
Arbuscular Mycorrhizal Fungi Improve Lycium barbarum Potassium Uptake by Activating the Expression of LbHAK


1
College of Forestry, Northwest A&F University, Yangling 712100, China
2
Shaanxi Engineering Research Center of Forage Plants of the Loess Plateau, College of Life Sciences, Yulin University, Yulin 719000, China
3
State Key Laboratory of Conservation and Utilization of Subtropical Agro-Bioresources, College of Forestry and Landscape Architecture, South China Agricultural University, Guangzhou 510642, China
*
Authors to whom correspondence should be addressed.
Plants 2024, 13(9), 1244; https://doi.org/10.3390/plants13091244
Submission received: 4 April 2024
/
Revised: 26 April 2024
/
Accepted: 29 April 2024
/
Published: 30 April 2024
(This article belongs to the Section Plant–Soil Interactions)
Abstract
:Arbuscular mycorrhizal (AM) fungi can establish a mutualistic relationship with the roots of most terrestrial plants to increase plant nutrient uptake. The effects of potassium uptake and transport by AM symbiosis are much less reported compared to other nutrients. In this research, a heterologous yeast system was used to verify that the LbHAK has capacity for potassium uptake. The split-roots system implemented using seedlings of Lycium barbarum confirmed that R. irregularis locally induced LbHAK expression, which means that LbHAK is only expressed in mycorrhizal roots. Furthermore, the impacts of overexpression of LbHAK on the growth, nutrients and water uptake, and transport of mycorrhizal tobacco (inoculation with Rhizophagus irregularis) at 0.2 mM and 2 mM K conditions were assessed. The mycorrhizal tobacco growth and potassium accumulation were significantly enhanced through LbHAK overexpression in tobacco. In addition, overexpression of LbHAK substantially enhanced phosphorus content, while stimulating the expression of NtPT4, Rir-AQP1, and Rir-AQP2 in mycorrhizal tobacco. Moreover, LbHAK overexpression greatly promoted AM colonization. LbHAK has a potential role in facilitating potassium absorption through the mycorrhizal pathway, and overexpression of LbHAK in tobacco may promote the transport of potassium, phosphorus, and water from AM fungi to tobacco. These data imply the important roles played by the LbHAK in AM-fungi-induced potassium uptake in L. barbarum and in improving plant nutrients and AM colonization.
1. Introduction
More than 80% of terrestrial plants can be colonized by arbuscular mycorrhizal (AM) fungi, which belong to monophyletic phylum Glomeromycota [1]. The symbiotic system between AM fungi and plants improves plant growth, particularly in nutrient-limited conditions [2,3]. Mycorrhizal plants can not only directly uptake water and nutrients through plant roots, but also indirectly through AM fungal hyphae, which expands the absorption area of plants and improves the nutrition and water status of plants and is also known as the mycorrhizal pathway [4]. Many studies have provided evidence indicating that the mycorrhizal pathway is crucial for the absorption of phosphate (Pi), and several Pi transporters in plants have been identified, which are induced by AM fungi, such as MtPT4, NtPT4, and LbPT4 [5,6,7]. The increasing evidence shows that the mycorrhizal pathway can also uptake substantial amounts of nitrogen, copper, and sulfur [8,9,10,11,12].
Potassium is a vital nutritional element in plants involved in many metabolic processes [13,14]. Potassium plays important roles in plant growth and development, as well as in coping with adverse conditions, especially drought stress [13,15,16,17]. Higher cytosolic potassium/sodium ratios contribute to enhanced salt tolerance in plants [18]. Potassium could promote chlorophyll synthesis and increases the rate of photosynthesis, which enhances plant resistance to drought [19,20]. Furthermore, potassium application can increase the activities of superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) in plants, thereby promoting the capacities of scavenging reactive oxygen species and reducing the damage caused by the accumulation of oxidative substances by drought stress [17,21,22]. Additionally, potassium also contributes to plant cell osmosis, promoting the accumulation of substances such as proline, helping plants overcome drought environments [23,24]. Application of potassium fertilization in Zea mays L. significantly reduces water consumption and increases water use efficiency [25]. In addition to drought stress, potassium fertilization also promotes the uptake and transport of nitrogen and phosphorus, thereby improving the nutritional conditions of plants [26]. However, potassium availability fluctuates widely in soils around the world and is often seen as a constraint on plant growth [27,28]. Thus, improving the capacity of potassium uptake is critical for plant growth and adaptation to stress conditions [13].
Garcia and Zimmermann (2014) [29] reported a strong potassium accumulation in AM fungi and in some plant tissues, implying the possibility of mycorrhizal potassium uptake. Moreover, previous studies discovered the AM-fungi-induced potassium transporters in Lotus japonicus (LjHAK) and tomato (SlHAK10) and proved that SlHAK10 mediated the mycorrhizal potassium uptake pathway [30,31]. Lycium barbarum L. is an important traditional medical plant in northwestern China, where water is one of main limiting factors for plant growth [6,32]. In the arid and semi-arid regions, the mobility of readily available potassium is reduced, making it more difficult to utilize, so plants often face a lack of potassium nutrition under drought conditions [24]. In this study, the expression of a putative orthologue gene of SlHAK10 (designated as LbHAK) in mycorrhizal L. barbarum roots was significantly induced compared with non-mycorrhizal roots [33]. We evaluated the capability of LbHAK in potassium uptake by conducting complementation analysis in a yeast (Saccharomyces cerevisiae) mutant deficient in potassium uptake (CY162). A split-root experiment with L. barbarum seedlings was carried out to investigate whether the expression of LbHAK was systemically or locally induced by AM fungi. Overexpression of LbHAK driven by the CaMV35S promoter in tobacco was conducted to further evaluate the function of LbHAK by comparing the growth, AM colonization, nutrient status, and gene expression with wild-type tobacco under different potassium conditions.
2. Materials and Methods
2.1. Growth Substrate and AM Fungal Inoculum
The growth substrate was a mixture of sand and vermiculite (1:1, v:v). The sand was sieved through a 2 mm sieve and thoroughly washed with tap water. Sand and vermiculite were autoclaved at 121 °C for 2 h twice with one-day interval.
The inoculum of Rhizophagus irregularis (BGC BJ109) was propagated with Plantago asiatica for 12 months. The inoculum consisted of the sandy substrate that contained spores, mycelium, and colonized root fragments. The inoculum consisted of the sandy substrate that contained spores, mycelium, and colonized root fragments containing 150 propagules per gram, determined by the most probable number method.
2.2. Clone and Bioformation Analysis of LbHAK
The partial sequence of LbHAK was obtained by independent de novo transcriptome sequencing (unpublished). The full LbHAK sequence was amplified by the 5′ and 3′ rapid amplification of cDNA ends (RACE) procedure using the SMARTer™ RACE cDNA Amplification Kit (Clontech Laboratories, Inc., Mountain View, CA, USA). The primers used are listed in Table S1.
The Open Reading Frame (ORF) Finder (https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 3 April 2024)) in NCBI was utilized for ORF analysis and amino acid sequence prediction. The ProtParam tool (http://web.expasy.org/protparam/ (accessed on 3 April 2024)) was utilized for the prediction of the molecular weight and isoelectric point of proteins. The subcellular localization was predicted by TargetP 2.0 Server (http://www.cbs.dtu.dk/services/TargetP/ (accessed on 3 April 2024)). DeepTMHMM (https://dtu.biolib.com/DeepTMHMM (accessed on 3 April 2024)) was used to predict transmembrane helices in proteins. The neighbor-joining (NJ) method was employed by MEGA 6.0.6 to construct a phylogenetic tree.
2.3. Heterologous Expression of LbHAK in Yeast
The LbHAK coding sequence was inserted into the KpnI to SacI sites of pYES2 yeast expression vector. The recombinant vector and pYES2 vector (Table S2) were separately introduced into the yeast potassium-uptake-defective mutant strain CY162 (MATα ura3-52 his4-15 trk1Δ trk2Δ1::pCK64) (Table S1) [34]. The wild-type yeast strain was BY4741 (Table S2) [35]. The LiAC/single-stranded DNA/polyethylene glycol method was utilized for yeast cell transformation, and the growth was measured on AP medium (supplemented with galactose) as suggested by Horie et al. [36].
2.4. Split-Root Experiment Analysis
A split-root experiment was conducted to test whether LbHAK was only expressed in AM-fungi-colonized root parts of plants (Figure S1). The preparation of L. barbarum seedlings, the split-root system, the setup of mycorrhizal (AM) and non-mycorrhizal (NM) treatments, and the cultivation and harvest were in accordance with the description of Han et al. [12].
2.5. Overexpression of LbHAK in Tobacco
The complete coding sequence of LbHAK was cloned and integrated to the pROKII vector (Table S2) at the SmaI to SacI sites. The LbHAK-pROKII recombinant vector was transformed into the Agrobacterium tumefaciens GV3101. The leaf disc method was employed to accomplish A. tumefaciens-mediated transformation of tobacco (Nicotiana tabacum), followed by cultivation of the transformed plants on Murashige and Skoog medium.
Wild-type (WT) and LbHAK-overexpression (OE) tobacco lines were used for the pot experiment. T0 seeds of WT and OE tobacco were surface-disinfected with 5% NaClO for 5 min and washed 3 times with sterile water. The sterile seeds were then germinated on moist filter paper at 28 °C in darkness. The germinated seeds were planted in a tray filled with growth substrate same as mentioned above for 5 weeks. Uniform tobacco seedlings of two lines were selected for transplantation. The seedlings were grown in a pot (10 cm × 10 cm × 9.6 cm) containing 400 g of growth substrate. The seedlings in AM treatment received 10 g AM fungal inoculum beneath roots during transplantation, while seedlings in NM treatment received autoclaved inoculum (121 °C for 2 h) and 10 mL filtrate (<20 μm nylon mesh) of inoculum. After transplantation, seedlings of tobacco were watered daily and irrigated weekly with 20 mL Hoagland solution containing 10% phosphate (0.1 mM KH2PO4) to ensure the colonization of AM fungi. After 3 weeks of growth, two concentrations (0.2 and 2 mmol K kg−1 growth substrate) of K2SO4 solution (20 mL) were applied to the tobacco seedlings, and the growth continued for another 3 weeks. There were 8 treatments in total. Each treatment had four replicates, and 2 seedlings were included in each replicate. The pot experiment was carried out in a controlled greenhouse environment where there was 16 h light per day, along with a temperature range of 24–28 °C and a relative humidity range of 40–60%.
2.6. Measurement of Plant Biomass, Mycorrhizal Colonization, and Potassium and Phosphorus Concentrations
At harvest, the fresh weight of the shoot and root of tobacco in the pot experiment were recorded, respectively. Parts of the shoots and roots of tobacco were killed green at 110 °C for 15 min, and then dried at 65 °C until the samples reached a constant weight. The remaining portions of shoots and roots from the split-root experiment and pot experiment were immediately frozen and ground into a powder using liquid nitrogen, and then stored at −80 °C until further use in relative gene expression analyses.
Part of the roots from the split-root experiment and pot experiment were cut into 1 cm fragments and stained with trypan blue [37], and then mycorrhizal colonization rate was assessed using the magnified intersection method with a light microscope [38].
Plant dry materials were ground and digested in accordance with Han et al. [39]. The flame photometer was used to measure potassium concentration and using method of molybdenum yellow colorimetric to determine phosphorus concentration [40]. The content was calculated by the concentration and dry weight of potassium and phosphorus.
2.7. Relative Gene Expression Analysis
Total RNA was extracted by the E.Z.N.ATM Plant RNA kit (Omega Bio-Tek, Norcross, GA, USA). Following the instructions provided by the supplier, 1 μg of high-quality RNA was digested DNAse utilizing the Hifair® Ⅲ 1st Strand cDNA Synthesis SuperMix for qPCR (gDNA digester plus) (YEASEN Bio, Shanghai, China). After DNA digestion, the control PCR was applied to check the possible trace DNA contamination. Afterwards, the RNA was transcribed to cDNA using the above-mentioned kit, and the cDNA was then employed as a template for PCR reactions. The transcript accumulation analysis of LbHAK, GintEF1α, NtPT4, Rir-AQP1, and Rir-AQP2 in roots and R. irregularis hyphae was performed by SYBR green-based qRT-PCR. The Lbactin and NtEF1a were used as an internal control of L. barbarum and tobacco roots, respectively. The GintEF1α was used as an internal control of R. irregular hyphae. The specific primers for qRT-PCR can be found in Table S1. The reactions of qRT-PCR were performed by the description of Han et al. [12]. The transcript abundance of LbHAK in the overexpression lines of tobacco was also evaluated by qRT-PCR as mentioned above; the NtEF1a gene was used as internal reference gene to normalize the expression data. The relative expressions were determined using the equation 2−ΔΔCT [41].
2.8. Statistical Analysis
IBM SPSS Statistics 21.0 software (IBM, Armonk, NY, USA) was conducted for statistical analysis. Data were analyzed by a three-way ANOVA (tobacco lines, AM status, and potassium status) and Duncan’s test. Spearman’s correlation was used for correlation analyses with data of root nutrient contents, colonization rate, and gene expression in tobacco roots and R. irregularis. Origin Pro 2023 (Origin Lab, Northampton, MA, USA) was utilized to create the figures.
3. Results
3.1. Identification and Functional Analysis of LbHAK
The complete cDNA sequence of LbHAK, which encodes a potassium transporter from the HAK/KUP/KT family in L. barbarum, was deposited in GenBank (accession NO. MZ416922.1). The LbHAK gene featured a 2004 bp ORF, potentially encoding 668 amino acids, and was predicted to have a molecular weight of 97.35 kDa (Table 1). The predicted isoelectric point of LbHAK was estimated as 9.07, furthermore LbHAK protein was predicted to be in the plasma membrane (Table 1). The deduced protein of LbHAK had ten transmembrane helices (Figure S2A). The outcome of multiple sequence alignment and phylogenetic tree analysis suggested that LbHAK is an orthologue of SlHAK10 and belongs to the cluster II of the HAK/KUP/KT family (Figure 1 and Figure S2B).
All the tested yeasts grew well on the AP medium at 10 mM and 5 mM KCl concentrations (Figure 2A). The potassium-uptake-defective yeast mutant CY162 with pYES2 showed growth inhibition at 1 mM KCl concentration and stopped growth at 0.5 mM KCl concentration (Figure 2A). Yeast mutant CY162 with LbHAK-pYES2 resembled the growth of wild-type yeast (BY4741) with pYES2 at 1 mM and 0.5 mM KCl concentrations (Figure 2A). Growth curve analysis showed that the growth rate of CY162 with LbHAK-pYES2 and wild-type yeast with pYES2 was significantly higher than that of the mutant yeast CY162 with pYES2 at 0.5 mM KCl concentration (Figure 2B). These data suggest the LbHAK can function as potassium transporter.
To characterize the expression patterns of LbHAK upon AM treatments in roots of L. barbarum, the split-root experiment was conducted (Figure S1). The LbHAK was only expressed in the AM-fungal-colonized root part of the AM treatment in the split-root experiment (Figure 3). No expression of LbHAK was detected in the non-colonized root part of the AM and NM treatments (Figure 3). These data suggest that the expression of LbHAK was specifically induced by AM in L. barbarum roots.
3.2. Overexpression of LbHAK Increased Growth and AM Colonization of Tobacco
A total of 8 different transgenic tobacco lines overexpressing LbHAK were obtained (Figure S3). Inoculation of R. irregularis significantly improved the fresh weight of shoots, roots, and plants in WT and OE tobacco at 0.2 mM and 2 mM potassium conditions in the pot experiment (Table S3 and Figure 4A–C). The roots and total fresh weight of all tobacco were significantly increased under 2 mM potassium (Table S3 and Figure 4B,C). The overexpression of LbHAK led to a notable increase in the shoot and root fresh weight of non-mycorrhizal tobacco under two potassium conditions and in mycorrhizal tobacco under 0.2 mM potassium condition (Figure 4A,B). Additionally, it enhanced the total fresh weight of all tobacco plants under two potassium conditions (Figure 4C).
In addition to growth, OE tobacco plants showed an obviously increased mycorrhizal colonization rate and arbuscular rate at two potassium conditions in the pot experiment (Figure 5A,B). Application of 2 mM potassium also obviously increased the arbuscular rate in WT and OE tobacco (Figure 5B). The expression of GintEF1α and NtPT4 were only expressed in mycorrhizal tobacco (Figure 5C,D). Overexpression of LbHAK significantly up-regulated GintEF1α expression of mycorrhizal tobacco at 2 mM potassium condition and mycorrhizal tobacco NtPT4 expression at two potassium conditions (Figure 5C,D). Application of 2 mM potassium significantly enhanced the expression of GintEF1α and NtPT4 in mycorrhizal OE tobacco plants (Figure 5C,D).
3.3. Overexpression of LbHAK Increased Tobacco Potassium and Phosphorus Uptake
Inoculation of R. irregularis significantly increased all tobacco shoot, root, and total potassium contents at two potassium conditions (Figure 6A–C). Application of 2 mM potassium significantly increased non-mycorrhizal WT tobacco shoot and total potassium contents and non-mycorrhizal OE tobacco root potassium content (Figure 6A–C). LbHAK overexpression significantly enhanced the non-mycorrhizal tobacco shoot and total potassium contents at 0.2 mM potassium condition and root potassium content at 2 mM potassium condition (Figure 6A–C).
Additionally, inoculation of R. irregularis significantly increased the phosphorus contents in all tobacco shoots, roots, and whole plants irrespective of potassium conditions (Table S3 and Figure 7A–C). The mycorrhizal OE tobacco shoot, root, and total phosphorus contents were significantly increased under 2 mM potassium, as well as non-mycorrhizal WT tobacco root phosphorus content and mycorrhizal WT tobacco total phosphorus content (Table S3 and Figure 7A–C). LbHAK overexpression significantly increased the non-mycorrhizal tobacco shoot, root, and total phosphorus contents at 0.2 mM potassium condition; mycorrhizal tobacco shoot phosphorus content at 2 mM potassium condition; and root and total phosphorus contents at two potassium conditions (Table S3 and Figure 7A–C).
3.4. Overexpression of LbHAK Increased Rir-AQP1 and Rir-AQP2 Expression
The expression levels of two marker genes (Rir-AQP1 and Rir-AQP2) were only detected in tobacco plants with mycorrhizal roots (Figure 8A,B). Overexpression of LbHAK significantly stimulated the expression levels of Rir-AQP1 irrespective of potassium conditions and the expression of Rir-AQP2 at 2 mM potassium condition (Table S3 and Figure 8A,B). Application of 2 mM potassium significantly increased the expression of Rir-AQP1 in OE tobacco and the expression of Rir-AQP2 in WT and OE tobacco (Table S3 and Figure 8A,B). These data suggest that overexpression of LbHAK may stimulate the expressions of Rir-AQP1 and Rir-AQP2, thus improving the water transport capacities of host plants.
3.5. Correlation Analysis
To unravel the possible relationships between the detected parameters, correlation analysis was conducted (Figure 9). The mRNA abundance of the NtPT4 gene was significantly positively correlated with mRNA levels of Rir-AQP1 and Rir-AQP2, arbuscular rate, as well as the potassium and phosphorus contents in roots (Figure 9). Root potassium content was also closely positively correlated with the mRNA levels of two genes (Rir-AQP1 and Rir-AQP2), arbuscular rate, and root phosphorus content (Figure 9).
4. Discussion
The plants colonized by AM fungi can not only directly uptake water and nutrients through plant roots, but also indirectly through AM fungal hyphae, which is known as the mycorrhizal pathway [4]. The mycorrhizal pathway is crucial for the absorption of Pi, and several AM-induced Pi transporters in plants have been identified [5,6]. Comparatively, the effects of AM symbiosis on the accumulation of potassium in plants received far less attention. Although there were reports that AM fungi improve plant potassium content, especially under potassium-limited condition [29,32], the direct evidence of the mycorrhizal pathway for potassium uptake was not discovered until the AM fungi specifically induced plant potassium transporter gene (SlHAK10 in tomato) was functionally analyzed [30].
L. barbarum, which belongs to the Solanaceae family like tomato, tobacco, etc., could be colonized by AM fungi and nutritionally promoted by AM symbiosis [6,12,32,42]. A putative orthologue gene of SlHAK10 in L. barbarum (designated as LbHAK) was isolated based on an independent de novo transcriptomic analysis (unpublished). The multiple sequence alignment and phylogenetic tree analysis indicated that LbHAK belongs to AM-induced cluster II of the HAK/KUP/KT family and has high similarity with SlHAK10 (Figure 1 and Figure S2B). The heterologous yeast system certified the potassium transport capacity of LbHAK (Figure 2), and the expression of LbHAK was restricted within the AM-fungi-colonized root part (Figure 3). These results implied that LbHAK functions as a potassium transporter and may participate in the mycorrhizal pathway in L. barbarum as SlHAK10 in tomato.
To further characterize the function of LbHAK, LbHAK was transformed into tobacco with the constitutive CaMV35S promoter. Despite the addition of potassium, LbHAK OE tobacco plants exhibited obvious growth promotion compared with WT tobacco plants (Figure 4). The result resembled our previous results when the potassium channel gene LbKAT3 [12] and ammonium transport gene LbAMT3-1 [10] were overexpressed in tobacco. The CaMV35S promoter-driven nutrition uptake genes from L. barbarum perform functions in tobacco [43]. However, the overexpression of SlHAK10 only promoted tomato growth with the inoculation of AM fungi under potassium-limited condition [30]. The potassium concentration in growth substrates and potassium demand of different plants may be the reason for this difference [12,30]. When the AM fungi were taken into consideration, the growth of LbHAK OE tobacco plants was further promoted. On one hand, this was a commonly observed result of the AM symbiosis effect on plant growth [33,39]. On the other hand, this may be a hint that LbHAK is in the preferred location in the peri-arbuscular membrane as its orthologue SlHAK10 and performs potassium transport function for AM symbiosis.
In AM symbiosis, most nutrient transport through the mycorrhizal pathway takes place in the plant root cells harboring arbuscules [3,44]. Limitation of nutrient transfer from AM fungi to host plants usually exhibits limited or even eliminated arbuscular function and abundance [6,45]. In this study, LbHAK OE tobacco plants exhibited a highly increased colonization rate and arbuscular rate, and the increment was more obvious when a high amount of potassium was available (Figure 5). The expression of NtPT4, which is the indicator for the viability of arbuscules and phosphate transport in tobacco [7], was simultaneously increased in LbHAK OE tobacco plants. Although the variation of potassium availability in substrate did not show obvious influence on root colonization in Medicago truncatula, the expression of AM-specific marker gene (MtPT4) was up-regulated at high potassium condition [46]. The up-regulation of AM-specific phosphate transporter gene (LbPT4) was also observed when the folia-applied potassium was transferred via mycelia network to L. barbarum that were grown under potassium-limited condition [33]. These alike results suggested that the AM symbiosis has a promoted performance for phosphate transport under high potassium available condition [12].
More than promoted colonization, LbHAK OE tobacco also exhibited promoted potassium and phosphorus uptake, and the promotion was especially obvious when tobacco was colonized by AM fungi (Figure 6 and Figure 7). This was another piece of evidence for the up-regulated expression of NtPT4 and the above-discussed point that AM symbiosis promotes plant phosphorus transport when more potassium is available. The simultaneously promoted potassium and phosphorus on one hand may be due to the co-distribution and a linked ratio by the nature of AM fungi (R. irregularis) [47,48]. On the other hand, the role of potassium as one of the major counter-ions of polyphosphate [49], which is the form of phosphate transfer along the hyphae of AM fungi to host plants, may explain the simultaneous potassium and phosphorus promotion.
Based on the model proposed by Kikuchi et al. (2016) [50], the transfer of polyphosphate along AM fungal hyphae required water flow that was facilitated by aquaporin in AM fungi. The expression of AM fungal aquaporin genes (Rir-AQP1 and Rir-AQP2) was up-regulated in the LbHAK OE tobacco, especially under high potassium condition (Figure 8). Coincidentally, the expression of NtPT4, Rir-AQP1, Rir-AQP2, and the potassium and phosphorus contents in roots showed positive correlations (Figure 9). These results not only supported the model that the transport of phosphate via AM symbiosis requires the participation of AM fungal aquaporin, but also suggested a co-transport of phosphate and potassium in AM fungi [33,47,48]. Moreover, the arbuscular abundance, the expression of NtPT4, and the content of phosphorus and potassium in the mycorrhizal LbHAK OE tobacco plants were always higher than those in the mycorrhizal WT tobacco plants. Considering the mycorrhizal tobacco plants were under similar phosphate nutrient condition, the promoted function of AM symbiosis in the LbHAK OE tobacco suggested that there may be a regulatory mechanism between potassium and phosphorus in tobacco for the regulation of AM fungi [29].
5. Conclusions
In conclusion, LbHAK belongs to cluster II of the HAK/KUP/KT family. The expression of LbHAK is restricted within plant root parts that are colonized by AM fungi, and LbHAK may preferentially locate in the peri-arbuscular membrane. Compared with wild-type tobacco, LbHAK-overexpressing tobacco exhibited promoted growth, colonization rate, arbuscular abundance, and potassium and phosphorus content, especially when a high amount of potassium was available to the mycorrhizal plants. LbHAK may be involved in the mycorrhizal pathway for the potassium transport that accompanies phosphate transport. Future studies focused on the regulatory mechanism of plants on AM symbiosis between potassium and phosphorus are needed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants13091244/s1, Figure S1: Schematic diagram of the split-roots system; Figure S2: Bioinformation analysis of LbHAK; Figure S3: Detection of LbHAK expression in transgenic tobacco; Table S1: Primers used for clone and qRT-PCR; Table S2: Information for yeast strains and plasmids [34,35,51,52]; Table S3: ANOVA analysis of the detected parameters in this study.
Author Contributions
Y.Z. designed the research and performed the experiments, analyzed the data, and wrote the manuscript. X.H. and W.R. provided the experimental methods. H.Z. revised the manuscript. M.T. and H.Z. participated in the design of this research. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (42277027, 31700530, and 32071639), the Laboratory of Lingnan Modern Agriculture Project (NZ2021025), and the State Key Laboratory for Conservation and Utilization of Subtropical Agro-bioresources (SKLCUSA-b202007).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors thank Fengxin Zhang for his contributions to the preparatory works of the research. Rhizophagus irregularis (BGC BJ109) was kindly provided by the Beijing Academy of Agriculture and Forestry. The yeast strain CY162 was kindly provided by the National BioResource Project (NBRP) of the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan. The wild-type yeast strain BY4741 was donated by Chen Peng (College of Life Sciences, Northwest A&F University). The seeds of L. barbarum were kindly provided by Yajun Wang (Ningxia Academy of Agriculture and Forestry Sciences).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Smith, S.E.; Smith, F.A. Roles of Arbuscular Mycorrhizas in Plant Nutrition and Growth: New Paradigms from Cellular to Ecosystem Scales. Annu. Rev. Plant Biol. 2011, 62, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Qin, C.; Ahanger, M.A.; Raza, S.; Khan, M.I.; Ashraf, M.; Ahmed, N.; Zhang, L. Role of Arbuscular Mycorrhizal Fungi in Plant Growth Regulation: Implications in Abiotic Stress Tolerance. Front. Plant Sci. 2019, 10, 1068. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Kafle, A. Role of Arbuscular Mycorrhizal Fungi in the Nitrogen Uptake of Plants: Current Knowledge and Research Gaps. Agronomy 2015, 5, 587–612. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R.; Glick, B.R. Contribution of Arbuscular Mycorrhizal Fungi, Phosphate–Solubilizing Bacteria, and Silicon to P Uptake by Plant. Front. Plant Sci. 2021, 12, 699618. [Google Scholar] [CrossRef] [PubMed]
- Javot, H.; Penmetsa, R.V.; Terzaghi, N.; Cook, D.R.; Harrison, M.J. A Medicago truncatula phosphate transporter indispensable for the arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2007, 104, 1720–1725. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Zhang, H.; Zhang, X.; Chen, H.; Tang, M. Characterization of six PHT1 members in Lycium barbarum and their response to arbuscular mycorrhiza and water stress. Tree Physiol. 2017, 37, 351–366. [Google Scholar] [PubMed]
- Chen, A.; Hu, J.; Sun, S.; Xu, G. Conservation and divergence of both phosphate- and mycorrhiza-regulated physiological responses and expression patterns of phosphate transporters in Solanaceous species. New Phytol. 2006, 173, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Giovannetti, M.; Tolosano, M.; Volpe, V.; Kopriva, S.; Bonfante, P. Identification and functional characterization of a sulfate transporter induced by both sulfur starvation and mycorrhiza formation in Lotus japonicus. New Phytol. 2014, 204, 609–619. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Chen, A.; Xie, K.; Yang, X.; Luo, Z.; Chen, J.; Zeng, D.; Ren, Y.; Yang, C.; Wang, L.; et al. Functional analysis of the OsNPF4.5 nitrate transporter reveals a conserved mycorrhizal pathway of nitrogen acquisition in plants. Proc. Natl. Acad. Sci. USA 2020, 117, 16649–16659. [Google Scholar] [CrossRef]
- Cheng, K.; Wei, M.; Jin, X.; Tang, M.; Zhang, H. LbAMT3-1, an ammonium transporter induced by arbuscular mycorrhizal in Lycium barbarum, confers tobacco with higher mycorrhizal levels and nutrient uptake. Plant Cell Rep. 2022, 41, 1477–1480. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Lin, A.-J.; Gao, Y.-L.; Reid, R.J.; Wong, M.-H.; Zhu, Y.-G. Arbuscular mycorrhizal colonisation increases copper binding capacity of root cell walls of Oryza sativa L. and reduces copper uptake. Soil Biol. Biochem. 2009, 41, 930–935. [Google Scholar] [CrossRef]
- Han, X.; Zhou, Y.; Li, Y.; Ren, W.; Liu, K.; Zhang, W.; Zhang, H.; Tang, M. LbKAT3 may assist in mycorrhizal potassium uptake, and overexpression of LbKAT3 may promote potassium, phosphorus, and water transport from arbuscular mycorrhizal fungi to the host plant. Front. Plant Sci. 2023, 14, 1161220. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, W.-H. Potassium Transport and Signaling in Higher Plants. Annu. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Shankar, A.; Chandran, A.K.N.; Sharma, M.; Jung, K.-H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2019, 71, 608–619. [Google Scholar] [CrossRef] [PubMed]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in Root Growth and Development. Plants 2019, 8, 435. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, J.; Asensio, V.; Bordron, B.; Nouvellon, Y.; le Maire, G.; Bouillet, J.; Domec, J.; Rojas, J.S.D.; Abreu-Junior, C.H.; Battie-Laclau, P.; et al. Increased hydraulic constraints in Eucalyptus plantations fertilized with potassium. Plant Cell Environ. 2021, 44, 2938–2950. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liao, W. Potassium signaling in plant abiotic responses: Crosstalk with calcium and reactive oxygen species/reactive nitrogen species. Plant Physiol. Biochem. 2022, 173, 110–121. [Google Scholar] [CrossRef]
- Mulet, J.M.; Porcel, R.; Yenush, L. Modulation of potassium transport to increase abiotic stress tolerance in plants. J. Exp. Bot. 2023, 74, 5989–6005. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Lyu, C.; Huang, L.; Chen, Q.; Zhuo, W.; Wang, X.; Lu, Y.; Zeng, F.; Lu, L. Physiology and proteomic analysis reveals root, stem and leaf responses to potassium deficiency stress in alligator weed. Sci. Rep. 2019, 9, 17366. [Google Scholar] [CrossRef]
- Battie-Laclau, P.; Laclau, J.-P.; Beri, C.; Mietton, L.; Muniz, M.R.A.; Arenque, B.C.; DE Cassia Piccolo, M.; Jordan-Meille, L.; Bouillet, J.-P.; Nouvellon, Y. Photosynthetic and anatomical responses of Eucalyptus grandis leaves to potassium and sodium supply in a field experiment. Plant Cell Environ. 2014, 37, 70–81. [Google Scholar] [CrossRef]
- Fang, S.; Yang, H.; Wei, G.; Shen, T.; Wan, Z.; Wang, M.; Wang, X.; Wu, Z. Potassium application enhances drought tolerance in sesame by mitigating oxidative damage and regulating osmotic adjustment. Front. Plant Sci. 2022, 13, 1096606. [Google Scholar] [CrossRef]
- Zhu, B.; Xu, Q.; Zou, Y.; Ma, S.; Zhang, X.; Xie, X.; Wang, L. Effect of potassium deficiency on growth, antioxidants, ionome and metabolism in rapeseed under drought stress. Plant Growth Regul. 2019, 90, 455–466. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant. 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Du, X.-Q.; Wang, F.-L.; Li, H.; Jing, S.; Yu, M.; Li, J.; Wu, W.-H.; Kudla, J.; Wang, Y. The transcription factor MYB59 regulates K+/NO3− translocation in the Arabidopsis response to low K+ stress. Plant Cell 2019, 31, 699–714. [Google Scholar] [CrossRef]
- Rengel, Z.; Damon, P.M. Crops and genotypes differ in efficiency of potassium uptake and use. Physiol. Plant. 2008, 133, 624–636. [Google Scholar] [CrossRef]
- Beier, S.; Marella, N.C.; Yvin, J.-C.; Hosseini, S.A.; von Wirén, N. Silicon mitigates potassium deficiency by enhanced remobilization and modulated potassium transporter regulation. Environ. Exp. Bot. 2022, 198, 104849. [Google Scholar] [CrossRef]
- Garcia, K.; Zimmermann, S.D. The role of mycorrhizal associations in plant potassium nutrition. Front. Plant Sci. 2014, 5, 337. [Google Scholar] [CrossRef]
- Liu, J.; Liu, J.; Liu, J.; Cui, M.; Huang, Y.; Tian, Y.; Chen, A.; Xu, G. The Potassium Transporter SlHAK10 Is Involved in Mycorrhizal Potassium Uptake. Plant Physiol. 2019, 180, 465–479. [Google Scholar] [CrossRef]
- Guether, M.; Balestrini, R.; Hannah, M.; He, J.; Udvardi, M.K.; Bonfante, P. Genome-wide reprogramming of regulatory networks, transport, cell wall and membrane biogenesis during arbuscular mycorrhizal symbiosis in Lotus japonicus. New Phytol. 2009, 182, 200–212. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, S.; Hu, W.; Xiao, L.; Tang, M. Arbuscular Mycorrhizal Fungus Rhizophagus irregularis Increased Potassium Content and Expression of Genes Encoding Potassium Channels in Lycium barbarum. Front. Plant Sci. 2017, 8, 440. [Google Scholar] [CrossRef]
- Han, X.; Du, X.; Wu, Y.; Wei, M.; Gu, Y.; Aba, X.; Tang, M.; Zhang, H. Foliar-applied potassium improved mycorrhizal Goji (Lycium barbarum L.) growth of the potassium free—Compartment in a compartmented culture system. Sci. Hortic. 2021, 293, 110681. [Google Scholar] [CrossRef]
- Anderson, J.A.; Huprikar, S.S.; Kochian, L.V.; Lucas, W.J.; Gaber, R.F. Functional expression of a probable Arabidopsis thaliana potassium channel in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1992, 89, 3736–3740. [Google Scholar] [CrossRef]
- Winston, F.; Dollard, C.; Ricupero-Hovasse, S.L. Construction of a set of convenient Saccharomyces cerevisiae strains that are isogenic to S288C. Yeast 1995, 11, 53–55. [Google Scholar] [CrossRef]
- Horie, T.; Brodsky, D.E.; Costa, A.; Kaneko, T.; Lo Schiavo, F.; Katsuhara, M.; Schroeder, J.I. K+ Transport by the OsHKT2; 4 Transporter from Rice with Atypical Na+ Transport Properties and Competition in Permeation of K+ over Mg2+ and Ca2+ Ions. Plant Physiol. 2011, 156, 1493–1507. [Google Scholar] [CrossRef]
- Koske, R.; Gemma, J. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 94, 486–488. [Google Scholar] [CrossRef]
- McGONIGLE, T.P.; Miller, M.H.; Evans, D.G.; Fairchild, G.L.; Swan, J.A. A new method which gives an objective measure of colonization of roots by vesicular—Arbuscular mycorrhizal fungi. New Phytol. 1990, 115, 495–501. [Google Scholar] [CrossRef]
- Han, X.; Wang, Y.; Cheng, K.; Zhang, H.; Tang, M. Arbuscular Mycorrhizal Fungus and Exogenous Potassium Application Improved Lycium barbarum Salt Tolerance. J. Plant Growth Regul. 2021, 41, 2980–2991. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Methods of Soil Analysis Part 2 Chemical and Microbiological Properties, American Society of Agronomy; Page, A.L., Ed.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gong, M.; Zhang, Q.; Cheng, K.; Zhang, H. Symbiosis of arbuscular mycorrhizal fungi and Lycium barbarum L. Prefers NO3− over NH4+. Horticulturae 2023, 9, 637. [Google Scholar] [CrossRef]
- Wu, J.; Ji, J.; Wang, G.; Wu, G.; Diao, J.; Li, Z.; Chen, X.; Chen, Y.; Luo, L. Ectopic expression of the Lycium barbarum β-carotene hydroxylase gene (chyb) enhances drought and salt stress resistance by increasing xanthophyll cycle pool in tobacco. Plant Cell Tissue Organ Cult. (PCTOC) 2015, 121, 559–569. [Google Scholar] [CrossRef]
- Xie, K.; Ren, Y.; Chen, A.; Yang, C.; Zheng, Q.; Chen, J.; Wang, D.; Li, Y.; Hu, S.; Xu, G. Plant nitrogen nutrition: The roles of arbuscular mycorrhizal fungi. J. Plant Physiol. 2021, 269, 153591. [Google Scholar] [CrossRef]
- Jiménez-Leyva, J.A.; Gutiérrez, A.; Orozco, J.A.; Vargas, G.; Esqueda, M.; Gardea, A.; González-Hernández, V.; Sánchez, E.; Muñoz, E. Phenological and ecophysiological responses of Capsicum annuum var. glabriusculum to native arbuscular mycorrhizal fungi and phosphorus availability. Environ. Exp. Bot. 2017, 138, 193–202. [Google Scholar] [CrossRef]
- Garcia, K.; Chasman, D.; Roy, S.; Ané, J.-M. Physiological Responses and Gene Co-Expression Network of Mycorrhizal Roots under K+ Deprivation. Plant Physiol. 2017, 173, 1811–1823. [Google Scholar] [CrossRef]
- Olsson, P.A.; Hammer, E.C.; Pallon, J.; van Aarle, I.M.; Wallander, H. Elemental composition in vesicles of an arbuscular mycorrhizal fungus, as revealed by PIXE analysis. Fungal Biol. 2011, 115, 643–648. [Google Scholar] [CrossRef]
- Olsson, P.l.A.; Hammer, E.C.; Wallander, H.k.; Pallon, J. Phosphorus availability influences elemental uptake in the mycorrhizal fungus Glomus intraradices, as revealed by particle-induced X-ray emission analysis. Appl. Environ. Microbiol. 2008, 74, 4144–4148. [Google Scholar] [CrossRef] [PubMed]
- Bücking, H.; Heyser, W. Elemental composition and function of polyphosphates in ectomycorrhizal fungi—An X-ray microanalytical study. Mycol. Res. 1999, 103, 31–39. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hijikata, N.; Ohtomo, R.; Handa, Y.; Kawaguchi, M.; Saito, K.; Masuta, C.; Ezawa, T. Aquaporin-mediated long-distance polyphosphate translocation directed towards the host in arbuscular mycorrhizal symbiosis: Application of virus-induced gene silencing. New Phytol. 2016, 211, 1202–1208. [Google Scholar] [CrossRef]
- Reddy, A.; Maley, F. Studies on identifying the catalytic role of Glu-204 in the active site of yeast invertase. J. Biol. Chem. 1996, 271, 13953–13957. [Google Scholar] [CrossRef]
- Liu, L.; Qu, J.; Wang, C.; Liu, M.; Zhang, C.; Zhang, X.; Guo, C.; Wu, C.; Yang, G.; Huang, J.; et al. An efficient genetic transformation system mediated by Rhizobium rhizogenes in fruit trees based on the transgenic hairy root to shoot conversion. Plant Biotechnol. J. 2024. [Google Scholar] [CrossRef]
Figure 1.
The phylogenetic tree of HAK/KUP/KT family of plant potassium transporters.

Figure 2.
Heterogeneous expression of LbHAK in yeast CY162 strain to compensate the K acquisition. (A) Growth assay for CY162 with empty vector (pYES2) and expressing LbHAK, and corresponding wild-type strain BY4741 (WT) on AP medium contained 0.5, 1, 5, and 10 mM KCl. (B) Growth kinetic analysis of CY162 with empty vector (pYES2) and expressing LbHAK, and corresponding wild-type strain BY4741 (WT) cultured in liquid AP medium with 0.5 mM KCl. Data indicate as mean ± SD.
Figure 2.
Heterogeneous expression of LbHAK in yeast CY162 strain to compensate the K acquisition. (A) Growth assay for CY162 with empty vector (pYES2) and expressing LbHAK, and corresponding wild-type strain BY4741 (WT) on AP medium contained 0.5, 1, 5, and 10 mM KCl. (B) Growth kinetic analysis of CY162 with empty vector (pYES2) and expressing LbHAK, and corresponding wild-type strain BY4741 (WT) cultured in liquid AP medium with 0.5 mM KCl. Data indicate as mean ± SD.

Figure 3.
Expression of LbHAK in L. barbarum roots in the split-roots system. 1 and 2 represent two split roots from one plant that were not treated with AM inoculation, 3 and 4 represents two split roots from one plant that one part was inoculated with R. irregularis or without R. irregularis, respectively (see detail in Figure S1, nd represents not detected).
Figure 3.
Expression of LbHAK in L. barbarum roots in the split-roots system. 1 and 2 represent two split roots from one plant that were not treated with AM inoculation, 3 and 4 represents two split roots from one plant that one part was inoculated with R. irregularis or without R. irregularis, respectively (see detail in Figure S1, nd represents not detected).

Figure 4.
Effects of overexpression of LbHAK ((A), shoot; (B), root; (C), total) on tobacco growth. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
Figure 4.
Effects of overexpression of LbHAK ((A), shoot; (B), root; (C), total) on tobacco growth. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).

Figure 5.
Evaluation of AM colonization efficiency in tobacco plants overexpressing LbHAK. (A) AM colonization rate; (B) arbuscular rate; GintEF1α (C) and NtPT4 (D) relative expression in mycorrhizal tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; nd, not detected. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
Figure 5.
Evaluation of AM colonization efficiency in tobacco plants overexpressing LbHAK. (A) AM colonization rate; (B) arbuscular rate; GintEF1α (C) and NtPT4 (D) relative expression in mycorrhizal tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; nd, not detected. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).

Figure 6.
Overexpression of LbHAK improved potassium contents ((A), shoot; (B), root; (C), total) in tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
Figure 6.
Overexpression of LbHAK improved potassium contents ((A), shoot; (B), root; (C), total) in tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).

Figure 7.
Overexpression of LbHAK improved phosphorus contents ((A), shoot; (B), root; (C), total)) in tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
Figure 7.
Overexpression of LbHAK improved phosphorus contents ((A), shoot; (B), root; (C), total)) in tobacco plants. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).

Figure 8.
Expression levels of Rir-AQP1 (A), and Rir-AQP2 (B) in wild-type and LbHAK-overexpressing tobacco with or without AM treatments. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; nd, not detected. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
Figure 8.
Expression levels of Rir-AQP1 (A), and Rir-AQP2 (B) in wild-type and LbHAK-overexpressing tobacco with or without AM treatments. Low potassium: 0.2 mM kg−1 substrate; High potassium: 2 mM kg−1 substrate; WT, wild-type tobacco plants; OE, tobacco plants overexpressing LbHAK; NM, non-mycorrhizal; AM, inoculated with R. irregularis; nd, not detected. Bars indicate Mean ± SD (n = 4). Different letters above the bars indicate significant difference at p < 0.05 (Duncan test).
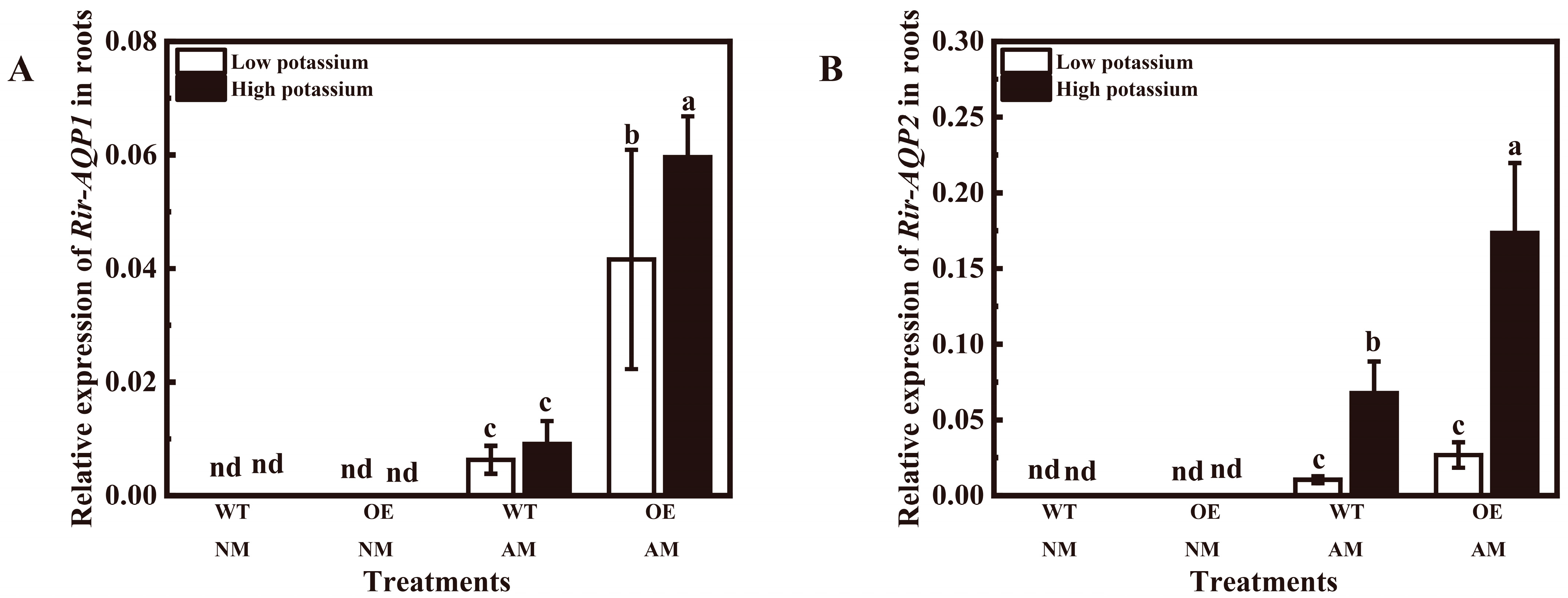
Figure 9.
Correlation analysis of gene expression, root nutrient contents, and colonization rate in tobacco roots and R. irregularis. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Figure 9.
Correlation analysis of gene expression, root nutrient contents, and colonization rate in tobacco roots and R. irregularis. *, p < 0.05; **, p < 0.01; ***, p < 0.001.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Bioinformation analysis of LbHAK in L. barbarum.
| Gene Name | GenBank Accession NO. | ORF Length (bp) | Protein Length (bp) | Predicted Molecular Weight (kDa) | Isoelectric Point | Predicted Subcellular Location |
|---|---|---|---|---|---|---|
| LbHAK | MZ416922.1 | 2004 | 668 | 97.35 | 9.07 | Plasma membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Han, X.; Ren, W.; Zhang, H.; Tang, M. Arbuscular Mycorrhizal Fungi Improve Lycium barbarum Potassium Uptake by Activating the Expression of LbHAK. Plants 2024, 13, 1244. https://doi.org/10.3390/plants13091244
AMA Style
Zhang Y, Han X, Ren W, Zhang H, Tang M. Arbuscular Mycorrhizal Fungi Improve Lycium barbarum Potassium Uptake by Activating the Expression of LbHAK. Plants. 2024; 13(9):1244. https://doi.org/10.3390/plants13091244
Chicago/Turabian StyleZhang, Yongxin, Xia Han, Wei Ren, Haoqiang Zhang, and Ming Tang. 2024. "Arbuscular Mycorrhizal Fungi Improve Lycium barbarum Potassium Uptake by Activating the Expression of LbHAK" Plants 13, no. 9: 1244. https://doi.org/10.3390/plants13091244
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.