inhibiting the NLRP3 Inflammasome
3.3. MAs Attenuate H2O2-Induced Tight Junction Protein Disruption by Inhibiting the NLRP3 Inflammasome
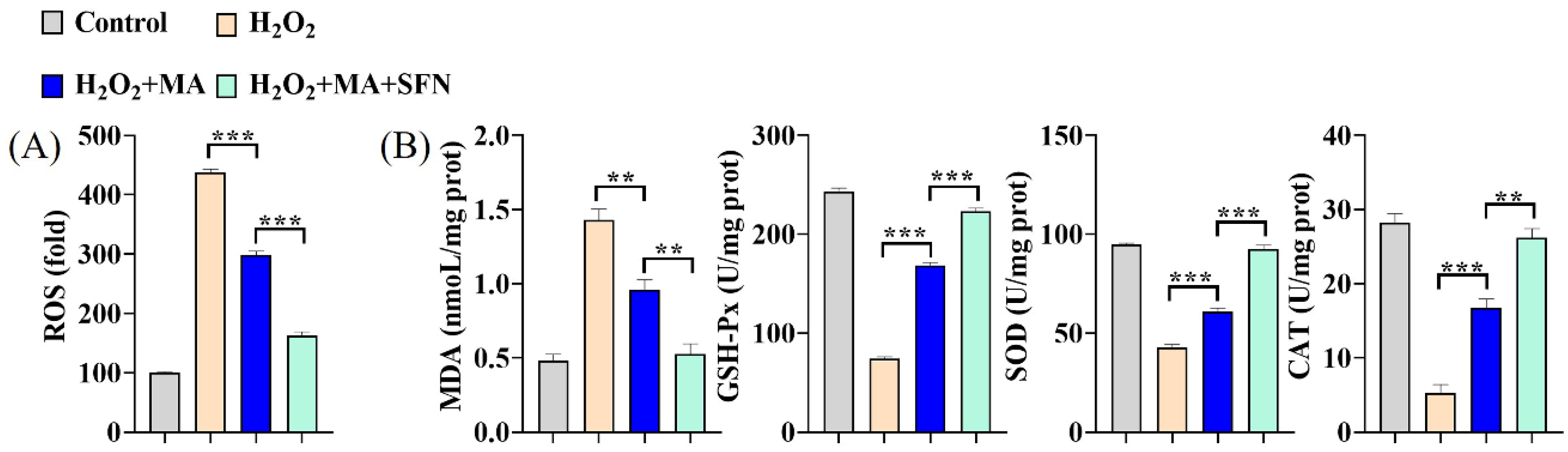
3.4. MAs Mitigate the Oxidative Stress Induced by H2O2 in IPEC-1 Cells
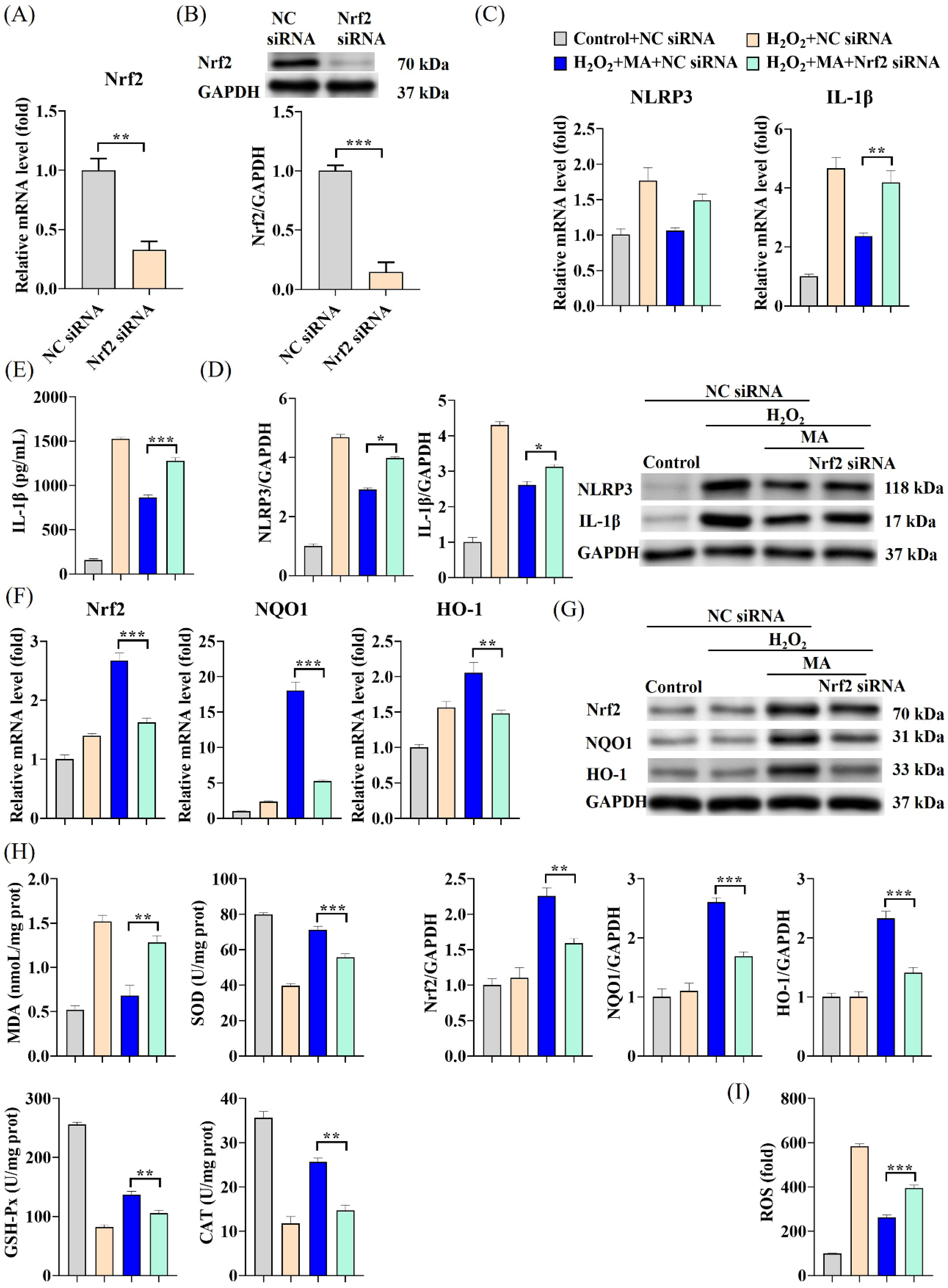
3.5. MAs Alleviate H2O2-Induced Cellular Inflammation via the Nrf2/NLRP3/IL-1β Signaling Pathway
3.6. MAs Attenuate H2O2-Induced Tight Junction Protein Disruption by Activating the Nrf2/NLRP3/IL-1β Signaling Pathway
3.7. MAs Attenuate H2O2-Induced Tight Junction Protein Disruption by Reducing IL-1β
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schett, G.; Neurath, M.F. Resolution of chronic inflammatory disease: Universal and tissue-specific concepts. Nat. Commun. 2018, 9, 3261. [Google Scholar] [CrossRef] [PubMed]
- Claytor, J.; Kumar, P.; Ananthakrishnan, A.N.; Colombel, J.-F.; Agrawal, M.; Ungaro, R.C. Mild Crohn’s Disease: Definition and Management. Curr. Gastroenterol. Rep. 2023, 25, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Kakiuchi, N.; Yoshida, K.; Uchino, M.; Kihara, T.; Akaki, K.; Inoue, Y.; Kawada, K.; Nagayama, S.; Yokoyama, A.; Yamamoto, S. Frequent mutations that converge on the NFKBIZ pathway in ulcerative colitis. Nature 2020, 577, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.F.; Shen, X.Y.; Lio, C.K.; Dai, Y.; Cheng, C.S.; Liu, J.X.; Yao, Y.D.; Yu, Y.; Xie, Y.; Luo, P.; et al. Activation of Nrf2/HO-1 Pathway by Nardochinoid C Inhibits Inflammation and Oxidative Stress in Lipopolysaccharide-Stimulated Macrophages. Front. Pharmacol. 2018, 9, 911. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Liang, M.; Sun, X.; Mohyuddin, S.G.; Chen, S.; Wen, J.; Yong, Y.; Ma, X.; Yu, Z.C.; Ju, X.H. Baicalin Alleviates LPS-Induced Oxidative Stress via NF-κB and Nrf2–HO1 Signaling Pathways in IPEC-J2 Cells. Front. Vet. Sci. 2022, 8, 1686. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Che, Q.; Chen, X.; Chen, D.; Yu, B.; He, J.; Chen, H.; Yan, H.; Zheng, P.; Luo, Y.; et al. Apple polyphenols improve intestinal antioxidant capacity and barrier function by activating the Nrf2/Keap1 signaling pathway in a pig model. J. Agric. Food Chem. 2022, 70, 7576–7585. [Google Scholar] [CrossRef] [PubMed]
- Tschopp, J.; Schroder, K. NLRP3 inflammasome activation: The convergence of multiple signalling pathways on ROS production? Nat. Rev. Immunol. 2010, 10, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhao, Y.; Ma, Y.; Wang, Z.; Rong, L.; Wang, B.; Zhang, N. Biological functions of NLRP3 inflammasome: A therapeutic target in inflammatory bowel disease. Cytokine Growth Factor Rev. 2021, 60, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zhang, J.; Xiang, J.; Li, Y.; Wu, D.; Xu, J.J. Calcitriol inhibits ROS-NLRP3-IL-1beta signaling axis via activation of Nrf2-antioxidant signaling in hyperosmotic stress stimulated human corneal epithelial cells. Redox Biol. 2019, 21, 101093. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2018, 29, 1727–1745. [Google Scholar] [CrossRef]
- Lefaki, M.; Papaevgeniou, N.; Chondrogianni, N. Redox regulation of proteasome function. Redox Biol. 2017, 13, 452–458. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Ge, M.; Jin, J.; Xu, H.; Mao, L.; Geng, S.; Wu, J.; Zhu, J.; Li, X.; Zhong, C. Mechanism investigation on Bisphenol S-induced oxidative stress and inflammation in murine RAW264.7 cells: The role of NLRP3 inflammasome, TLR4, Nrf2 and MAPK. J. Hazard. Mater. 2020, 394, 122549. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Lu, M.; Yan, W.; Li, L.; Ma, H. Dehydroepiandrosterone alleviates intestinal inflammatory damage via GPR30-mediated Nrf2 activation and NLRP3 inflammasome inhibition in colitis mice. Free. Radic. Biol. Med. 2021, 172, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Chen, X.L.; Yang, F.; Xu, J.X.; Gu, J.; Zhang, C. A Preliminary Research of Antioxidant Capacity by Micro—Derived Antioxidants in vitro. Biotechnology 2011, 21, 84–87. [Google Scholar]
- Luo, Z.; Gao, Q.Y.; Zhang, H.C.; Zhang, Y.; Zhou, S.; Zhang, J.; Xu, W.N.; Xu, J.X. Microbe-derived antioxidants attenuate cobalt chloride-induced mitochondrial function, autophagy and BNIP3-dependent mitophagy pathways in BRL3A cells. Ecotoxicol. Environ. Saf. 2022, 232, 113219. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Xu, X.; Zhao, S.; Sho, T.; Luo, W.L.; Zhang, J.; Xu, W.N.; Xu, J.X. Inclusion of microbe-derived antioxidant during pregnancy and lactation attenuates high-fat diet-induced hepatic oxidative stress, lipid disorders, and NLRP3 inflammasome in mother rats and offspring. Food Nutr. Res. 2019, 63, 3504. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.Y.; Luo, Z.; Ma, S.; Yu, C.B.; Shen, C.; Xu, W.N.; Zhang, J.; Zhang, H.C.; Xu, J.X. Microbe-Derived Antioxidants Alleviate Liver and Adipose Tissue Lipid Disorders and Metabolic Inflammation Induced by High Fat Diet in Mice. Int. J. Agric. Biol. 2023, 24, 3269. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Liu, C.C.; Tu, Z.X.; Xu, Q.; Chen, S.K.; Zhang, Y.; Wang, X.Y.; Zhang, J.; Hu, C.-A.A.; Liu, Y.L. Activation of the NF-κB and MAPK signaling pathways contributes to the inflammatory responses, but not cell injury, in IPEC-1 cells challenged with hydrogen peroxide. Oxid. Med. Cell Longev. 2020, 2020, 5803639. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Liu, X.; Tan, X.Z.; Jiang, C.X.; Chen, S.W.; Liang, G.N.; He, X.M.; Wu, J.; Chen, T.; Xu, Y. ROS-induced NLRP3 inflammasome priming and activation mediate PCB 118- induced pyroptosis in endothelial cells. Ecotoxicol. Environ. Saf. 2020, 189, 109937. [Google Scholar] [CrossRef]
- Swanson, K.V.; Deng, M.; Ting, J.P. The NLRP3 inflammasome: Molecular activation and regulation to therapeutics. Nat. Rev. Immunol. 2019, 19, 477–489. [Google Scholar] [CrossRef]
- Abderrazak, A.; Syrovets, T.; Couchie, D.; El Hadri, K.; Friguet, B.; Simmet, T.; Rouis, M. NLRP3 inflammasome: From a danger signal sensor to a regulatory node of oxidative stress and inflammatory diseases. Redox Biol. 2015, 4, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.S.; Yuan, Q.Z.; Cao, S.C.; Cui, S.M.; Xue, L.; Song, X.; Li, Z.; Xu, R.; Yuan, Q.H.; Li, R.J. Aldehyde dehydrogenase 2 inhibited oxidized LDL-induced NLRP3 inflammasome priming and activation via attenuating oxidative stress. Biochem. Biophys. Res. Commun. 2020, 529, 998–1004. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, L.W.; Al-Sadi, R.; Ma, T.Y. IL-1β and the Intestinal Epithelial Tight Junction Barrier. Front. Immunol. 2021, 12, 767456. [Google Scholar] [CrossRef] [PubMed]
- Maroni, L.C.; Agostinelli, L.; Saccomanno, S.; Pinto, C.; Giordano, D.M.; Rychlicki, C.; De Minicis, S.; Trozzi, L.; Banales, J.M.; Melum, E. Nlrp3 activation induces Il-18 synthesis and affects the epithelial barrier function in reactive cholangiocytes. Am. J. Pathol. 2017, 187, 366–376. [Google Scholar] [CrossRef]
- Feng, Y.H.; Wang, Y.; Wang, P.; Huang, Y.L.; Wang, F.J. Short-chain fatty acids manifest stimulative and protective effects on intestinal barrier function through the inhibition of NLRP3 inflammasome and autophagy. Cell Physiol. Biochem. 2018, 49, 190–205. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.H.; Huang, Y.L.; Wang, Y.; Wang, P.; Song, H.P.; Wang, F.J. Antibiotics induced intestinal tight junction barrier dysfunction is associated with microbiota dysbiosis, activated NLRP3 inflammasome and autophagy. PLoS ONE 2019, 14, e0218384. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, Y.; Jiang, X.; Du, H.; Shi, Y.; Xiu, M.; Liu, Y.; He, J. Natural products targeting Nrf2/ARE signaling pathway in the treatment of inflammatory bowel disease. Biomed. Pharmacother. 2023, 164, 114950. [Google Scholar] [CrossRef]
- Han, Y.W.; Wang, C.C.; Li, X.M.; Liang, G.B. Oleanolic acid reduces oxidative stress and neuronal apoptosis after experimental subarachnoid hemorrhage by regulating Nrf2/HO-1 pathway. Drug Dev. Res. 2022, 83, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Zhang, Y.; Zhang, C.; Chen, L.; Kuang, Y. Rutin modified selenium nanoparticles reduces cell oxidative damage induced by H2O2 by activating Nrf2/HO-1 signaling pathway. J. Biomater. Appl. 2023, 38, 109–121. [Google Scholar] [CrossRef]
- Chen, Y.F.; Li, T.X.; Tan, P.; Shi, H.; Cheng, Y.L.; Cai, T.Y.; Bai, J.J.; Du, Y.C.; Fu, W.G. Kaempferol from Penthorum chinense Pursh attenuates hepatic ischemia/reperfusion injury by suppressing oxidative stress and inflammation through activation of the Nrf2/HO-1 signaling pathway. Front. Pharmacol. 2022, 13, 857015. [Google Scholar] [CrossRef]
- Matsushima, M.; Nose, H.; Tsuzuki, H.; Takekoshi, M.; Kusatsugu, Y.; Taniguchi, H.; Ohdachi, T.; Hashimoto, N.; Sato, M.; Kawabe, T. Decrease in cholesterol in the cell membrane is essential for Nrf2 activation by quercetin. J. Nutr. Biochem. 2023, 116, 109329. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, C.; Luo, Z.; Ma, S.; Yu, C.; Lai, T.; Tang, S.; Zhang, H.; Zhang, J.; Xu, W.; Xu, J. Microbe-Derived Antioxidants Protect IPEC-1 Cells from H2O2-Induced Oxidative Stress, Inflammation and Tight Junction Protein Disruption via Activating the Nrf2 Pathway to Inhibit the ROS/NLRP3/IL-1β Signaling Pathway. Antioxidants 2024, 13, 533. https://doi.org/10.3390/antiox13050533
Shen C, Luo Z, Ma S, Yu C, Lai T, Tang S, Zhang H, Zhang J, Xu W, Xu J. Microbe-Derived Antioxidants Protect IPEC-1 Cells from H2O2-Induced Oxidative Stress, Inflammation and Tight Junction Protein Disruption via Activating the Nrf2 Pathway to Inhibit the ROS/NLRP3/IL-1β Signaling Pathway. Antioxidants. 2024; 13(5):533. https://doi.org/10.3390/antiox13050533
Chicago/Turabian StyleShen, Cheng, Zhen Luo, Sheng Ma, Chengbing Yu, Ting Lai, Shangshang Tang, Hongcai Zhang, Jing Zhang, Weina Xu, and Jianxiong Xu. 2024. "Microbe-Derived Antioxidants Protect IPEC-1 Cells from H2O2-Induced Oxidative Stress, Inflammation and Tight Junction Protein Disruption via Activating the Nrf2 Pathway to Inhibit the ROS/NLRP3/IL-1β Signaling Pathway" Antioxidants 13, no. 5: 533. https://doi.org/10.3390/antiox13050533