
Nicotinic Acid-Mediated Modulation of Metastasis-Associated Protein 1 Methylation and Inflammation in Brain Arteriovenous Malformation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Biochemical Analysis
2.2. Cerebral Artery CT Angiography
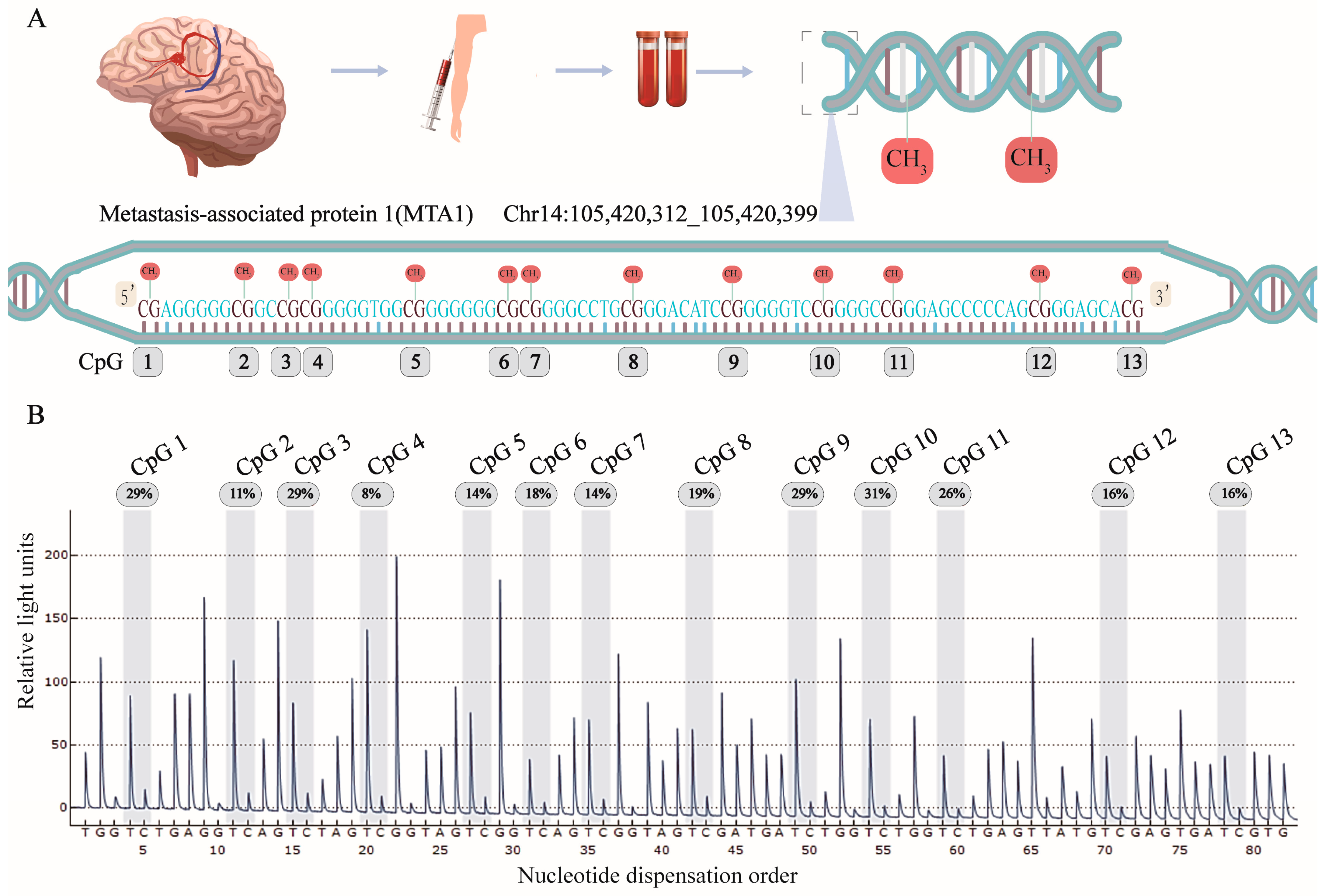
2.3. Pyrosequencing
2.4. Quantitative Real-Time Polymerase Chain Reactions (RT-qPCR)
Western Blot Analysis
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Detection of Interleukin Using Cytometric Bead Array
2.7. Cell Culture and Drug Treatment
2.8. Transfection
2.9. Statistical Analyses
3. Results
3.1. Characteristics of Study Participants
3.2. Cerebral Artery CT Angiography
3.3. mRNA and Protein Expression
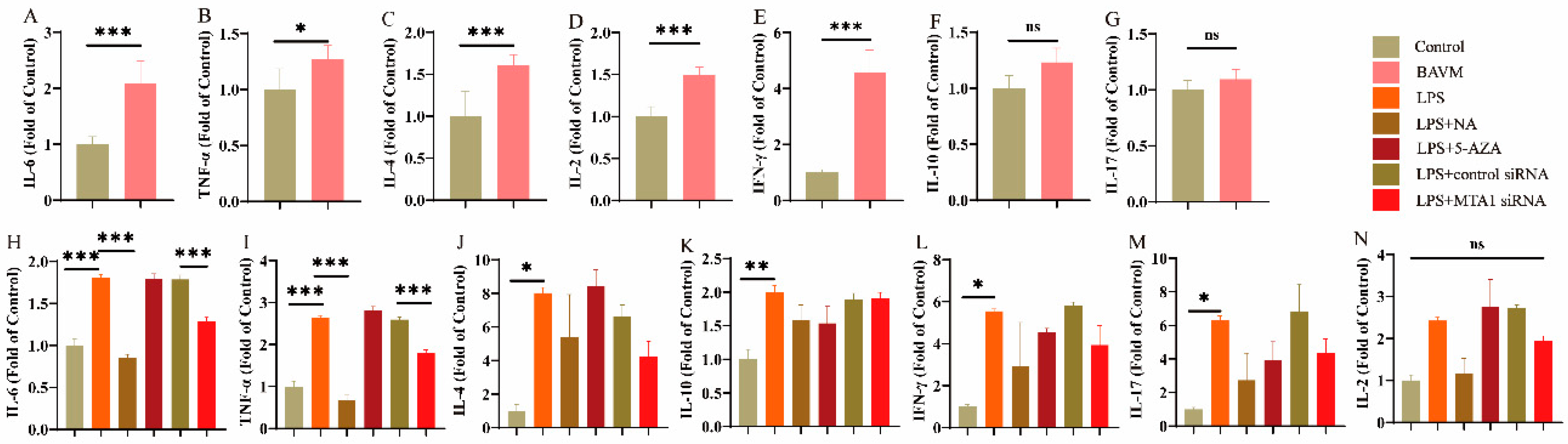
3.4. Cytokine Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Friedlander, R.M. Clinical practice. Arteriovenous malformations of the brain. N. Engl. J. Med. 2007, 356, 2704–2712. [Google Scholar] [CrossRef]
- Chen, G.-Z.; Ke, Y.; Qin, K.; Dong, M.-Q.; Zeng, S.-J.; Lin, X.-F.; Zhan, S.-Q.; Tang, K.; Peng, C.; Ding, X.-W.; et al. Analysis of the Expression of Angioarchitecture-related Factors in Patients with Cerebral Arteriovenous Malformation. Chin. Med. J. 2017, 130, 2465–2472. [Google Scholar] [CrossRef]
- Chen, Y.; Pawlikowska, L.; Yao, J.S.; Shen, F.; Zhai, W.; Achrol, A.S.; Lawton, M.T.; Kwok, P.-Y.; Yang, G.-Y.; Young, W.L. Interleukin-6 involvement in brain arteriovenous malformations. Ann. Neurol. 2006, 59, 72–80. [Google Scholar] [CrossRef]
- Chen, Y.; Zhu, W.; Bollen, A.W.; Lawton, M.T.; Barbaro, N.M.; Dowd, C.F.; Hashimoto, T.; Yang, G.-Y.; Young, W.L. Evidence of inflammatory cell involvement in brain arteriovenous malformations. Neurosurgery 2008, 62, 1340. [Google Scholar] [CrossRef]
- Guo, Y.; Tihan, T.; Kim, H.; Hess, C.; Lawton, M.T.; Young, W.L.; Zhao, Y.; Su, H. Distinctive distribution of lymphocytes in unruptured and previously untreated brain arteriovenous malformation. Neuroimmunol. Neuroinflamm. 2014, 1, 147–152. [Google Scholar]
- Jeltsch, A. Beyond Watson and Crick: DNA methylation and molecular enzymology of DNA methyltransferases. Chembiochem 2002, 3, 274–293. [Google Scholar] [CrossRef]
- Guay, S.-P.; Brisson, D.; Munger, J.; Lamarche, B.; Gaudet, D.; Bouchard, L. ABCA1 gene promoter DNA methylation is associated with HDL particle profile and coronary artery disease in familial hypercholesterolemia. Epigenetics 2012, 7, 464–472. [Google Scholar] [CrossRef]
- Caranci, F.; Briganti, F.; Cirillo, L.; Leonardi, M.; Muto, M. Epidemiology and genetics of intracranial aneurysms. Eur. J. Radiol. 2013, 82, 1598–1605. [Google Scholar] [CrossRef]
- Thomas, J.M.; Surendran, S.; Abraham, M.; Rajavelu, A.; Kartha, C.C. Genetic and epigenetic mechanisms in the development of arteriovenous malformations in the brain. Clin. Epigenet. 2016, 8, 78. [Google Scholar] [CrossRef]
- Thomas, J.M.; Sasankan, D.; Abraham, M.; Surendran, S.; Kartha, C.C.; Rajavelu, A. DNA methylation signatures on vascular differentiation genes are aberrant in vessels of human cerebral arteriovenous malformation nidus. Clin. Epigenet. 2022, 14, 127. [Google Scholar] [CrossRef]
- Joo, J.E.; Hiden, U.; Lassance, L.; Gordon, L.; Martino, D.J.; Desoye, G.; Saffery, R. Variable promoter methylation contributes to differential expression of key genes in human placenta-derived venous and arterial endothelial cells. BMC Genom. 2013, 14, 475. [Google Scholar] [CrossRef]
- Wang, T.; Li, W.; Huang, H.; Wang, C. Metastasis-Associated 1 (MTA1) Gene Expression Promotes Angiogenesis in Mouse Xenografts from Human Non-Small Cell Lung Cancer (NSCLC) Cells. Med. Sci. Monit. 2019, 25, 484–491. [Google Scholar] [CrossRef]
- Kai, L.; Wang, J.; Ivanovic, M.; Chung, Y.-T.; Laskin, W.B.; Schulze-Hoepfner, F.; Mirochnik, Y.; Satcher, R.L.; Levenson, A.S. Targeting prostate cancer angiogenesis through metastasis-associated protein 1 (MTA1). Prostate 2011, 71, 268–280. [Google Scholar] [CrossRef]
- Yoo, Y.-G.; Kong, G.; Lee, M.-O. Metastasis-associated protein 1 enhances stability of hypoxia-inducible factor-1alpha protein by recruiting histone deacetylase 1. EMBO J. 2006, 25, 1231–1241. [Google Scholar] [CrossRef]
- Pakala, S.B.; Reddy, S.D.N.; Bui-Nguyen, T.M.; Rangparia, S.S.; Bommana, A.; Kumar, R. MTA1 coregulator regulates LPS response via MyD88-dependent signaling. J. Biol. Chem. 2010, 285, 32787–32792. [Google Scholar] [CrossRef]
- Chen, J.-K.; Wang, W.-C.; Zang, L.; Zhao, J.; Li, W.; Jiang, T. Repression of a chromatin modifier aggravates lipopolysaccharide-induced acute lung injury in mouse. Biochem. Biophys. Res. Commun. 2016, 471, 515–521. [Google Scholar] [CrossRef]
- Wang, H.; Dong, B.-W.; Zheng, Z.-H.; Wu, Z.-B.; Li, W.; Ding, J. Metastasis-associated protein 1 (MTA1) signaling in rheumatoid synovium: Regulation of inflammatory response and cytokine-mediated production of prostaglandin E2 (PGE2). Biochem. Biophys. Res. Commun. 2016, 473, 442–448. [Google Scholar] [CrossRef]
- Li, C.; Wang, H.; Lin, F.; Li, H.; Wen, T.; Qian, H.; Zhan, Q. Bioinformatic exploration of MTA1-regulated gene networks in colon cancer. Front. Med. 2016, 10, 178–182. [Google Scholar] [CrossRef]
- Li, L.; Liu, J.; Xue, H.; Li, C.; Liu, Q.; Zhou, Y.; Wang, T.; Wang, H.; Qian, H.; Wen, T. A TGF-β-MTA1-SOX4-EZH2 signaling axis drives epithelial-mesenchymal transition in tumor metastasis. Oncogene 2020, 39, 2125–2139. [Google Scholar] [CrossRef]
- Winkler, E.A.; Pacult, M.A.; Catapano, J.S.; Scherschinski, L.; Srinivasan, V.M.; Graffeo, C.S.; Oh, S.P.; Lawton, M.T. Emerging pathogenic mechanisms in human brain arteriovenous malformations: A contemporary review in the multiomics era. Neurosurg. Focus. 2022, 53, E2. [Google Scholar] [CrossRef]
- Hashimoto, T.; Mesa-Tejada, R.; Quick, C.M.; Bollen, A.W.; Joshi, S.; Pile-Spellman, J.; Lawton, M.T.; Young, W.L. Evidence of increased endothelial cell turnover in brain arteriovenous malformations. Neurosurgery 2001, 49, 124–132. [Google Scholar] [PubMed]
- Sure, U.; Butz, N.; Schlegel, J.; Siegel, A.M.; Wakat, J.P.; Mennel, H.D.; Bien, S.; Bertalanffy, H. Endothelial proliferation, neoangiogenesis, and potential de novo generation of cerebrovascular malformations. J. Neurosurg. 2001, 94, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Barros, K.V.; Carvalho, P.O.; Cassulino, A.P.; Andrade, I.; West, A.L.; Miles, E.A.; Calder, P.C.; Silveira, V.L.F. Fatty acids in plasma, white and red blood cells, and tissues after oral or intravenous administration of fish oil in rats. Clin. Nutr. 2013, 32, 993–998. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhang, J.; Zhou, C.; Gong, F.; Zhu, X.; Pan, X.; Sun, J.; Gao, X.; Huang, Y. DNA Methylation of Patatin-Like Phospholipase Domain-Containing Protein 6 Gene Contributes to the Risk of Intracranial Aneurysm in Males. Front. Aging Neurosci. 2022, 14, 885680. [Google Scholar] [CrossRef]
- Huang, X.; Qiu, W.; Pan, Y.; Li, J.; Chen, Z.; Zhang, K.; Luo, Y.; Wu, B.; Xu, W. Exosomes from LPS-Stimulated hDPSCs Activated the Angiogenic Potential of HUVECs In Vitro. Stem Cells Int. 2021, 2021, 6685307. [Google Scholar] [CrossRef]
- Huang, Z.; Kraus, V.B. Does lipopolysaccharide-mediated inflammation have a role in OA? Nat. Rev. Rheumatol. 2016, 12, 123–129. [Google Scholar] [CrossRef]
- Schandelmaier, S.; Briel, M.; Saccilotto, R.; Olu, K.K.; Arpagaus, A.; Hemkens, L.G.; Nordmann, A.J. Niacin for primary and secondary prevention of cardiovascular events. Cochrane Database Syst. Rev. 2017, 6, CD009744. [Google Scholar] [CrossRef]
- Zandi-Nejad, K.; Takakura, A.; Jurewicz, M.; Chandraker, A.K.; Offermanns, S.; Mount, D.; Abdi, R. The role of HCA2 (GPR109A) in regulating macrophage function. FASEB J. 2013, 27, 4366–4374. [Google Scholar] [CrossRef]
- Spetzler, R.F.; Martin, N.A. A proposed grading system for arteriovenous malformations. J. Neurosurg. 1986, 65, 476–483. [Google Scholar] [CrossRef]
- Cullell, N.; Soriano-Tárraga, C.; Gallego-Fábrega, C.; Cárcel-Márquez, J.; Torres-Águila, N.P.; Muiño, E.; Lledós, M.; Llucià-Carol, L.; Esteller, M.; Castro de Moura, M.; et al. DNA Methylation and Ischemic Stroke Risk: An Epigenome-Wide Association Study. Thromb. Haemost. 2022, 122, 1767–1778. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.; Zhou, S.; Nie, S.; Lin, Z.; Zhou, C.; Sun, J.; Gao, X.; Huang, Y. Methylation of the CDKN2A Gene Increases the Risk of Brain Arteriovenous Malformations. J. Mol. Neurosci. 2019, 69, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Jabari, S.; Kobow, K.; Pieper, T.; Hartlieb, T.; Kudernatsch, M.; Polster, T.; Bien, C.G.; Kalbhenn, T.; Simon, M.; Hamer, H.; et al. DNA methylation-based classification of malformations of cortical development in the human brain. Acta Neuropathol. 2022, 143, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Gao, X.; Sun, J.; Lin, Z.; Huang, Y. DNA Methylation of the PDGFD Gene Promoter Increases the Risk for Intracranial Aneurysms and Brain Arteriovenous Malformations. DNA Cell Biol. 2017, 36, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Feng, B.; Luan, L.; Fang, Z.; Zhao, G. MeCP2 inhibits ischemic neuronal injury by enhancing methylation of the FOXO3a promoter to repress the SPRY2-ZEB1 axis. Exp. Mol. Med. 2022, 54, 1076–1085. [Google Scholar] [CrossRef]
- Zhu, Y.; Wloch, A.; Wu, Q.; Peters, C.; Pagenstecher, A.; Bertalanffy, H.; Sure, U. Involvement of PTEN promoter methylation in cerebral cavernous malformations. Stroke 2009, 40, 820–826. [Google Scholar] [CrossRef]
- Pearce, M.S.; McConnell, J.C.; Potter, C.; Barrett, L.M.; Parker, L.; Mathers, J.C.; Relton, C.L. Global LINE-1 DNA methylation is associated with blood glycaemic and lipid profiles. Int. J. Epidemiol. 2012, 41, 210–217. [Google Scholar] [CrossRef]
- Cash, H.L.; McGarvey, S.T.; Houseman, E.A.; Marsit, C.J.; Hawley, N.L.; Lambert-Messerlian, G.M.; Viali, S.; Tuitele, J.; Kelsey, K.T. Cardiovascular disease risk factors and DNA methylation at the LINE-1 repeat region in peripheral blood from Samoan Islanders. Epigenetics 2011, 6, 1257–1264. [Google Scholar] [CrossRef]
- Kraler, S.; Wenzl, F.A.; Akhmedov, A.; Blaser, M.C.; Aikawa, E.; Lüscher, T.F. ApoA-I mimetics improve aortic stenosis-associated left-ventricular diastolic dysfunction but fail to benefit rabbit models with normal aortic valves. Int. J. Cardiol. 2021, 332, 159–161. [Google Scholar] [CrossRef]
- Federici, A.B. HDL/ApoA-I: Role in VWF-dependent thrombosis. Blood 2016, 127, 526–528. [Google Scholar] [CrossRef]
- Guo, Y.; Huang, B.; Li, R.; Li, J.; Tian, S.; Peng, C.; Dong, W. Low APOA-1 Expression in Hepatocellular Carcinoma Patients Is Associated With DNA Methylation and Poor Overall Survival. Front. Genet. 2021, 12, 760744. [Google Scholar] [CrossRef]
- Winkler, E.A.; Kim, C.N.; Ross, J.M.; Garcia, J.H.; Gil, E.; Oh, I.; Chen, L.Q.; Wu, D.; Catapano, J.S.; Raygor, K.; et al. A single-cell atlas of the normal and malformed human brain vasculature. Science 2022, 375, eabi7377. [Google Scholar] [CrossRef] [PubMed]
- Mouchtouris, N.; Jabbour, P.M.; Starke, R.M.; Hasan, D.M.; Zanaty, M.; Theofanis, T.; Ding, D.; Tjoumakaris, S.I.; Dumont, A.S.; Ghobrial, G.M.; et al. Biology of cerebral arteriovenous malformations with a focus on inflammation. J. Cereb. Blood Flow. Metab. 2015, 35, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Han, Z.; Degos, V.; Shen, F.; Choi, E.-J.; Sun, Z.; Kang, S.; Wong, M.; Zhu, W.; Zhan, L.; et al. Persistent infiltration and pro-inflammatory differentiation of monocytes cause unresolved inflammation in brain arteriovenous malformation. Angiogenesis 2016, 19, 451–461. [Google Scholar] [CrossRef]
- Wülfroth, P.; Grünwald, J. Differential effect of nicotinic acid derivatives on smooth muscle and endothelial cell proliferation. Basic. Res. Cardiol. 1989, 84, 291–297. [Google Scholar] [CrossRef]
- Ghanta, K.S.; Pakala, S.B.; Reddy, S.D.N.; Li, D.-Q.; Nair, S.S.; Kumar, R. MTA1 coregulation of transglutaminase 2 expression and function during inflammatory response. J. Biol. Chem. 2011, 286, 7132–7138. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Shen, G.; Zhang, Y.; Moustafa, A.A.; Ge, D.; You, Z. Interleukin-17 Promotes Migration and Invasion of Human Cancer Cells Through Upregulation of MTA1 Expression. Front. Oncol. 2019, 9, 546. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Hao, Q.; Chen, Y.; Li, Z.; Zhang, H.; Yuan, K.; Li, R.; Li, R.; Zhao, Y.; Wang, K.; et al. mTOR-FABP4 signal is activated in brain arteriovenous malformations in humans. J. Mol. Med. 2022, 100, 1287–1297. [Google Scholar] [CrossRef]
- Li, F.; Shao, X.; Liu, D.; Jiao, X.; Yang, X.; Yang, W.; Liu, X. Vascular Disruptive Hydrogel Platform for Enhanced Chemotherapy and Anti-Angiogenesis through Alleviation of Immune Surveillance. Pharmaceutics 2022, 14, 1809. [Google Scholar] [CrossRef]
- Brüning, A.; Makovitzky, J.; Gingelmaier, A.; Friese, K.; Mylonas, I. The metastasis-associated genes MTA1 and MTA3 are abundantly expressed in human placenta and chorionic carcinoma cells. Histochem. Cell Biol. 2009, 132, 33–38. [Google Scholar] [CrossRef]
- Okamoto, M.; Matsuda, H.; Joetham, A.; Lucas, J.J.; Domenico, J.; Yasutomo, K.; Takeda, K.; Gelfand, E.W. Jagged1 on dendritic cells and Notch on CD4+ T cells initiate lung allergic responsiveness by inducing IL-4 production. J. Immunol. 2009, 183, 2995–3003. [Google Scholar] [CrossRef]
- Jabbour, M.N.; Elder, J.B.; Samuelson, C.G.; Khashabi, S.; Hofman, F.M.; Giannotta, S.L.; Liu, C.Y. Aberrant angiogenic characteristics of human brain arteriovenous malformation endothelial cells. Neurosurgery 2009, 64, 139–148. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | BAVM (n = 74) | Control (n = 74) | t/x | p |
|---|---|---|---|---|
| Age (year) | 49.70 ± 12.18 | 52.99 ± 8.35 | 1.913 | 0.058 |
| Sex (male, n) | 37 | 37 | 0.020 | 0.888 |
| Smoking (n) | 18 | 11 | 0.970 | 0.325 |
| Drinking (n) | 11 | 12 | 0.040 | 0.841 |
| TG (mmol/L) | 1.40 ± 0.79 | 1.50 ± 0.68 | 0.825 | 0.410 |
| TC (mmol/L) | 4.66 ± 0.96 | 4.74 ± 0.94 | 0.526 | 0.600 |
| HDL (mmol/L) | 1.15 ± 0.28 | 1.23 ± 0.27 | 1.852 | 0.066 |
| LDL (mmol/L) | 2.96 ± 0.64 | 3.03 ± 0.66 | 0.583 | 0.561 |
| ApoA (g/L) | 1.18 ± 0.24 | 1.31 ± 0.26 | 3.077 | 0.002 |
| ApoB (g/L) | 0.89 ± 0.22 | 0.92 ± 0.19 | 0.843 | 0.401 |
| Lpa (mg/dL) | 26.96 ± 26.05 | 19.59 ± 17.15 | 2.032 | 0.044 |
| ApoE (g/L) | 51.63 ± 16.54 | 46.33 ± 15.48 | 2.012 | 0.046 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; Zhou, S.; Hu, Z.; Gong, F.; Zhang, J.; Zhou, C.; Lan, W.; Gao, X.; Huang, Y. Nicotinic Acid-Mediated Modulation of Metastasis-Associated Protein 1 Methylation and Inflammation in Brain Arteriovenous Malformation. Biomolecules 2023, 13, 1495. https://doi.org/10.3390/biom13101495
Deng X, Zhou S, Hu Z, Gong F, Zhang J, Zhou C, Lan W, Gao X, Huang Y. Nicotinic Acid-Mediated Modulation of Metastasis-Associated Protein 1 Methylation and Inflammation in Brain Arteriovenous Malformation. Biomolecules. 2023; 13(10):1495. https://doi.org/10.3390/biom13101495
Chicago/Turabian StyleDeng, Xinpeng, Shengjun Zhou, Ziliang Hu, Fanyong Gong, Junjun Zhang, Chenhui Zhou, Wenting Lan, Xiang Gao, and Yi Huang. 2023. "Nicotinic Acid-Mediated Modulation of Metastasis-Associated Protein 1 Methylation and Inflammation in Brain Arteriovenous Malformation" Biomolecules 13, no. 10: 1495. https://doi.org/10.3390/biom13101495