
A Novel PCR-Based Functional Marker of Rice Blast Resistance Gene Pi25
by
 , and
, and
Yeyang Fan
1,
Zhenhua Zhang
1,
Derun Huang
1,
Tingxu Huang
2,
Hongfei Wang
2,
Jieyun Zhuang
1 and
Yujun Zhu
1,*
1
State Key Laboratory of Rice Biology and Breeding, China National Rice Research Institute, Hangzhou 310006, China
2
Rice Research Institute, Fujian Academy of Agricultural Sciences, Fuzhou 350018, China
*
Author to whom correspondence should be addressed.
Agriculture 2023, 13(10), 1926; https://doi.org/10.3390/agriculture13101926
Submission received: 14 August 2023
/
Revised: 28 September 2023
/
Accepted: 28 September 2023
/
Published: 30 September 2023
(This article belongs to the Special Issue Innovations and Advances in Rice Molecular Breeding)
Abstract
:Rice blast is arguably the most devastating fungal disease of rice. Utilization of resistance genes to breed resistant cultivars is one of the most economical and environmentally friendly approaches to combat the disease. Pi25, a major resistance gene conferring broad-spectrum resistance to both leaf and neck blast, is an ideal gene resource to improve the resistance of rice varieties to blast. Recently, several allele-specific markers were developed. However, they were deficiently efficient due to either an additional process of restriction enzyme digestion for cleaved amplified polymorphic sequence (CAPS) markers or the risk of false-positive error in identifying susceptible Tetep allele (Pi25TTP) for PCR-based markers. In this study, based on a conserved single nucleotide polymorphism (SNP) between resistant and susceptible alleles, a tetra-primer amplification refractory mutation system (ARMS)-PCR marker was developed. The new marker, namely Pi25-2687R3, could effectively distinguish the resistant Gumei 2 (GM2) allele (Pi25GM2) and the susceptible allele Pi25TTP. Moreover, a perfect consistency of genotyping was exhibited between Pi25-2687R3 and published CAPS marker CAP3/Hpy 99I. A more accurate genotyping was also displayed compared to the previous PCR-based SNP marker Pi25-2566. Our finding proved that Pi25-2687R3 could achieve the same result as CAP3/Hpy 99I with less workload and cost and could promote the accuracy in the identification of genotypes superior to Pi25-2566. This study provided a quick and reliable functional marker for discriminating Pi25 alleles, which would be a valuable tool for genotypic assay and rice molecular breeding of blast resistance.
1. Introduction
Rice blast, caused by the fungus Magnaporthe oryzae, is arguably the most devastating fungal disease of rice [1]. Annually, 10–30% of yield losses occurred due to rice blast. However, even 10% is significant, being sufficient to feed 60 million people for one year [2]. Therefore, it is critical to adopt effective means to control the disease to ensure global food security. The exploitation of host plant resistance has generally been considered one of the most economical and environmentally friendly approaches to combat the disease [3]. A prerequisite for improving or breeding the resistant varieties is mining desired resistance (R) genes against rice blast [4,5].
Pi25 is a major R gene conferring broad-spectrum resistance to both leaf and neck blast, with a favorable allele derived from a durable resistance indica variety Gumei 2 (GM2) [6,7]. The functional allele Pi25GM2 encodes a typical coiled coil-nucleotide binding site-leucine rich repeat (CC-NBS-LRR) protein [8]. The loss of function may be probably caused by a premature stop codon or missense mutations in the coding sequence (CDS) region. For example, Pi25TTP (Tetep allele at Pi25 locus, GenBank: JQ838018.1) contains a nucleotide substitution from “a” to “g” at position 2462 from the start codon, leading to a truncated protein. Pi25ZJ100 (Zhongjian 100 allele at Pi25 locus, GenBank: JQ838019.1) contains five missense mutations (g775a, t1197c, t2444a, c2566g, g2680a, and g2687a), resulting in a susceptible (S) protein [9].
Since Pi25 is an ideal gene resource to improve the resistance to blast in rice breeding programs, several kinds of molecular markers of Pi25 have been developed and used for marker assist-selection (MAS) breeding. Firstly, simple sequence repeat markers flanking the Pi25 locus, including RM3330, RM564, and RM7193, were applied to MAS breeding of rice blast-resistant restorer lines [10,11]. Subsequently, sequence-targeted site markers tightly linked to the Pi25 locus, including SA7, Si13070C, and Si13070D, were applied to MAS breeding of elite rice parents and hybrids with improved resistance to rice blast [10,11,12,13,14]. Two kinds of markers were both PCR-based and convenient for utilization in MAS. However, they were deficient in the accuracy of genotypic identification among breeding parents and germplasm due to the certain recombination with the Pi25 locus.
Along with the map-based cloning of Pi25, the allele-specific and gene-targeted markers were achieved according to the variations in the CDS region. Wang et al. developed four sets of cleaved amplified polymorphic sequence (CAPS) markers. CAP1/Hinc II and CAP3/Hpy 99I could digest specifically the R allele Pi25GM2, according to single nucleotide polymorphism (SNP) 775 and SNP 2687, respectively. Meanwhile, CAP3/Bgl II and CAP3/Nde I could digest specifically the S allele Pi25ZJ100, according to SNP 2444 and SNP 2566, respectively [9]. These markers were used to improve the blast resistance of varieties [15,16]. Furthermore, they were also utilized to clarify the distribution of the R allele of the Pi25 gene in rice germplasm in China [17,18,19,20,21]. Nevertheless, the CAPS marker requires an additional restriction enzyme digestion process after PCR amplification, resulting in the limit in simplicity and low cost.
Recently, a PCR-based functional SNP marker was developed by Zhou et al. [22]. The principle of primer design is similar to the method of tetra-primer amplification refractory mutation system (ARMS)-PCR marker [23]. The 3′-terminal nucleotide of the inner primer corresponded to the site of the SNP, and an artificial mismatch at position −3 from 3′-terminus was introduced. Consequently, allele-specific PCR product would be amplified due to higher efficiency with matched primer and lower efficiency with mismatched one. This marker was based on the SNP at the site of 2566, where the resistant allele Pi25GM2 carried “c”, while the susceptible allele Pi259311 carried “g”. However, Zhou et al. did not assess the susceptible allele Pi25TTP, which owned the same nucleotide as Pi25GM2 at this site. This conflict would bring the risk of false-positive identification of the resistant allele of the Pi25 gene when this marker was used.
To improve the convenience and accuracy of the allele-specific marker for the Pi25 gene, we developed a novel tetra-primer ARMS-PCR marker in this study. The new marker was based on the SNP at the site of 2687, where the resistant allele and the susceptible allele were highly conserved in “g” and “a”, respectively. For effective screening, we designed multiple primer pairs and conducted examinations using different annealing temperatures. For validation of the accuracy and reliability of the new marker, we also assayed various genotypes and rice germplasm compared with previously developed markers.
2. Materials and Methods
2.1. Plant Materials
Four sets of rice materials were applied in this study. (1) GM2, Zhong 156 (Z156), and TTP were used to screen the primer pairs of tetra-primer ARMS-PCR marker; (2) Ten varieties with known alleles at Pi25 locus were employed to validate the accuracy of the selected marker. They included GM2, Z156, TTP, ZJ100, Gumei 4 (GM4), Lijiangxintuanheigu (LTH), IRBLz-Fu, IRBLzt-T, IRBLz5-CA and IRBL9-W. The latter four are near-isogenic lines (NILs) of LTH and contain multiple alleles Piz, Pizt, Pi2, and Pi9 at Piz locus, respectively [24]; (3) Twelve breeding restorer lines and five rice hybrid varieties with different blast reactions were genotyped to verify the correlation between the genotype and phenotype; (4) Ninety-four rice landraces collected from 23 countries worldwide were genotyped to identify Pi25 alleles (Table 1).
LTH, IRBLz-Fu, IRBLzt-T, IRBLz5-CA, and IRBL9-W were provided by the Zhejiang Academy of Agricultural Sciences, Zhejiang Province, China, while the others were from the China National Rice Research Institute.
2.2. Primer Design
The design of primer pairs was followed Ye’s method [23]. Because SNP 2687 is located very close to the 3′ end of CDS (2775 bp) (GenBank: HM448480.1), and the CDS sequence of Pi25TTP is only 2463 bp (GenBank: JQ838018.1), we re-sequenced GM2, Z156 and TTP alleles at Pi25 locus and extended the nucleotide sequence to about 3000 bp. The forward and reverse outer primers were designed 100~300 bps away from SNP 2687 using Oligo Primer Analysis Software Version 7.0. The forward and reverse inner primers were determined using the following principles: (1) SNP 2687 was put in the 3′ terminus of either forward primer or reverse primer (Figure 1); (2) Extra destabilizing mismatches were introduced at the position −2 and −3 from the 3′ terminus in order to increase the specificity in amplification of each allele (Figure 1); (3) The length of primer was adjusted to ensure the similar Tm value output within the paired primers using Oligo Primer Analysis Software Version 7.0.
Near SNP 2687, additional SNP (g2680a) was occurred. At SNP 2680, the R allele Pi25GM2 and the S allele Pi25TTP owned the identical nucleotide “g”, while another susceptible allele Pi25ZJ 100 carried “a” (Figure 1). To obtain the best effect of amplification and specificity, we carried out two ways to design the inner primers based on whether two SNPs were involved in producing an R-specific fragment or an S-specific fragment. (A) The oligonucleotide of the Pi25GM2 allele containing both “g” at SNP 2680 and SNP 2687 was used as the template of forward inner (FI) primer-R. The R-specific fragment would be produced when FI primer-R is paired with a common reverse outer (RO) primer. On the other hand, the S-specific fragment was generated using the reverse inner (RI) primer-S based on the antisense strand of Pi25Z156 with a common forward outer (FO) primer (Figure. 1A). (B) The oligonucleotide of the Pi25Z156 allele containing both “a” at SNP 2680 and SNP 2687 was used as the template of forward inner (FI) primer-S. The S-specific fragment would be generated using FI primer-S and common RO primer. The R-specific fragment was produced using FO primer and RI primer-R based on the antisense strand of Pi25GM2 (Figure 1B).
As a result, two sets of primer pairs were employed. The first set (set A) consisted of three pairs, namely R1, R2 and R3. They shared the same primers, 2687-FO, 2687-RO, and 2687-FI-R, but differed in the 2687-RI-S primer with multiple artificial mismatches, such as TT/CC, TT/GG, and TT/AA, respectively. Another one (set B) is composed of F1, F2, and F3 primer pairs. The difference occurred in the 2687-FI-S primers with GA mutated to TC, TG, and TT, respectively (Table 2).
2.3. DNA Extraction and Marker Assay
Genomic DNA was isolated from the fresh leaves of each plant following the mini-preparation protocol of Zheng et al. [25].
PCR was performed using 2×Taq MasterMix (CWBIO, Beijing, China), 0.2 μM of each primer, and ~50 ng of template DNA in a 15-μL reaction volume. Reaction conditions were: initial denaturation at 94 °C for 2 min, followed by 30 cycles of denaturation at 94 °C for 30 s; annealing for 30 s; and extension at 72 °C for 30 with a final extension at 72 °C for 5 min. For screening primer pairs, gradient annealing temperatures were used and set to be 50 °C, 53 °C, 55 °C and 58 °C, respectively. For the final primer pair selected, the annealing temperature was optimized to 53 °C. PCR products were electrophoretically separated in 2% agarose, followed by staining with GelRed dye (Biotium, CA, USA) and photographing with Gel Doc XR+ system (BIO-RAD, CA, USA).
2.4. Rice Blast Resistance Evaluation
Seventeen breeding materials were planted in the blast nursery, Chadi town, Shanghang country, Longyan city, Fujian Province, China. Resistance to leaf blast and neck blast was investigated after the susceptible control variety was gotten susceptible at the seedling and maturity stages, respectively. The evaluation system was employed following the national standard NY/T2646-2014, which was finally divided into six resistance grades. The details are high resistance (HR), resistance (R), moderate resistance (MR), moderate susceptibility (MS), susceptibility (S) and high susceptibility (HS), respectively.
3. Results
3.1. Selection of Primer Pairs
For set A of primer pairs, R-specific fragment was generated in the length of 318 bp, whereas S-specific fragment was 193 bp in R1 and R2 and 196 bp in R3, respectively. For set B, R-specific and S-specific product was 196 bp and 320 bp in length, respectively. For both sets, a 469 bp-length fragment could be amplified as the common product (Figure 2).
Three expected products, including common fragment, R-specific fragment, and S-specific fragment, were all clearly visible at 50 °C except R2. When the annealing temperature was increased to 58 °C, amplicons appeared very weak or even invisible for all pairs. Furthermore, the bands of the R-specific fragment and S-specific fragment were all brighter in R3 than in R1 at 50 °C, 53 °C and 55 °C, respectively. On the other hand, the product of TTP was weak or even missing at 53 °C and 55 °C in F2 and F3, while the amplicon of GM2 was much weaker than Z156 and TTP at 50 °C, 53 °C and 55 °C in F1 (Figure 2).
Therefore, R3, comprising 2687-FO, 2687-RO, 2687-FI-R, and 2687-RI-S3, was selected as the final primer pair to conduct the next investigations. The marker was consequently named Pi25-2687R3. The common, R-specific, and S-specific fragment was 469 bp, 318 bp, and 196 bp in length, respectively. Considering the non-specific amplification at 50 °C, as well as weaker amplification at 55 °C, an optimal 53 °C was recommended in the PCR reaction.
3.2. Validation of Newly Developed Marker
To validate the accuracy of the newly developed marker, we performed genotyping using 10 varieties with known alleles at the Pi25 locus. As Wang’s report, only GM2, GM4, and C101A51 (the donor of Pi2) possessed the R allele, and others carried the S allele when they were genotyped by CAPS marker CAP3/Hpy 99I [9]. Using Pi25-2687R3, GM2, GM4, and IRBLz5-CA (a NIL of LTH carrying Pi2 allele) showed the R-specific band, and others showed the S-specific band (Figure 3). It proved that Pi25-2687R3 could obtain the identical result with CAP3/Hpy 99I.
Meanwhile, we carried out genotyping using a previous marker developed by Zhou et al. [21]. Because Zhou’s marker was not given a name, here we temporarily named it Pi25-2566 due to its design based on SNP2566. It was as expected that all the varieties presented the same pattern between Pi25-2687R3 and Pi25-2566, except TTP (Figure 3). TTP displayed a distinct pattern, which corresponds to the fact that TTP presents the identical nucleotide of Z156 at SNP2687 and that of GM2 at SNP2566. The result indicated that Pi25-2687R3 could greatly promote the accuracy of characterizing the Pi25GM2 allele compared to Pi25-2566.
Furthermore, 12 restorer lines and five hybrid varieties were employed to verify the effectiveness of Pi25-2687R3. Among 12 restorer lines, 10 were identified to carry the homozygous R allele, and two lines were homozygous S allele, whereas all hybrid varieties took the heterozygous allele (Figure 4). At the same time, the rice blast resistances of these lines/varieties were investigated in the blast nursery. Ten lines carrying the homozygous R allele, as well as five hybrids with heterozygous allele, displayed resistance grade “R” or “MR”, whereas two lines taking the homozygous S allele exhibited “S” or “MS” (Table 3). Results showed a perfect correlation between the Pi25-2687R3 genotype and the observed phenotype. The Pi25GM2 presence is consistent with the resistance to rice blast, indicating Pi25-2687R3 could be a potential functional marker to predicate the presence of the Pi25GM2 allele with resistance to rice blast.
3.3. Genotyping Germplasm Using Allele-Specific Markers
Ninety-four rice landraces from 23 countries worldwide were utilized to genotype alleles at the Pi25 locus. To compare the efficiency of a new marker, published markers CAP3/Hpy 99I and Pi25-2566 were also used.
A perfect consistency was revealed between Pi25-2687R3 and CAP3/Hpy 99I. Only two accessions were identified to carry the Pi25GM2 allele. They were 5173 (No. 13) and PANAMA 1537 (No. 66), collected from Colombia and Panama, respectively (Figure 5, Table 1). It further proved that Pi25-2687R3 was as reliable as CAP3/Hpy 99I but with less workload and cost.
For Pi25-2566, a total of 10 accessions were observed to carry the same pattern as Pi25GM2. Among 10 accessions, 5173 (No. 13) and PANAMA 1537 (No. 66) were stably identified. The others might be false-positive, which were MA HNAN THU KA (No. 7), C.CORTO 1 (No. 16), PADI SEGUTUK (No. 49), KHARA GANJA (No. 63), TAK RATIA (No. 65), CHAH NONG NGHE AN (No. 89), RE BAU (No. 93) and RE CHANH (No. 94), respectively (Figure 5, Table 1). Results supported that Pi25-2687R3 was superior to Pi25-2566 in the accuracy of characterizing the Pi25GM2 allele.
4. Discussion
The ability to quickly and reliably select desirable materials and to eliminate individuals that contain favorable alleles is critical to the success of a plant breeding program [26]. With the progress of cloning of blast resistance genes, new gene-target markers or functional markers showed more accuracy than linked markers due to gene/functional markers being complete co-segregation and non-recombination with the target gene. For rice blast resistance gene Pi25, several gene/functional markers have been developed [9,22]. However, they were deficiently efficient due to either an additional process of restriction enzyme digestion for CAPS markers or the risk of false-positive error in identifying susceptible Pi25TTP for Pi25-2566. In the present study, we provided a new functional marker, Pi25-2687R3, which can specifically identify Pi25GM2 and efficiently distinguish it from Pi25TTP. The investigations demonstrated that Pi25-2687 was accurate and reliable.
Pi25GM2 has been successfully utilized in modern breeding to improve the blast resistance of elite rice parents, hybrids, and conventional varieties [10,11,12,13,14,15,16,22,27]; a low frequency of the R allele was found in the germplasm in China [9,17,18,19,20,21], implying Pi25GM2 is rare. Furthermore, little is known about the distribution of Pi25GM2 abroad, which would largely limit the utilization of this important gene. In this work, we employed Pi25-2687 to genotype 94 rice landraces from 23 countries worldwide. Only two accessions were identified to possess Pi25GM2, indicating a similar low frequency was also found in other countries. The two landraces were 5173 and PANAMA 1537, from Colombia and Panama, respectively. It was known that 5173 was also the donor of the resistant Pi2 allele, and the favorable Pi2 allele of C101A51 was originally introduced from 5173 [28]. Thus, it could be understandable that 5173 carries the resistant allele Pi25GM2. Geographically, Panama is situated in the south of Central America, and Colombia is located in the north of South America. Two countries are adjacent to each other, indicating 5173 and PANAMA 1537 might possibly have a close relationship. The exploitation of two rice landraces would provide a valuable resource for the utilization of the R allele of the Pi25 gene.
Tetra-primer ARMS-PCR marker, a SNP-based allelic-specific marker, is simple and cost-effective [29]. Hence, it has been successfully utilized in rice MAS breeding, such as rice blast resistance, herbicide resistance, and rice quality [30,31,32]. However, the usual tetra-primer ARMS-PCR has a laborious process for optimization [33]. In this study, we conducted various examinations using six combinations of primer pairs and optimized the annealing temperature. In this process, two phenomena were observed.
(1) When the annealing temperature was increased from 50 °C to 58 °C, the amplified products were changed from brighter to weaker or even invisible. We finally chose 53 °C as the annealing temperature for the Pi25-2687R3 marker because of the non-specific amplification at 50 °C and weaker amplification at 55 °C. However, it does not mean that other primer pairs could not be used. For example, primer pairs of F2 and F3 could also produce bright bands at 50 °C (Figure 2). Anyway, R3 and 53 °C would be the most optimal choice and recommended.
(2) Different mismatches resulted in diverse outcomes. For instance, distinct amplifications were displayed in F1 and F2, which differ only in the mutation of “a/c” and “a/g” at position -2 from the 3′ terminus of primer 2687-FI-S1 and 2687-FI-S2, respectively (Figure 1, Table 2). Based on Ye’s report, “a/c” belongs to a “weak” mismatch, and “a/g” belongs to a “strong” mismatch [23]. Accordingly, F1 contained two “weak” mismatches, whereas F2 included a “weak” mismatch plus a “strong” one. Because there was an additional SNP2680 (g/a, a strong mismatch) located in the middle of the primer, an over-mismatch could probably have happened in F2, resulting in no product. For F3, “a/t” belongs to the “medium” mismatch, and a weak band was appeared. In short, bright, weak, and invisible band was demonstrated in F1, F3, and F2 due to “two weak and a strong” mismatches, “a weak, a medium and a strong” mismatches, and “a weak and two strong” mismatches, respectively.
5. Conclusions
In this study, a novel tetra-primer ARMS-PCR marker for quick and reliable identification of Pi25 was developed. Compared to previous gene-targeted markers, it could promote the accuracy in the identification of genotypes or could achieve the same result with less workload and cost. The newly developed marker would be a valuable tool for genotypic assay and rice molecular breeding of blast resistance.
Author Contributions
Conceptualization, J.Z. and T.H.; methodology, Y.F. and Y.Z.; investigation, Y.F., Z.Z., D.H. and H.W.; writing—original draft preparation, Y.F.; writing—review and editing, Y.F. and Y.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Major Agricultural Science and Technology Projects (2022ZD0400202); The Agricultural Science and Technology Innovation Program (CAAS-ASTIP-2021-CNRRI); Project of the State Key Laboratory of Rice Biology and Breeding (2020ZZKT10105); Science and Technology Plan Project of Fujian Province (2021R1023009).
Data Availability Statement
Not applicable.
Acknowledgments
We thank Haiping Qiu from Zhejiang Academy of Agricultural Sciences for kindly providing LTH and its NILs.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Against the grain: Safeguarding rice from rice blast disease. Trends Biotechnol. 2009, 27, 141–150. [Google Scholar] [CrossRef]
- Khush, G.S.; Jena, K.K. Current status and future prospects for research on blast resistance in rice (Oryza sativa L.). In Advances in Genetics, Genomics and Control of Rice Blast Disease; Wang, G.-L., Valent, B., Eds.; Springer: New York, NY, USA, 2009; pp. 1–10. [Google Scholar] [CrossRef]
- Li, W.; Chern, M.; Yin, J.; Wang, J.; Chen, X. Recent advances in broad-spectrum resistance to the rice blast disease. Curr. Opin. Plant Biol. 2019, 50, 114–120. [Google Scholar] [CrossRef]
- Cobb, J.N.; Biswas, P.S.; Platten, J.D. Back to the future: Revisiting MAS as a tool for modern plant breeding. Theor. Appl. Genet. 2019, 132, 647–667. [Google Scholar] [CrossRef]
- Zhuang, J.-Y.; Ma, W.-B.; Wu, J.-L.; Chai, R.-Y.; Lu, J.; Fan, Y.-Y.; Jin, M.-Z.; Leung, H.; Zheng, K.-L. Mapping of leaf and neck blast resistance genes with resistance gene analog, RAPD and RFLP in rice. Euphytica 2002, 128, 363–370. [Google Scholar] [CrossRef]
- Wu, J.-L.; Fan, Y.-Y.; Li, D.-B.; Zheng, K.-L.; Leung, H.; Zhuang, J.-Y. Genetic control of rice blast resistance in the durably resistant cultivar Gumei 2 against multiple isolates. Theor. Appl. Genet. 2005, 111, 50–56. [Google Scholar] [CrossRef]
- Chen, J.; Shi, Y.; Liu, W.; Chai, R.; Fu, Y.; Zhuang, J.; Wu, J. A Pid3 allele from rice cultivar Gumei2 confers resistance to Magnaporthe oryzae. J. Genet. Genom. 2011, 38, 209–216. [Google Scholar] [CrossRef]
- Wang, H.-M.; Chen, J.; Shi, Y.-F.; Pan, G.; Shen, H.-C.; Wu, J.-L. Development and validation of CAPS markers for marker-assisted selection of rice blast resistance gene Pi25. Acta Agron. Sini. 2012, 38, 1960–1968. [Google Scholar] [CrossRef]
- Zhou, H.; Zhan, X.; Chai, R.; Cheng, S.; Cao, L. Breeding of rice restorer lines carrying blast resistance gene Pi25 by marker-assisted selection. Chin. J. Rice Sci. 2008, 22, 590–596, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhan, X.; Zhou, H.; Chai, R.; Zhuang, J.; Cheng, S.; Cao, L. Breeding of R8012, a rice restorer line resistant to blast and bacterial blight through marker-assisted selection. Rice Sci. 2012, 19, 29–35. [Google Scholar] [CrossRef]
- Yu, S.; Zheng, X.; Fan, T.; Huang, Y.; Chen, H.; Chen, S.; Hong, X.; Ruan, G. Breeding of PTGMS lines with blast resistance gene Pi25 by molecular marker-assisted selection. China Rice 2013, 19, 15–17, (In Chinese with English Abstract). [Google Scholar]
- Tu, S.; Zhou, P.; Zheng, Y.; Dong, R.; Zhang, S.; Huang, T.; Zheng, J. Breeding blast-resistant male sterile line of three line hybrid by Pi25 gene marker assisted selection. Mol. Plant Breed. 2015, 13, 1911–1917, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Dong, R.; Wang, H.; Dong, L.; Zhou, P.; Tu, S.; You, Q.; Liao, F.; Huang, T. Improving the rice blast resistance for a CMS line of rice Zhenda A and its hybrids using molecular marker-assisted selection. J. Plant Genet. Resour. 2017, 18, 573–586, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Liu, W.; Li, X.; Li, Y.; Pan, X.; Sheng, X.; Duan, Y. Improvement of rice blast resistance of Xiangwanxian No.13 with high quality by molecular marker-assisted selection. Mol. Plant Breed. 2017, 15, 3063–3069, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Cao, N.; Ji, Z.; Zeng, Y.; Zheng, A.; Fang, J.; Wang, S.; Wu, K.; Chen, H.; Yang, C.; Liang, Y. Improving the blast resistance of super early-season rice variety Zhongzao39. J. Plant Genet. Resour. 2020, 21, 809–818, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Mei, W.; Liu, P.; Hong, B.; Mu, H.; Lv, Q.; Sha, A.; Guo, S. Molecular identification of blast resistance genes Pi25, Pi56(t), Pit, and Pita from rice varieties. Hubei Agri. Sci. 2016, 55, 6604–6607, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Xu, H.; Zhou, L.; Jiang, Y.; Li, S.; Zha, W.; Liu, K.; Chen, Z.; Yang, G.; Li, P.; Fang, G.; et al. Distribution of some disease and insect resistance genes and quality related genes in rice core germplasm resources. Mol. Plant Breed. 2018, 16, 1121–1126, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Wang, Y.; Chen, X.; Yin, H.; Wang, Y.; Yang, R.; Zang, Z.; Wang, S. Molecular detection of rice blast resistance gene in the main rice germplasms in Henan Province. Mol. Plant Breed. 2018, 16, 3203–3212, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Qi, C.; Xu, X.; Ma, J.; Wang, S.; Tian, P.; Meng, L.; Yan, W.; Zhao, Z.; Wang, J.; Wang, J.; et al. Distribution of blast resistance genes Pib, Pita, Pi5, Pi25 and Pi54 in mini-core collection of Chinese rice germplasm. J. Plant Genet. Resour. 2019, 20, 1240–1246+1254, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, Q.; Zhao, Y.; Liu, R.; Fu, Z.; Li, X.; Sun, Y.; Jin, X. Evaluation of rice blast resistance and genetic structure analysis of rice germplasm in Heilongjiang Province. Sci. Agri. Sini. 2022, 55, 625–640, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zhou, L.; Cai, H.; Dai, F.; Xu, H.; Liu, K.; Zha, W.; Li, S.; Yang, G.; Chen, Z.; Li, P.; et al. Development and application of a functional SNP marker of the blast resistant gene Pi25 in rice. Mol. Plant Breed. 2016, 14, 2680–2685, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Ye, S.; Dhillon, S.; Ke, X.; Collins, A.R.; Day, I.N. An efficient procedure for genotyping single nucleotide polymorphisms. Nucleic Acids Res. 2001, 29, E88. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Qiu, H.; Chai, R.; Mao, X.; Wang, Y.; Sun, G. Distribution and evaluation of 6 blast genes in major cultivated rice varieties in Zhejiang. Fujian J. Agri. Sci. 2019, 34, 214–222, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Zheng, K.; Huang, N.; Bennett, J.; Khush, G.S. PCR-Based Marker-Assisted Selection in Rice Breeding; IRRI Discussion Paper Series No.12; International Rice Research Institute: Los Baños, Philippines, 1995. [Google Scholar]
- Zhou, B.; Qu, S.; Liu, G.; Dolan, M.; Sakai, H.; Lu, G.; Bellizzi, M.; Wang, G. The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea. Mol. Plant Microbe Interact. 2006, 19, 1216–1228. [Google Scholar] [CrossRef]
- Peng, M.; Lin, X.; Xiang, X.; Ren, H.; Fan, X.; Chen, K. Characterization and evaluation of transgenic rice pyramided with the Pi genes Pib, Pi25 and Pi54. Rice 2021, 14, 78. [Google Scholar] [CrossRef]
- Iyer-Pascuzzi, A.S.; McCouch, S.R. Functional marker for xa5-mediated resistance in rice (Oryza sativa L.). Mol. Breed. 2007, 19, 291–296. [Google Scholar] [CrossRef]
- Medrano, R.F.; de Oliveira, C.A. Guidelines for the tetra-primer ARMS-PCR technique development. Mol. Biotechnol. 2014, 56, 599–608. [Google Scholar] [CrossRef]
- Pereira, A.; Tcacenco, F.A.; Klabunde, G.H.F.; de Andrade, A. Detecting acetyl-coenzyme a carboxylase resistance gene in rice (Oryza sativa L.). Mol. Biol. Rep. 2019, 46, 6271–6276. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, A.; Wang, F.; Wang, J.; Bi, J.; Kong, D.; Zhang, F.; Luo, L.; Liu, G.; Yu, X. Development and validation of a PCR-based functional marker system for identifying the low amylose content-associated gene Wx hp in rice. Breed. Sci. 2019, 69, 702–706. [Google Scholar] [CrossRef]
- Mao, T.; Zhu, M.; Ahmad, S.; Ye, G.; Sheng, Z.; Hu, S.; Jiao, G.; Xie, L.; Tang, S.; Wei, X.; et al. Superior japonica rice variety YJ144 with improved rice blast resistance, yield, and quality achieved using molecular design and multiple breeding strategies. Mol. Breed. 2021, 41, 65. [Google Scholar] [CrossRef]
- Mesrian Tanha, H.; Mojtabavi Naeini, M.; Rahgozar, S.; Rasa, S.M.; Vallian, S. Modified tetra-primer ARMS PCR as a single-nucleotide polymorphism genotyping tool. Genet. Test. Mol. Biomark. 2015, 19, 156–161. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Principle of primer designation of tetra-primer ARMS-PCR marker of rice blast resistance gene Pi25 based on two SNPs. (A) Resistance-specific fragment generated by forward inner primer-R containing both “g” at the SNP site of 2680 and 2687; (B) Susceptibility-specific fragment generated by forward inner primer-S containing both “a” at the SNP site of 2680 and 2687. Note: 2680 and 2687 mean the SNP position from the start codon of the Pi25 gene; The nucleotide in red refers to that of the resistant allele, and blue one refers to that of the susceptible allele; Green asterisk stands for mismatched nucleotide introduced into the primer; Rectangle in red, blue and black presents resistance-specific, susceptibility-specific and common PCR product, respectively.
Figure 1.
Principle of primer designation of tetra-primer ARMS-PCR marker of rice blast resistance gene Pi25 based on two SNPs. (A) Resistance-specific fragment generated by forward inner primer-R containing both “g” at the SNP site of 2680 and 2687; (B) Susceptibility-specific fragment generated by forward inner primer-S containing both “a” at the SNP site of 2680 and 2687. Note: 2680 and 2687 mean the SNP position from the start codon of the Pi25 gene; The nucleotide in red refers to that of the resistant allele, and blue one refers to that of the susceptible allele; Green asterisk stands for mismatched nucleotide introduced into the primer; Rectangle in red, blue and black presents resistance-specific, susceptibility-specific and common PCR product, respectively.

Figure 2.
The products were amplified by two sets of primer pairs using gradient annealing temperatures. Note: M, DNA Marker; 1, Gumei 2; 2, Zhong156; 3, Tetep; R1, R2 and R3, three primer pairs composed of 2687-FO, 2687-RO, 2687-FI-R and three 2687-RI-S primers with different mismatches, respectively; F1, F2 and F3, three primer pairs composed of 2687-FO, 2687-RO, 2687-RI-R and three 2687-FI-S primers with different mismatches, respectively.
Figure 2.
The products were amplified by two sets of primer pairs using gradient annealing temperatures. Note: M, DNA Marker; 1, Gumei 2; 2, Zhong156; 3, Tetep; R1, R2 and R3, three primer pairs composed of 2687-FO, 2687-RO, 2687-FI-R and three 2687-RI-S primers with different mismatches, respectively; F1, F2 and F3, three primer pairs composed of 2687-FO, 2687-RO, 2687-RI-R and three 2687-FI-S primers with different mismatches, respectively.

Figure 3.
The amplification of Pi25-2687R3 and Pi25-2566. Note: M, DNA Marker; 1, Gumei 2; 2, Zhong156; 3, Tetep; 4, Zhongjian 100; 5, Gumei 4; 6, Lijiangxintuanheigu (LTH); 7, IRBLz-Fu; 8, IRBLzt-T; 9, IRBLz5-CA; 10, IRBL9-W.
Figure 3.
The amplification of Pi25-2687R3 and Pi25-2566. Note: M, DNA Marker; 1, Gumei 2; 2, Zhong156; 3, Tetep; 4, Zhongjian 100; 5, Gumei 4; 6, Lijiangxintuanheigu (LTH); 7, IRBLz-Fu; 8, IRBLzt-T; 9, IRBLz5-CA; 10, IRBL9-W.
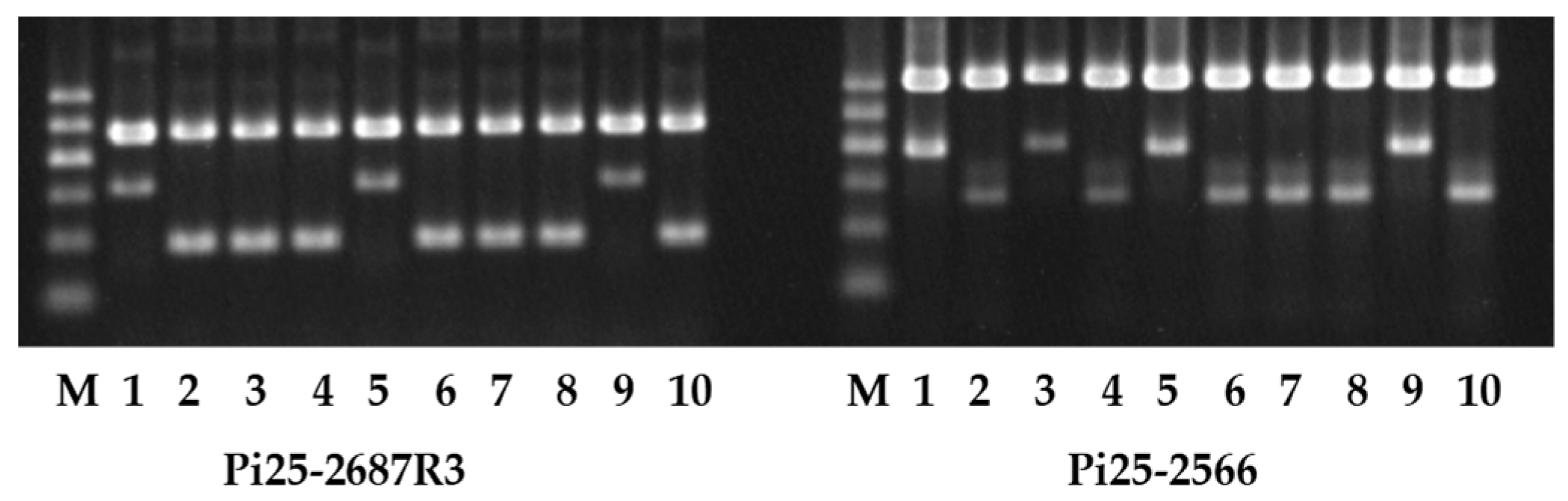
Figure 4.
Genotyping of 17 breeding lines/varieties using SNP2687R3. Note: M, DNA Marker; +, Gumei 2; −, Zhong156; 1~12, restorer lines; 13~17, rice hybrid varieties.
Figure 4.
Genotyping of 17 breeding lines/varieties using SNP2687R3. Note: M, DNA Marker; +, Gumei 2; −, Zhong156; 1~12, restorer lines; 13~17, rice hybrid varieties.

Figure 5.
Genotyping of 94 rice landraces using newly developed and published markers. Note: M, DNA Marker; +, Gumei 2; −, Zhong156; 1~94, rice landraces.
Figure 5.
Genotyping of 94 rice landraces using newly developed and published markers. Note: M, DNA Marker; +, Gumei 2; −, Zhong156; 1~94, rice landraces.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Information on rice germplasm was used in this study.
| No. | Accession Name | Country | Pi25-2687R3 | CAP3/Hpy 99I | Pi25-2566 |
|---|---|---|---|---|---|
| 1 | AKHNI SAIL | Bangladesh | − | − | − |
| 2 | AUS 43 | Bangladesh | − | − | − |
| 3 | BERI | Bangladesh | − | − | − |
| 4 | BR 601-3-3-2-4 | Bangladesh | − | − | − |
| 5 | CHAKULA | Bangladesh | − | − | − |
| 6 | RANGPUR | Bangladesh | − | − | − |
| 7 | MA HNAN THU KA | Burma | − | − | + |
| 8 | X72-7-1 | Burma | − | − | − |
| 9 | IB 28 | Burundi | − | − | − |
| 10 | IB 42 | Burundi | − | − | − |
| 11 | NEANG CHHOUK | Cambodia | − | − | − |
| 12 | NEANG SAN | Cambodia | − | − | − |
| 13 | 5173 | Colombia | + | + | + |
| 14 | RT 1031-69 | Congo | − | − | − |
| 15 | AMISTAD 82-8 | Cuba | − | − | − |
| 16 | C.CORTO 1 | Cuba | − | − | + |
| 17 | GU2725 | Cuba | − | − | − |
| 18 | SELECCION VG-5 | Cuba | − | − | − |
| 19 | GIZA 181 | Egypt | − | − | − |
| 20 | CSR 11 | India | − | − | − |
| 21 | CSR 12 | India | − | − | − |
| 22 | CSR 13 | India | − | − | − |
| 23 | CSR 5 | India | − | − | − |
| 24 | CSR 9 | India | − | − | − |
| 25 | DULAR | India | − | − | − |
| 26 | HARWA | India | − | − | − |
| 27 | HIMDHAN | India | − | − | − |
| 28 | HPA 74 | India | − | − | − |
| 29 | JC 195 | India | − | − | − |
| 30 | JW 42 | India | − | − | − |
| 31 | JW 60 | India | − | − | − |
| 32 | K 428-25 | India | − | − | − |
| 33 | KAJARU | India | − | − | − |
| 34 | KALING A2 | India | − | − | − |
| 35 | NDR 4012 | India | − | − | − |
| 36 | NDR 89 | India | − | − | − |
| 37 | OR 79-21 | India | − | − | − |
| 38 | PR32-PD47-PD4 | India | − | − | − |
| 39 | RP 1017-76-1-3-2 | India | − | − | − |
| 40 | T 23 | India | − | − | − |
| 41 | TM 10265 | India | − | − | − |
| 42 | VL DHAN 16 | India | − | − | − |
| 43 | B 6397F-MR-7-5M-1-1 | Indonesia | − | − | − |
| 44 | BAHBUTONG | Indonesia | − | − | − |
| 45 | BATANG PANE | Indonesia | − | − | − |
| 46 | BOYO | Indonesia | − | − | − |
| 47 | CIMANUK | Indonesia | − | − | − |
| 48 | CIPUNEGARA | Indonesia | − | − | − |
| 49 | PADI SEGUTUK | Indonesia | − | − | + |
| 50 | DOMSIAH 138 | Iran | − | − | − |
| 51 | HABATAKI | Japan | − | − | − |
| 52 | SANJIANG | Japan | − | − | − |
| 53 | SHINKWANG | Korea | − | − | − |
| 54 | JHALI | Nepal | − | − | − |
| 55 | ITA 304 | Nigeria | − | − | − |
| 56 | MILYANG 70 | North Korea | − | − | − |
| 57 | SHUIYUAN299 | North Korea | − | − | − |
| 58 | 368 | Pakistan | − | − | − |
| 59 | BASMATI 370 | Pakistan | − | − | − |
| 60 | BASMATI 685 | Pakistan | − | − | − |
| 61 | GANJAY | Pakistan | − | − | − |
| 62 | JIJAI NIKI | Pakistan | − | − | − |
| 63 | KHARA GANJA | Pakistan | − | − | + |
| 64 | RATRIA | Pakistan | − | − | − |
| 65 | TAK RATIA | Pakistan | − | − | + |
| 66 | PANAMA 1537 | Panama | + | + | + |
| 67 | 75-1-120 | Philippines | − | − | − |
| 68 | 97A-M54 | Philippines | − | − | − |
| 69 | A1452 | Philippines | − | − | − |
| 70 | BINICOL | Philippines | − | − | − |
| 71 | PIRURUTONG | Philippines | − | − | − |
| 72 | PR 23631-98 | Philippines | − | − | − |
| 73 | PRATAO PRECOCE | Philippines | − | − | − |
| 74 | SINABA | Philippines | − | − | − |
| 75 | SINANDOMENG | Philippines | − | − | − |
| 76 | BATHKIRIEL | Sri Lanka | − | − | − |
| 77 | BG 1165-1 | Sri Lanka | − | − | − |
| 78 | BG 170 | Sri Lanka | − | − | − |
| 79 | BG 915 | Sri Lanka | − | − | − |
| 80 | KATAHATA HAMBA | Sri Lanka | − | − | − |
| 81 | MALKORA | Sri Lanka | − | − | − |
| 82 | MURUNGAWEE | Sri Lanka | − | − | − |
| 83 | SINNA SIVAPPU | Sri Lanka | − | − | − |
| 84 | NIAW SANPAH TAWNG | Thailand | − | − | − |
| 85 | XIANLUOSICHI | Thailand | − | − | − |
| 86 | DAWN CI9534 | USA | − | − | − |
| 87 | NOVA 66 CI9481 | USA | − | − | − |
| 88 | BAU HUONG DOONG | Vietnam | − | − | − |
| 89 | CHAH NONG NGHE AN | Vietnam | − | − | + |
| 90 | CHIEM BAC | Vietnam | − | − | − |
| 91 | LAI TRANG | Vietnam | − | − | − |
| 92 | QUCM | Vietnam | − | − | − |
| 93 | RE BAU | Vietnam | − | − | + |
| 94 | RE CHANH | Vietnam | − | − | + |
Note: +, resistant allele; −, susceptible allele.
Table 2.
Primers of tetra-primer ARMS-PCR markers developed in this study.
| Type | Name | Sequence (5′-3′) |
|---|---|---|
| Forward outer primer | 2687-FO | CACACCTGAATGAAATTAAGATGACA |
| Reverse outer primer | 2687-RO | ATATACAATATTGAGGGTATGGAAC |
| Forward inner primer-R | 2687-FI-R | GCTTGTGGATAGAGTCCTTCG |
| Reverse inner primer-S | 2687-RI-S1 | CAAATCATTCGCTCTTTTCCCT |
| 2687-RI-S2 | CAAATCATTCGCTCTTTTGGCT | |
| 2687-RI-S3 | TCACAAATCATTCGCTCTTTTAACT | |
| Forward inner primer-S | 2687-FI-S1 | GAGCTTGTGGATAGAATCCTCCA |
| 2687-FI-S2 | GAGCTTGTGGATAGAATCCTGCA | |
| 2687-FI-S3 | GAGCTTGTGGATAGAATCCTTCA | |
| Reverse inner primer-R | 2687-RI-R | TCACAAATCATTCGCTCTTTTAACC |
Note: Red letter represents nucleotide at the SNP site of 2680 and 2687; Green letter represents mismatched nucleotide.
Table 3.
Phenotype of rice breeding lines/varieties genotyped with Pi25-2687R3.
| No. in Figure 4 | Line/Variety | Type | Genotype | Phenotype |
|---|---|---|---|---|
| 1 | R173 | Restorer line | + | R |
| 2 | R1909 | Restorer line | + | R |
| 3 | R1913 | Restorer line | + | R |
| 4 | R1925 | Restorer line | + | MR |
| 5 | R1927 | Restorer line | + | R |
| 6 | R1932 | Restorer line | + | MR |
| 7 | R1933 | Restorer line | + | MR |
| 8 | R1934 | Restorer line | + | R |
| 9 | R1937 | Restorer line | − | S |
| 10 | ZP174 | Restorer line | + | R |
| 11 | ZP175 | Restorer line | − | MS |
| 12 | ZP178 | Restorer line | + | R |
| 13 | Maoxiangyou 61 | Hybrid variety | +/− | R |
| 14 | Mixiangyou 153 | Hybrid variety | +/− | R |
| 15 | Huangyou 153 | Hybrid variety | +/− | R |
| 16 | Zhongxiangyou 173 | Hybrid variety | +/− | R |
| 17 | Zhiliangyou 173 | Hybrid variety | +/− | R |
Note: +, homozygous resistant allele; −, homozygous susceptible allele; +/−, heterozygous allele; R, resistance; MR, moderate resistance; MS, moderate susceptibility; S, susceptibility.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Fan, Y.; Zhang, Z.; Huang, D.; Huang, T.; Wang, H.; Zhuang, J.; Zhu, Y. A Novel PCR-Based Functional Marker of Rice Blast Resistance Gene Pi25. Agriculture 2023, 13, 1926. https://doi.org/10.3390/agriculture13101926
AMA Style
Fan Y, Zhang Z, Huang D, Huang T, Wang H, Zhuang J, Zhu Y. A Novel PCR-Based Functional Marker of Rice Blast Resistance Gene Pi25. Agriculture. 2023; 13(10):1926. https://doi.org/10.3390/agriculture13101926
Chicago/Turabian StyleFan, Yeyang, Zhenhua Zhang, Derun Huang, Tingxu Huang, Hongfei Wang, Jieyun Zhuang, and Yujun Zhu. 2023. "A Novel PCR-Based Functional Marker of Rice Blast Resistance Gene Pi25" Agriculture 13, no. 10: 1926. https://doi.org/10.3390/agriculture13101926
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.