
8q24.21 Locus: A Paradigm to Link Non-Coding RNAs, Genome Polymorphisms and Cancer



Department of Molecular Physiology and Cell Signalling, Institute of Systems, Molecular and Integrative Biology, University of Liverpool, Crown Street, Liverpool L69 3BX, UK
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2021, 22(3), 1094; https://doi.org/10.3390/ijms22031094
Submission received: 19 December 2020
/
Revised: 15 January 2021
/
Accepted: 20 January 2021
/
Published: 22 January 2021
(This article belongs to the Special Issue The Role of Polymorphic Variation in Non-coding DNA in Predicting Disease)
Abstract
:The majority of the human genome is comprised of non-protein-coding genes, but the relevance of non-coding RNAs in complex diseases has yet to be fully elucidated. One class of non-coding RNAs is long non-coding RNAs or lncRNAs, many of which have been identified to play a range of roles in transcription and translation. While the clinical importance of the majority of lncRNAs have yet to be identified, it is puzzling that a large number of disease-associated genetic variations are seen in lncRNA genes. The 8q24.21 locus is rich in lncRNAs and very few protein-coding genes are located in this region. Interestingly, the 8q24.21 region is also a hot spot for genetic variants associated with an increased risk of cancer. Research focusing on the lncRNAs in this area of the genome has indicated clinical relevance of lncRNAs in different cancers. In this review, we summarise the lncRNAs in the 8q24.21 region with respect to their role in cancer and discuss the potential impact of cancer-associated genetic polymorphisms on the function of lncRNAs in initiation and progression of cancer.
1. Introduction
Until recently, proteins and DNA were considered the main functional molecules of cells, with RNA being only a messenger between the two. This is the main reason why it was quite surprising to find that merely 2% of the human genome codes for proteins while 98% is non-protein coding. Since the human genome was first sequenced in 2003 [1], the biggest puzzle has been to understand the necessity of the 98% non-coding genome. Recent advances in sequencing technologies, however, have revealed that a large majority of the non-coding genome is transcribed into RNA transcripts. These non-coding RNAs, or ncRNAs, if functional, can explain the mystery of the non-coding human genome. However, functions of a large majority of the ncRNAs remain unknown, raising the possibility that they are merely a result of transcriptional noise in the cell. On the other hand, a number of observations support the functional importance of non-coding RNAs. For example, the observation that nearly 90% of all disease-associated genomic variations are within non-coding transcripts points towards their functional role [2]. However, a detailed analysis of genetic variations in the ncRNAs and their significance in diseases is needed. In this review, we will discuss disease-associated single-nucleotide variations in long non-coding RNAs or lncRNAs that are expressed from the 8q24.21 locus and their implications in cancers.
2. LncRNAs and Their Functions
LncRNAs are non-coding RNA transcripts that are more than 200 nucleotides in length [3]. With over 50,000 lncRNAs currently annotated and more still being discovered, the number of lncRNAs is significantly higher than that of protein-coding genes in the human genome [4]. LncRNAs are transcribed from stretches of DNA sequence located between genes (intergenic lncRNAs) or from introns of protein-coding genes (intronic lncRNAs). Much of the current research on lncRNAs has focused on intergenic lncRNAs or lincRNAs due to the difficulty of deciphering the function of intronic lncRNAs from the protein-coding gene that they reside in [5].
Although the functionality of a large majority of lncRNAs still remains unknown, examples suggest that lncRNAs can participate in diverse functions in the nucleus and cytoplasm, playing roles from transcription to translation (Figure 1). Often lncRNAs are located within the nucleus, where they are involved in chromatin remodelling, and transcriptional and post-transcriptional regulation [6]. Some lncRNAs are also exported to the cytoplasm. Within the cytoplasm, lncRNAs regulate translation, mRNA turnover and post-transcriptional modifications [6]. Some of the well-characterised lncRNAs, such as the X chromosome inactivation regulator Xist, can in fact co-ordinate multiple functions [7].
Within the nucleus, lncRNAs can act to regulate transcription through binding and sequestering proteins to prevent them from binding to their target RNA [8]. One set of proteins which can be modulated by lncRNAs are transcription factors. The lncRNA linc-YY1 has been shown to act as an RNA decoy for the transcription factor yin yang 1 or YY1, preventing the protein from binding to target promoters [9]. In addition, lncRNAs have been shown to bind to chromatin modifiers to alter transcription. Depending on the chromatin complex, lncRNAs can act to either repress or activate genes. For example, Kcnq1ot1 has been shown to repress transcription through binding and recruiting the DNA methyltransferase DNMT1, resulting in increased DNA methylation [10]. Contrastingly, the lncRNA HOTTIP binds to an adaptor protein to promote histone H3 lysine 4 trimethylation and gene transcription [11].
LncRNAs can also act as protein scaffolds and form ribonucleoproteins. In the nucleus, these complexes can regulate gene transcription. For example, lincRNA-p21 recruits the heterogeneous nuclear ribonucleoprotein-K or hnRNP-K to the promoter region of p21 and activates transcription [12]. In the cytoplasm, interactions between lncRNAs and ribonucleoproteins can regulate translation. For example, lincRoR interacts with hnRNP-I and supresses p53 translation by preventing the hnRNP-I from interacting with p53 mRNA [13].
In the cytoplasm, lncRNAs can modify gene expression post-transcriptionally through various mechanisms. LncRNAs can bind to microRNAs or miRNAs through complementary base pairings. This prevents miRNA from binding to target mRNAs, therefore enhancing expression of these genes by increasing translation and/or stability of mRNA. For example, the lncRNA SNHG7 has been identified to sponge miR-216 resulting in increased expression of the miR-216 target gene GALNT1 [14]. LncRNAs can also mediate mRNA decay though interacting with RNA-binding proteins. An example is a group of identified lncRNAs called half(½)-sbsRNAs which can bind to mRNA and form a functional Staufen 1-binding site, therefore triggering Staufen 1-mediated mRNA decay [15]. To regulate genes post-transcriptionally, lncRNAs can influence splicing events. MALAT1 has been shown to influence mRNA splicing by interacting with serine/arginine splicing factors and influencing the distribution of these splicing factors [16]. Finally, lncRNAs can act as translation inhibitors through binding to proteins associated with translation regulation. For example, lincRNA-p21 inhibits translation of target mRNAs through the recruitment of the translation repressor Rck [17], while AS Uchl1 acts as a translation activator of Uchl1 via an embedded SINEB2 element [18].
3. LncRNAs, GWAS and Cancer
Cancer is often described as a genetic disease arising from dysfunction of genes which are responsible for normal cell division and growth. Somatic mutations during one’s lifetime are believed to underlie the development of most cancers [20]. In a proportion of cancers, hereditary mutations confer a greater risk for the disease. In these cases, while some high-penetrance cancer susceptibility genes have been identified, such as BRCA1 for breast cancer, it is believed that genetic susceptibility in the majority of cases is a result of combined effects of common low-penetrance alleles and rare disease-causing variants [21]. Genome-wide association studies or GWAS have helped to identify novel genes associated with cancer, which has in turn helped improve not only our understanding of the disease but also drive forward new therapeutic advances related to population screening and pharmacological approaches.
Most causal variants associated with cancer risk identified through GWAS are located within non-coding regions of the genome [21]. Since the identification of lncRNAs, emerging evidence has begun to suggest that lncRNAs play a pivotal role in cancer development. Oncogenes are mutated genes which drive cancer initiation and progress. Under normal conditions, these genes are termed protooncogenes and are responsible for cell division and growth [22]. Like protooncogenes, lncRNAs have been proposed to regulate key cellular mechanisms associated with tumourgenesis including cell proliferation, apoptosis, migration and invasion [23]. For example, the lncRNA HOTAIR has been shown to promote proliferation and invasion of cancer cells in part through the regulation of p53 transcriptionally and SOX2 post-transcriptionally [24]. LncRNAs also have a role in epigenetic regulation including DNA methylation and histone modification which are important processes in cancer initiation [25]. In this role, HOTAIR can recruit histone-modifiers and chromatin remodelling complexes to regulate gene transcription [26]. In breast cancer, the interaction of HOTAIR with the Polycomb Repressive Complex 2 results in changes in histone methylation and gene expression associated with an increase in cancer cell metastasis [27]. HOTAIR highlights how a lncRNA can have a multi-faceted role in cancer development.
Investigating the role of specific lncRNA in cancer phenotypes is therefore a popular area of research. The identification of causal variants within non-coding loci has helped highlight potentially important lncRNAs related to cancer development.
4. 8q24.21
The human genome locus, 8q24.21, has been previously described as a “gene desert” due to sparse presence of protein-coding genes in this 4.1 MB region [28] (Figure 2). Despite this, genetic variation within this area has been associated with numerous cancer phenotypes. Lying within this region is one of the most studied oncogenes MYC, which is estimated to be involved in 20% of human cancers [29]. MYC primarily acts as a transcription factor and is shown to regulate cell cycle, metabolism, ribosome biogenesis and cell adhesion [29]. Numerous genetic polymorphisms associated with cancer risk are located within the non-coding regions surrounding MYC, which has led to characterising these regions as regulatory elements influencing MYC expression [30]. Other protein-coding genes within this region have also been shown to contribute to tumourigenesis [31,32,33,34]. These are FAM84B, GSDMC, FAM49B, and ASAP1. In addition, the OCT4 pseudogene POU5F1B is also present in the 8q24.21 region and has been showed to be amplified in cancer [35]. While POU51B has been referred to as a pseudogene, the latest human genome assembly GRCh38 from the Genome Reference Consortium states that POU5F1B is protein coding. However, in addition to the oncogenic protein-coding genes, the region 8q24.21 is a host to numerous lncRNAs associated with various cancers and with functions that are independent of MYC.
A total of 12 lncRNAs are located within the 8q24.21 region (Figure 2). Upstream of the protein-coding gene MYC are the lncRNAs PCAT1, CASC19, PRNCR1, CCAT1, CASC8, CCAT2 and CASC11. Downstream of MYC lies PVT1, LINC00924, and CCDC26. The majority of the lncRNAs located within 8q24.21 can be categorised as lincRNAs as they are located in intergenic regions between the five protein-coding genes in this region. The exception is CASC8, which is anti-sense to the protein-coding gene POU5F1B. In addition to lncRNAs, there are also several microRNAs or miRNAs located within the region. Five miRNAs have been shown to arise from the PVT1 locus. These are miR-1204, miR-1205, miR-1206, miR-1207 and miR-1208. In addition, miR-3686 is located within CCDC26, with miR-5194 located downstream of CCDC26.
Numerous genetic variants associated with increased risk of cancer are located within the lncRNAs in the 8q24.21 locus. According to the NHGRI-EBI GWAS Catalog [36], a total of 324 single-nucleotide polymorphisms or SNPs have been reported in the 8q24.21 region. Interestingly, to date, only four SNPs have been located within the MYC locus and none of these SNPs are associated with cancer. Contrastingly, several of the lincRNAs in this region appear to be hotspots for SNPs including PCAT1 and CCDC26 which are associated with 73 and 81 SNPs, respectively, with many of these SNPs associated with various cancers. While it is possible that SNPs located in the non-coding regions may influence protein-coding genes, they also highlight the potential importance of the 8q24.21 lncRNAs in disease [37]. Therefore, unsurprisingly, many studies have focused on the 8q24.21 region to help elucidate the role of lncRNAs in the initiation and progression of cancer in the hope of identifying novel biomarkers and new therapeutic targets. In this review, as a paradigm, we summarise the potential impact of genetic polymorphisms on tumourigenic functions 8q24.21 lncRNAs.
4.1. 8q24.21 LncRNA Expression Changes in Cancer
Relative to protein-coding RNA, the expression levels of lncRNA are low [38]. However, lncRNAs are often found to be elevated cancer and contribute to the transition of normal cells into tumour cells [39]. The lncRNAs located within the 8q24.21 locus have been shown to be upregulated in a number of cancers including colon cancer, pancreatic cancer, glioma and leukaemia (Table 1). While the functional significance of the increased expression is not fully known, upregulation of these lncRNAs often correlated with poorer overall survival and a more aggressive cancer phenotype. Therefore, high expression of the lncRNAs appears to be an independent prognosis factor for advanced cancer and poorer prognosis in various cancer subtypes. The impact of the lncRNAs in promoting a tumourigenic phenotype is attributed to their ability to drive proliferation, migration and invasion of cancer cells. Silencing of these lncRNA genes was found to supress cell growth and apoptosis in cancer cells. Targeting these lncRNAs therefore appears to be a promising therapeutic approach for many cancers.
4.2. 8q24.21 LncRNAs in miRNA Regulation
Numerous binding sites of miRNAs have been located within the lincRNAs of the 8q24.21 locus. MiRNAs are small ncRNAs which are an average of 22 nucleotides in length. The main function of miRNAs is in gene silencing, in which they bind to mRNAs, predominantly the 3′UTR region, and supress expression [117]. MiRNAs have been shown to regulate tumourigenesis through the suppression of oncogenes and tumour suppressor genes, and dysregulation of miRNAs is evident in various cancers [118]. LncRNAs have been shown to act as a molecular sponge to sequester miRNAs and prevent their interaction with target messenger RNAs. Studies have identified complementary base pair regions between the majority of the lincRNAs located in 8q24.21 (PVT1 [119], CCAT1 [120], PCAT1 [121], CCAT2 [122], CASC11 [123], PRNCR1 [52], CASC19 [50], CCDC26 [116]) and miRNAs which have been previously identified as tumour suppressors. Within this role, many of these lincRNAs can act as competitive endogenous RNA or ceRNA for miRNA target genes (Table 2). Like other miRNA target genes, lncRNA expression changes are often associated with the initiation and/or progression of cancer. A good example of this is the 8q24.21 lincRNA CCAT2, which was found to act as a ceRNA, in glioma, for miR-424, which also targets VEGFA. In this case, subsequent activation of VEGFA signalling promotes angiogenesis [89].
4.3. 8q24.21 LncRNAs and Chromatin Modifications
LncRNAs can often modulate gene expression through changes in chromatin modifications such as DNA and histone methylation. One such example is that of lincRNA CCDC26, which is expressed from a locus approximately 1MB downstream of MYC. Within numerous cancer lines, CCDC26 has been shown to regulate cell growth and apoptosis [116,143,144], and be enriched within nuclear fraction of myeloid leukaemia cells [144]. We have further explored the role of CCDC26 in apoptosis in relation to DNA methylation. We found that CCDC26 interacts with the methyltransferase DNMT1 and drives changes in methylation levels in genomic DNA [144]. Inhibition of CCDC26 results in the hypomethylation of the genome, leading to inhibition of cell growth and an increase in apoptosis associated with DNA damage. While DNMT expression levels remained unchanged following CCDC26 removal, DNTM1 was found to be mis-localised in cytoplasm which is the likely cause for the observed genomic hypomethylation [144]. Our results suggest that CCDC26 influenced subcellular localisation of DNMT1 through protein–RNA interaction in the nucleus [144].
Overexpression of another lincRNA PVT1 in 8q24.21 resulted in increased levels of the methyltransferases DNMT1, DNMT3a and DNMT3b and promoted the methylation CpG islands located in the miRNA miR-146a promoter region [145]. The methylation of the promoter region resulted in the suppression of miR-146a expression. While the role of miR-146a in cancer was not studied further, significantly low expression of miR-146a was found in prostate cancer cells compared to normal tissue, suggesting a role in tumour development [145]. In addition, lincRNAs PVT1 and PCAT1 have been shown to directly bind to the histone methyltransferase EZH2 and induce trimethylation of lysine 27 on histone H3, H3K27me3, which leads to transcriptional repression. PVT1 induces H3K27me3 modification at the miR-200b promoter, leading to silencing of miR-200b [146]. Mir-200b has been shown to inhibit proliferation and migration of cancer cells, and is dysregulated in numerous cancers [146]. Similarly binding of EZH2 to PCAT1 is needed for PCAT1-associated silencing of E-cadherin and p21 [147,148]. LincRNA CCAT2 has also been shown to bind directly to EZH2 as well as post-transcriptionally downregulate EZH2 expression [149].
4.4. 8q24.21 LncRNAs and c-Myc Regulation
Many studies have investigated the link between the lncRNAs located on 8q24.21 and the oncogene MYC and the protein it encodes c-Myc, especially lncRNAs PVT1 and PCAT1, which lie in a close proximity to MYC. Co-amplification of PVT1 and MYC is evident in numerous cancers [150]. Studies have suggested that PVT1 and MYC work synergistically in the regulation of proliferation [151]. Indeed, many of PVT1 targets are also downstream signalling molecules of c-Myc, strengthening the idea that they both work together to drive cancer progression [151]. PVT1 is involved in the transcriptional and post-transcriptional regulation of c-Myc [151,152,153]. In addition, PVT1 itself is a target for c-Myc. Within the PVT1 promoter region, two enhancer E-box sites have been identified in which c-Myc binds to transcriptionally activate PVT1 [154]. Therefore, it is possible that a positive feedback mechanism exists between PVT1 and c-Myc, in which c-Myc promotes PVT1 expression and then PVT1 prevents the degradation of c-Myc, driving the expression of both oncogenes in cancer.
Contrastingly, a study has found that silencing the PVT1 promoter results in increased cell proliferation and competition in breast cancer cells, which was associated with an increase in MYC expression [155]. The study identified that four intragenic enhanced elements have been identified in the PVT1 gene which bind to the PVT1 promoter in a preferential manner over MYC. Silencing of the PVT1 promoter resulted in these enhancer elements making stronger connections with the MYC promoter and enhancer region resulting in increased MYC expression. Altogether, the study proposed that the PVT1 promoter can act to repress MYC transcription in a cis manner [155].
On a post-transcriptional level, PCAT1 has also been found to regulate c-Myc. Overexpression of PCAT1 resulted in the post-transcriptional upregulation of c-Myc [156]. PCAT1 was also shown to interfere with miRNA-mediated degradation of MYC. The upregulation of c-Myc by PCAT1 promotes cell proliferation and the expression of genes involved in protein biosynthesis and transcriptional elongation [156]. Supporting this, the knockdown of PCAT1 also resulted in reduced cell proliferation associated with downregulation of the c-Myc/MAPK signalling pathway [45].
In addition to PVT1 and PCAT1, other lncRNAs from the 8q24.21 locus are shown to regulate MYC expression. LincRNA CASC11, for example, transcriptionally regulates MYC expression by modulating a MYC enhancer that is 557 kb upstream of MYC. The binding occurred in a PCBP2-dependent manner, with CASC11 and the RNA-binding protein PCBP2 forming a looping structure between the MYC promoter and enhancer [157]. Similarly, lincRNA CCAT1 has been shown to be required for the maintenance of chromatin loops at the MYC locus in colorectal cancer. The study suggested that CCAT1 interacts with CTCF, a transcriptional regulator protein, and regulates the binding of CTCF to chromatin, which mediates the long-range interactions between MYC promoter and enhancers [158].
Therefore, targeting lncRNAs which modulate c-Myc activity proposes a novel approach to treating Myc-associated cancers.
5. Potential Impact of Genetic Polymorphisms on 8q24.21 LncRNAs
5.1. Amplifications
Amplifications within the 8q24.21 region have been detected in various forms of cancer. This genomic site is known as one of the most prevalent sites of copy number gains in cancer [159,160]. The presence of copy number gains in this region could result in upregulation of lncRNAs and promote abnormal cell growth of cells (Figure 3A). In a study focusing on ovarian cancer, gains localised to 8q24.21 were evident in 59% of samples and resulted in a significant overexpression of PVT1, suggesting gains within this region can be a crucial factor of elevating lncRNAs in cancer [161,162]. In addition, copy number variants have been shown to drive co-amplification of PVT1 and MYC in numerous cancers [152]. Analysis of The Cancer Genome Atlas database found that out of the tumours which displayed an 8q24 copy-number increase, 97% showed a gain of both PVT1 and MYC, while less than 1% of tumours showed an increase in copy number of MYC alone [152]. This gain of both PVT1 and MYC has been shown to be essential for tumourigenesis in 8q24-amplified cancer cells. In the study, gain of MYC or the PVT1/CCDC26/GSDMC region alone did not result in tumourigenesis, but co-amplification of MYC and PVT1/CCDC26/GSDMC resulted in a pro-tumour transformation. Specifically, the PVT1 copy number increase was found to be the critical factor for MYC, and subsequent c-Myc, elevation in MYC-driven cancers [152].
In addition to PVT1, amplifications in other 8q24.21 lncRNAs have been identified. In studies investigating paediatric acute myeloid leukaemia, the most common copy number alteration was found to be a low burden increase in the CCDC26 locus [163]. In castration-resistant prostate cancer samples, duplications hotspots were evident in the region hosting PCAT1 and PRNCR1 [164]. While the study did not investigate expression of the lncRNAs, it is possible that these events lead to amplification of these genes.
5.2. Structural Variations
Chromosomal instability is a source of genetic variation and is a common feature of tumours. Often this leads to chromosomal rearrangements including translocations [165]. Within the 8q24.21 loci, a common site of translocation is within PVT1. PVT1 was initially identified as a site for variant translocation involving the Ig kappa locus and PVT1 gene in murine plasmocytomas [166], with the equivalent translocation observed in Burkitt’s lymphoma in human [167]. Since then, recurrent intra-chromosomal inversions within intron 1 of PVT1 have been identified in breast cancer patients [155]. Similar to breast cancer, formation of PVT1 fusion proteins as a result of translocations occurring in intron 1 of the gene is also evident in numerous other cancers (Figure 3B) [155,168,169]. The promoter region of PVT1 appears to be critical to the function of the lincRNA, especially in relation to c-Myc regulation. The intra-chromosomal inversion, in addition to forming a fusion protein, is also thought to affect the PVT1 promoter region by separating it from intragenic enhancers [155]. PVT1 has also been identified to fuse with MYC [150], CCDC26 [161], NSMCE2 [170], and NDRG1 [171]. While the specific effects of these gene fusions are unclear, it is possible that they have functions distinct from the individual genes. For example, in a breast cancer model, gain of both PVT1 and MYC resulted in an increased proliferation of cells which was not evident by increase in PVT1 and MYC alone [172].
Other genomic rearrangements which often occur alongside amplification events include the formation of double-minute chromosomes. These extrachromosomal fragments of DNA consist of repeats of specific chromosomal regions termed an amplicon. Amplification of double minutes are believed to increase the copy number of oncogenes to drive tumour heterogeneity and are associated with treatment resistance [173]. For example, in acute myeloid leukaemia patients, 8q24 chromosome is often amplified by forming double-minute chromosomes [174,175]. Surprisingly, evidence suggests that MYC is not the target gene for this type of amplification due to the gene often being silent in these subsets of acute myeloid leukaemia cases [174]. Therefore, it is possible that the target of the amplicon is the lncRNAs residing within this region. Supporting this idea, mapping of the 8q24.21 amplicon in acute myeloid leukaemia has identified alterations in the structure of lincRNA CCDC26 as a result of recombination event upstream of exon 4 [175]. It is unknown whether this partial amplification of the gene results in a gain or loss of function, but it is likely abnormalities in CCDC26 structure drive a pro-tumour phenotype.
5.3. Single-Nucleotide Polymorphisms
Despite a large number of cancer-associated SNPs being located within lncRNAs in the 8q24.21 locus, studies have yet to identify the impact of these genetic polymorphisms on the functions of the lncRNAs. One SNP that has been explored is rs698267, located within CCAT2, and it is highly prevalent in patients with myelodysplastic syndrome [149]. The CCAT2 SNP was shown to result in a specific RNA editing event causing CCAT2 DNA-to-RNA allelic imbalance, with the RNA transcribed from the SNP locus being different compared to corresponding genomic DNA. The cancer-predisposing allele resulting from this SNP resulted in gene expression dysregulation and repression of EZH2 function [149]. Interestingly, of the patients carrying the SNP, 78% expressed heterozygous CCAT2 RNA. The study found that the presence of both the normal and SNP-carrying CCAT2 allele resulted in a combined oncogenic effect and a stronger downregulation of EZH2 than that achieved by homozygous expression of either allele [149]. This phenomenon is believed to be due to the specific alleles independently promoting a pro-tumour phenotype through regulating distinct pathways [149]. The same SNP has also been investigated in relation to cancer metabolism [176]. In this study, the SNP was shown to induce changes in the secondary structure of CCAT2 and affect the binding of CCAT2 to the CFIam protein. This resulted in alternative splicing of GLS to preferentially induce the expression of the oncogenic isoform [176].
The impact of SNPs on 8q24.21 lncRNAs can be guessed based on the position of the SNPs within the lncRNA genes. Studies investigating lncRNA SNPs in other areas of the genome have shown that SNPs can influence the expression of lncRNAs (Figure 3A). A cancer-associated SNP located with the lncRNA PTCSC3 reduces the binding affinity of the transcription factor C/EBPα, resulting in decreased PTCSC3 promoter activation and reduced expression of the lncRNA which is evident in thyroid carcinoma [177]. Within the 8q24.21 region, a large number of cancer-associated SNPs are clustered within the first intron of CCDC26 gene. On the background that the main transcriptional start site of CCDC26 is located within exon 2 of the gene [144], we can speculate that cancer-associated SNP hotspot in the first intron may affect the promoter region of CCDC26 and therefore impact expression of the lincRNA. Similarly, SNPs associated with cancers have been shown to be located within intron 1 of PVT1 which can modify promoter strength of the gene [36].
In addition, a cancer-associated SNP located within the intronic region of the lncRNA PCAT19 has been found to regulate the isoforms of PCAT19 in a reciprocal manner. The SNP is associated with decreased levels of PCAT1-short and increased levels of PCAT1-long, with the long isoform associated with prostate cancer progression [178]. Many of the lncRNAs located in the 8q24.21 region have numerous different isoforms. While the functional role of the varied isoforms has not been assessed, it is possible that SNPs within the intronic regions of these lncRNAs could mediate a switch between isoforms in cancer (Figure 3C).
SNPs can also impact the structure of lncRNAs and their ability to form RNA and protein interactions. Studies have begun to look at this concept by using computational modelling to predict the impact of SNPs on lncRNA function. A lung cancer-associated SNP located in exon 2 of NEXN-1 has been predicted to change the secondary structure of the lncRNA, which may influence the ability of NEXN-1 to interact with proteins [179]. In addition, a SNP located within the intronic region of HOTAIR has been shown to alter the binding affinity of the lncRNA to transcription factors associated with cancer including PAX-4, SOX, SPZ1 and ZFP281 [180]. In terms of 8q24.21 lncRNAs, no studies have yet investigated the impact of SNPs on their ability to bind to target protein. However, it is probable that SNPs influence the function of 8q24.21 lncRNA in gene regulation, notably in relation to MYC (Figure 3D).
Cancer-associated SNPs have also been identified within miRNA binding sites of lncRNAs [181]. These SNPs can interfere or enhance hybridisation of the lncRNAs with miRNAs. A SNP located in the lncRNA MALAT1 has been shown to inhibit binding of the lncRNA to miR-194-5p, resulting in increased expression of MALAT1 and contributing to colorectal cancer development [182]. SNPs can also result in new binding sites for miRNA. A SNP identified in CCSlnc362 which confers a protective role from colorectal cancer was shown to result in a new miRNA binding site for miR-4658 in exon 1 of the lncRNA. Binding of miR-4658 reduced expression and supressed the role of CClnc362 in tumour progression [183]. While these studies have focused on the role of miRNAs on the expression of lncRNAs, it is possible that SNPs within miRNA binding domains can impact the ability of lncRNAs to sponge miRNAs, a feature common to 8q24.21 lncRNAs. In this role, SNPs could result in novel lncRNA-mediated miRNA sponging events or prevent lncRNA:miRNA binding, resulting in increased expression of cancer-associated miRNAs (Figure 3E).
In complex diseases such as cancer, the underlying genetic cause is usually a result of multiple genetic polymorphisms. SNPs have been shown to co-occur with other SNPs, and it is likely that this occurs in the 8q24.21 region. While individual SNPs generally confer a small effect, the combined effect of numerous relevant SNPs is thought to result in a synergistic increased risk. In addition, co-occurrence of SNPs can lead to SNP–SNP interactions [184]. A study investigating pancreatic cancer risk identified numerous SNP–SNP interactions involving SNPs located in the 8q24.21 region. Interestingly, a SNP located in CASC11 was involved in 70% of the SNP–SNP interaction pairs significantly associated with pancreatic cancer risk, with the most common genetic interaction being between PVT1 and CASC11 [185]. Although a direct link between the two lncRNAs still remains missing, it is thought that PVT1 and CASC11 are linked through MYC [91,185,186].
6. Future Work and Limitations
Genetic polymorphisms are a frequent feature of cancer. While not necessarily of greater importance compared to other cancer polymorphisms, 8q24.21 can serve as an excellent paradigm for understanding the role of lncRNAs and non-coding genetic polymorphisms in oncogenesis. A possible approach to assessing the importance of 8q24.21 lncRNAs is through the use of animal models. However, there are limitations. Firstly, the majority of the lncRNAs in the 8q24.21 locus are not evolutionary conserved, and there are no mouse orthologs for these lncRNAs. Despite this, some parts of the non-coding region of 8q24.21 have been evaluated in mouse models [187]. One study generated a mouse model lacking a 430 kb gene desert region between the protein-coding genes FAM84B and MYC. This region had previously been associated with breast cancer susceptibility in humans. The study found that deletion of this region resulted in an anti-tumourigenic effect in early and late stages of mammary cancer in mouse breast cancer models [187]. Interestingly, deletion of the region resulted in a decreased expression of MYC, located approximately 200 kb from the deleted region. The study further identified a chromatin loop conserved between mouse and human from the MYC promoter to the deleted region [187]. These findings suggest that the 8q24.21 gene desert region does contain regulatory elements which are conserved in both humans and mice. The evolutionary relation of these conserved regions with lncRNAs found in the human 8q24.21 region needs to be scrutinised.
While animal models have been effective in the study of some evolutionary conserved lncRNAs, some discrepancies between human and animal data have raised questions regarding functional significance of lncRNAs. For example, the evolutionary conserved, cancer-associated lncRNA MALAT1 has been suggested to regulate gene transcription in human cells. However, mouse models lacking MALAT1 have shown no obvious phenotypes and had no apparent changes in global gene expression or pre-mRNA splicing [188]. Altered expression of MALAT1 has been described in many human cancers, with in vitro studies indicating that MALAT1 promotes tumour growth. Studies investigating reduced MALAT1 expression in mouse models of cancer have found conflicting results depending on the strategy of MALAT1 inactivation [188]. Using the MMTV-PyMT (mouse mammary tumour virus-polyoma middle tumour antigen) cancer model, knockout of a 3 kb portion of MALAT1 promoter region and the lncRNAs 5′ end resulted in a reduction in lung metastases. On the other hand, using the same MMTV-PyMT model but with targeted insertional inactivation of MALAT1 resulted in an increase in metastatic sites in the lungs [188]. The work involving MALAT1 highlights the limitations in current understanding and the need for robust characterisation of lncRNA evolution and function.
7. Conclusions
Genetic polymorphisms associated with cancers are often located within the non-coding region of the genome. The 8q24.21 locus is the site of numerous lncRNAs and a hotspot of cancer polymorphisms. Studies have identified elevated expression of the 8q24.21 lncRNAs in numerous cancers associated with the progression of tumour phenotype. Indeed, in various studies, high levels of these lncRNAs have been identified as an independent prognostic factor for poor overall survival. The potential importance of these lncRNAs is evident by functional studies showing that, within cancer tissues and cells, the 8q24.21 lncRNAs sponge miRNAs drive chromatin modification and regulate the expression of the oncoprotein c-Myc. The 8q24.21 region of the genome is therefore an interesting region to explore the link between genetic variants, lncRNAs and cancer progression. While the advent in GWAS has highlighted the large number of cancer-associated variants within this region, research is still lacking on identifying the impact that these polymorphisms have on lncRNA function. We can begin to speculate the potential impact of genetic polymorphisms, including SNPs, based on previous studies investigating the impact of variants on lncRNA function. Further work to help bridge this knowledge gap is needed which will allow a clearer understanding of the functionality of lncRNAs in cancer and will boost the use of lncRNAs as novel targets in cancer therapeutics.
Author Contributions
A.K. and C.W. both contributed conceptually to the paper. C.W. completed the first draft of the manuscript, including the figures and tables. A.K. contributed to the final version of the manuscript and supervised all the work. Both authors reviewed the manuscript prior to submission. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the University of Liverpool.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Human Genome Sequencing Consortium. Finishing the euchromatic sequence of the human genome. Nature 2004, 431, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Giral, H.; Landmesser, U.; Kratzer, A. Into the Wild: GWAS Exploration of Non-coding RNAs. Front. Cardiovasc. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [PubMed]
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Djebali, S.; Davis, C.A.; Merkel, A.; Dobin, A.; Lassmann, T.; Mortazavi, A.; Tanzer, A.; Lagarde, J.; Lin, W.; Schlesinger, F.; et al. Landscape of transcription in human cells. Nature 2012, 489, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Bajic, V.B.; Zhang, Z. On the classification of long non-coding RNAs. RNA Biol. 2013, 10, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.-W.; Wang, Y.; Chen, L.-L. Cellular functions of long noncoding RNAs. Nat. Cell Biol. 2019, 21, 542–551. [Google Scholar] [CrossRef]
- Pintacuda, G.; Young, A.N.; Cerase, A. Function by Structure: Spotlights on Xist Long Non-coding RNA. Front. Mol. Biosci. 2017, 4, 90. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Chang, H.Y. Molecular mechanisms of long noncoding RNAs. Mol. Cell. 2011, 43, 904–914. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Sun, K.; Zhao, Y.; Zhang, S.; Wang, X.; Li, Y.; Lu, L.; Chen, X.; Chen, F.; Bao, X.; et al. Linc-YY1 promotes myogenic differentiation and muscle regeneration through an interaction with the transcription factor YY1. Nat. Commun. 2015, 6, 10026. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, F.; Mondal, T.; Guseva, N.; Pandey, G.K.; Kanduri, C. Kcnq1ot1 noncoding RNA mediates transcriptional gene silencing by interacting with Dnmt1. Development 2010, 137, 2493–2499. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.C.; Yang, Y.W.; Liu, B.; Sanyal, A.; Corces-Zimmerman, R.; Chen, Y.; Lajoie, B.R.; Protacio, A.; Flynn, R.A.; Gupta, R.A.; et al. A long noncoding RNA maintains active chromatin to coordinate homeotic gene expression. Nat. Cell Biol. 2011, 472, 120–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimitrova, N.; Zamudio, J.R.; Jong, R.M.; Soukup, D.; Resnick, R.; Sarma, K.; Ward, A.J.; Raj, A.; Lee, J.T.; Sharp, P.A. Lin-cRNA-p21 activates p21 in cis to promote Polycomb target gene expression and to enforce the G1/S checkpoint. Mol. Cell. 2014, 54, 777–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Haider, A.M.S.S.; Moran, M. The role of interactions of long non-coding RNAs and heterogeneous nuclear ribonu-cleoproteins in regulating cellular functions. Biochem. J. 2017, 474, 2925–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, Y.; Ma, J.; Pan, Y.; Hu, J.; Liu, B.; Jia, L. LncRNA SNHG7 sponges miR-216b to promote proliferation and liver metastasis of colorectal cancer through upregulating GALNT1. Cell Death Dis. 2018, 9, 722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, C.; Maquat, L.E. lncRNAs transactivate STAU1-mediated mRNA decay by duplexing with 3’ UTRs via Alu elements. Nature 2011, 470, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The Nuclear-Retained Noncoding RNA MALAT1 Regulates Alternative Splicing by Modulating SR Splicing Factor Phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Yoon, J.H.; Abdelmohsen, K.; Srikantan, S.; Yang, X.; Martindale, J.L.; De, S.; Huarte, M.; Zhan, M.; Becker, K.G.; Gorospe, M. Lin-cRNA-p21 suppresses target mRNA translation. Mol. Cell 2012, 47, 648–655. [Google Scholar] [CrossRef] [Green Version]
- Carrieri, C.; Cimatti, L.; Biagioli, M.; Beugnet, A.; Zucchelli, S.; Fedele, S.; Pesce, E.; Ferrer, I.; Collavin, L.; Santoro, C.; et al. Long non-coding antisense RNA controls Uchl1 translation through an embedded SINEB2 repeat. Nat. Cell Biol. 2012, 491, 454–457. [Google Scholar] [CrossRef]
- Lee, J.T. Epigenetic Regulation by Long Noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Sud, A.; Kinnersley, B.; Houlston, R.S. Genome-wide association studies of cancer: Current insights and future perspectives. Nat. Rev. Cancer 2017, 17, 692–704. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Muller, W.J. Oncogenes and tumor suppressor genes. Cold Spring Harb. Perspect. Biol. 2010, 2, a003236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, A.M.; Chang, H.Y. Long Noncoding RNAs in Cancer Pathways. Cancer Cell 2016, 29, 452–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, J.; Yang, M.; Jiang, R.; An, N.; Wang, X.; Liu, B. Long Non-Coding RNA HOTAIR Regulates the Proliferation, Self-Renewal Capacity, Tumor Formation and Migration of the Cancer Stem-Like Cell (CSC) Subpopulation Enriched from Breast Cancer Cells. PLoS ONE 2017, 12, e0170860. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Liu, G.; Zhou, F.; Su, B.; Li, Y. DNA methylation profiles in cancer diagnosis and therapeutics. Clin. Exp. Med. 2018, 18, 1–14. [Google Scholar] [CrossRef]
- Hanly, D.J.; Esteller, M.; Berdasco, M. Interplay between long non-coding RNAs and epigenetic machinery: Emerging targets in cancer? Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170074. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.-C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nat. Cell Biol. 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Huppi, K.; Pitt, J.J.; Wahlberg, B.M.; Caplen, N.J. The 8q24 Gene Desert: An Oasis of Non-Coding Transcriptional Activity. Front. Genet. 2012, 3, 69. [Google Scholar] [CrossRef] [Green Version]
- Dang, C.V.; O’Donnell, K.A.; Zeller, K.I.; Nguyen, T.; Osthus, R.C.; Li, F. The c-Myc target gene network. Semin. Cancer Biol. 2006, 16, 253–264. [Google Scholar] [CrossRef]
- Grisanzio, C.; Freedman, M.L. Chromosome 8q24-Associated Cancers and MYC. Genes Cancer 2010, 1, 555–559. [Google Scholar] [CrossRef]
- Gu, Y.; Lin, X.; Kapoor, A.; Chow, M.J.; Jiang, Y.; Zhao, K.; Tang, D. The Oncogenic Potential of the Centromeric Border Protein FAM84B of the 8q24.21 Gene Desert. Genes 2020, 11, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Xu, Z.; Chen, X.; Wang, X.; Zeng, S.; Qian, L.; Yang, X.; Ou, C.; Lin, W.; Gong, Z.; et al. Overexpression of GSDMC is a prognostic factor for predicting a poor outcome in lung adenocarcinoma. Mol. Med. Rep. 2019, 21, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattaragada, M.S.; Riganti, C.; Sassoe, M.; Principe, M.; Santamorena, M.M.; Roux, C.; Curcio, C.; Evangelista, A.; Allavena, P.; Salvia, R.; et al. FAM49B, a novel regulator of mitochondrial function and integrity that sup-presses tumor metastasis. Oncogene 2018, 37, 697–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, T.; Stein, U.; Poletti, A.; Garzia, L.; Rothley, M.; Plaumann, D.; Thiele, W.; Bauer, M.; Galasso, A.; Schlag, P.; et al. ASAP1 promotes tumor cell motility and invasiveness, stimulates metastasis formation in vivo, and correlates with poor survival in colorectal cancer patients. Oncogene 2010, 29, 2393–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, H.; Arao, T.; Togashi, Y.; Kato, H.; Fujita, Y.; De Velasco, M.A.; Kimura, H.; Matsumoto, K.; Tanaka, K.L.; Okamoto, I.; et al. The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene 2015, 34, 199–208. [Google Scholar] [CrossRef]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The NHGRI-EBI GWAS Catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hrdlickova, B.; de Almeida, R.C.; Borek, Z.; Withoff, S. Genetic variation in the non-coding genome: Involvement of micro-RNAs and long non-coding RNAs in disease. Biochim. Biophys. Acta 2014, 1842, 1910–1922. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Huarte, M. The emerging role of lncRNAs in cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Ge, X.; Chen, Y.; Liao, X.; Liu, D.; Li, F.; Ruan, H.; Jia, W. Overexpression of long noncoding RNA PCAT-1 is a novel biomarker of poor prognosis in patients with colorectal cancer. Med Oncol. 2013, 30, 588. [Google Scholar] [CrossRef]
- Yan, T.-H.; Yang, H.; Jiang, J.-H.; Lu, S.-W.; Peng, C.-X.; Que, H.-X.; Lu, W.-L.; Mao, J.-F. Prognostic significance of long non-coding RNA PCAT-1 expression in human hepatocellular carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 4126–4131. [Google Scholar] [PubMed]
- Zhang, F.; Wan, M.; Xu, Y.; Li, Z.; Leng, K.; Kang, P.; Cui, Y.; Jiang, X. Long noncoding RNA PCAT1 regulates extrahepatic chol-angiocarcinoma progression via the Wnt/beta-catenin-signaling pathway. Biomed Pharmacother. 2017, 94, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.-H.; Wu, Q.-Q.; Li, S.-Q.; Yang, T.-X.; Liu, Z.-H.; Tong, Y.-S.; Tuo, L.; Wang, S.; Cao, X.-F. Upregulation of the long noncoding RNA PCAT-1 correlates with advanced clinical stage and poor prognosis in esophageal squamous carcinoma. Tumor Biol. 2015, 36, 2501–2507. [Google Scholar] [CrossRef] [PubMed]
- Sarrafzadeh, S.; Geranpayeh, L.; Ghafouri-Fard, S. Expression Analysis of Long Non-Coding PCAT-1in Breast Cancer. Int. J. Hematol. Stem Cell Res. 2017, 11, 185–191. [Google Scholar]
- Sur, S.; Nakanishi, H.; Steele, R.; Ray, R.B. Depletion of PCAT-1 in head and neck cancer cells inhibits tumor growth and induces apoptosis by modulating c-Myc-AKT1-p38 MAPK signalling pathways. BMC Cancer 2019, 19, 354. [Google Scholar] [CrossRef] [Green Version]
- Hu, W.; Dong, N.; Huang, J.; Ye, B. Long non-coding RNA PCAT1 promotes cell migration and invasion in human laryngeal cancer by sponging miR-210-3p. J. BUON 2020, 24, 2429–2434. [Google Scholar]
- Zhang, X.; Zhang, Y.; Mao, Y.; Ma, X. The lncRNA PCAT1 is correlated with poor prognosis and promotes cell proliferation, invasion, migration and EMT in osteosarcoma. Onco Targets Ther. 2018, 11, 629–638. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Ge, Y.; Wang, Y.; Ma, G.; Wang, X.; Liu, H.; Wang, M.; Zhang, Z.; Chu, H. The association of rs710886 in lncRNA PCAT1 with bladder cancer risk in a Chinese population. Gene 2017, 627, 226–232. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Fu, C.; Wang, C.; Duan, X.; Zou, W.; Zhao, T. Knockdown of long non-coding RNA PCAT1 in glioma stem cells promotes radiation sensitivity. Med Mol. Morphol. 2018, 52, 114–122. [Google Scholar] [CrossRef]
- Wang, X.D.; Lu, J.; Lin, Y.S.; Gao, C.; Qi, F. Functional role of long non-coding RNA CASC19/miR-140-5p/CEMIP axis in colorectal cancer progression in vitro. World J. Gastroenterol. 2019, 25, 1697–1714. [Google Scholar] [CrossRef]
- Liu, Y.J.; Guo, R.X.; Han, L.P.; Gu, H.; Liu, M.Z. Effect of CASC19 on proliferation, apoptosis and radiation sensitivity of cervical cancer cells by regulating miR-449b-5p expression. Zhonghua Fu Chan Ke Za Zhi 2020, 55, 36–44. [Google Scholar] [PubMed]
- Guo, R.; Hu, T.; Liu, Y.; He, Y.; Cao, Y. Long non-coding RNA PRNCR1 modulates non-small cell lung cancer cell proliferation, apoptosis, migration, invasion, and EMT through PRNCR1/miR-126-5p/MTDH axis. Biosci. Rep. 2020, 40, BSR20193153. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Zou, Y.; Li, R.; Liu, D. Long noncoding RNA PRNCR1 exerts oncogenic effects in tongue squamous cell carcinoma in vitro and in vivo by sponging microRNA944 and thereby increasing HOXB5 expression. Int. J. Mol. Med. 2020, 46, 119–130. [Google Scholar] [PubMed] [Green Version]
- He, X.; Tan, X.; Wang, X.; Jin, H.; Liu, L.; Ma, L.; Yu, H.; Fan, Z. C-Myc-activated long noncoding RNA CCAT1 promotes colon cancer cell proliferation and invasion. Tumor Biol. 2014, 35, 12181–12188. [Google Scholar] [CrossRef]
- Treeck, O.; Skrzypczak, M.; Schüler-Toprak, S.; Weber, F.; Ortmann, O. Long non-coding RNA CCAT1 is overexpressed in endometrial cancer and regulates growth and transcriptome of endometrial adenocarcinoma cells. Int. J. Biochem. Cell Biol. 2020, 122, 105740. [Google Scholar] [CrossRef]
- Liu, J.; Ding, D.; Jiang, Z.; Du, T.; Liu, J.; Kong, Z. Long non-coding RNA CCAT1/miR-148a/PKCzeta prevents cell migration of prostate cancer by altering macrophage polarization. Prostate 2019, 79, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Shi, H.; Ren, F.; Jia, Y.; Zhang, R. Long non-coding RNA CCAT1 promotes metastasis and poor prognosis in epithelial ovarian cancer. Exp. Cell Res. 2017, 359, 185–194. [Google Scholar] [CrossRef]
- Li, Y.; Zhu, G.; Ma, Y.; Qu, H. lncRNA CCAT1 contributes to the growth and invasion of gastric cancer via targeting miR-219-1. J. Cell. Biochem. 2019, 120, 19457–19468. [Google Scholar] [CrossRef]
- Dou, C.; Sun, L.; Jin, X.; Han, M.; Zhang, B.; Jiang, X.; Lv, J.; Li, T. Long non-coding RNA CARLo-5 promotes tumor progression in hepatocellular carcinoma via suppressing miR-200b expression. Oncotarget 2017, 8, 70172–70182. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.; Chen, Y.; Lin, Y.; Ye, L. Down-regulation of LncRNA CCAT1 enhances radiosensitivity via regulating miR-148b in breast cancer. Cell Biol. Int. 2018, 42, 227–236. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, Y. CCAT-1 promotes proliferation and inhibits apoptosis of cervical cancer cells via the Wnt signaling pathway. Oncotarget 2017, 8, 68059–68070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, L.; Jia, J.-Q.; Chen, J. The lncRNA CCAT1 Upregulates Proliferation and Invasion in Melanoma Cells via Suppressing miR-33a. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, K.; Wu, Q.; Jiang, S.; Yuan, H.; Huang, S.; Li, H. CCAT1 promotes laryngeal squamous cell carcinoma cell proliferation and invasion. Am. J. Transl. Res. 2016, 8, 4338–4345. [Google Scholar] [PubMed]
- Cui, B.; Li, B.; Liu, Q.; Cui, Y. lncRNA CCAT1 Promotes Glioma Tumorigenesis by Sponging miR-181b. J. Cell. Biochem. 2017, 118, 4548–4557. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, G.; Murugan, A.K.; Rao, H.P.S.; Subbiah, S.; Rajaraman, R.; Munirajan, A.K. Long non-coding RNA CCAT1 is overexpressed in oral squamous cell carcinomas and predicts poor prognosis. Biomed. Rep. 2017, 6, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhong, J.; Bian, Z.; Fang, X.; Peng, Y.; Hu, Y. Long non-coding RNA CCAT1 promotes human retinoblastoma SO-RB50 and Y79 cells through negative regulation of miR-218-5p. Biomed. Pharmacother. 2017, 87, 683–691. [Google Scholar] [CrossRef]
- Ma, M.-Z.; Chu, B.-F.; Zhang, Y.; Weng, M.-Z.; Qin, Y.-Y.; Gong, W.; Quan, Z.-W. Long non-coding RNA CCAT1 promotes gallbladder cancer development via negative modulation of miRNA-218-5p. Cell Death Dis. 2015, 6, e1583. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Zhang, Q.; Tian, X.H.; Wang, J.L.; Niu, Y.X.; Li, G. lncRNA CCAT1 is a biomarker for the proliferation and drug resistance of esophageal cancer via the miR-143/PLK1/BUBR1 axis. Mol. Carcinog. 2019, 58, 2207–2217. [Google Scholar] [CrossRef]
- Izadifard, M.; Pashaiefar, H.; Yaghmaie, M.; Montazeri, M.; Sadraie, M.; Momeny, M.; Jalili, M.; Ahmadvand, M.; Ghaffari, S.H.; Mohammadi, S.; et al. Expression Analysis of PVT1, CCDC26, and CCAT1 Long Noncoding RNAs in Acute Myeloid Leukemia Patients. Genet. Test. Mol. Biomark. 2018, 22, 593–598. [Google Scholar] [CrossRef]
- Hu, B.; Zhang, H.; Wang, Z.; Zhang, F.; Wei, H.; Li, L. LncRNA CCAT1/miR-130a-3p axis increases cisplatin resistance in non-small-cell lung cancer cell line by targeting SOX4. Cancer Biol. Ther. 2017, 18, 974–983. [Google Scholar] [CrossRef]
- Zhao, J.; Cheng, L. Long non-coding RNA CCAT1/miR-148a axis promotes osteosarcoma proliferation and migration through regulating PIK3IP1. Acta Biochim. Biophys. Sin. 2017, 49, 503–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zhang, W.; Hao, S. LncRNA CCAT1 modulates the sensitivity of paclitaxel in nasopharynx cancers cells via miR-181a/CPEB2 axis. Cell Cycle 2017, 16, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Hua, L.; Wei, L.; Meng, J.; Hu, J. Correlation Between CASC8, SMAD7 Polymorphisms and the Susceptibility to Colo-rectal Cancer: An Updated Meta-Analysis Based on GWAS Results. Medicine 2015, 94, e1884. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Zhong, P.; Xiong, L.; Duan, L. Long Noncoding RNA Cancer Susceptibility Candidate 8 Suppresses the Proliferation of Bladder Cancer Cells via Regulating Glycolysis. DNA Cell Biol. 2017, 36, 767–774. [Google Scholar] [CrossRef]
- Zhou, H.-H.; Chen, S.-H.; Lv, Q.-L.; Sun, B.; Qu, Q.; Qin, C.-Z.; Fan, L.; Guo, Y.; Cheng, L.; Zhou, H.-H. Clinical Significance of Long Non-Coding RNA CASC8 rs10505477 Polymorphism in Lung Cancer Susceptibility, Platinum-Based Chemotherapy Response, and Toxicity. Int. J. Environ. Res. Public Health 2016, 13, 545. [Google Scholar]
- Wu, E.R.; Hsieh, M.J.; Chiang, W.L.; Hsueh, K.C.; Yang, S.F.; Su, S.C. Association of lncRNA CCAT2 and CASC8 Gene Polymor-phisms with Hepatocellular Carcinoma. Int. J. Environ. Res. Public Health 2019, 16, 2833. [Google Scholar] [CrossRef] [Green Version]
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafà, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Li, X.; Shen, P.; Zhang, D. CCAT2 is an oncogenic long non-coding RNA in pancreatic ductal adenocarcinoma. Biol. Res. 2018, 51, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Wu, X.; Yin, X.; Du, F.; Liu, Y.; Ding, X. LncRNA CCAT2 promoted osteosarcoma cell proliferation and invasion. J. Cell. Mol. Med. 2018, 22, 2592–2599. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Wang, L.; He, X.; Zhang, J.; Zhu, Z.; Zhang, M.; Li, X. lncRNA CCAT2 promotes radiotherapy resistance for human esophageal carcinoma cells via the miR-145/p70S6K1 and p53 pathway. Int. J. Oncol. 2019, 56, 327–336. [Google Scholar] [CrossRef]
- Li, J.-J.; Zhu, J.-F.; Zhang, F.; Zhang, P.-P.; Zhang, J.-J. Upregulation of lncRNA CCAT2 predicts poor prognosis in patients with acute myeloid leukemia and is correlated with leukemic cell proliferation. Int. J. Clin. Exp. Pathol. 2018, 11, 5658–5666. [Google Scholar] [PubMed]
- Zhao, C.; Qiao, C.; Zong, L.; Chen, Y. Long non-coding RNA-CCAT2 promotes the occurrence of non-small cell lung cancer by regulating the Wnt/beta-catenin signaling pathway. Oncol. Lett. 2018, 16, 4600–4606. [Google Scholar] [PubMed]
- Qiu, M.; Xu, Y.; Yang, X.; Wang, J.; Hu, J.; Xu, L.; Yin, R. CCAT2 is a lung adenocarcinoma-specific long non-coding RNA and promotes invasion of non-small cell lung cancer. Tumor Biol. 2014, 35, 5375–5380. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.-D.; Jiang, J.; Liu, M.-M.; Zhuang, R.-J.; Wang, H.; Li, P.-L. Silencing CCAT2 inhibited proliferation and invasion of epithelial ovarian carcinoma cells by regulating Wnt signaling pathway. Int. J. Clin. Exp. Pathol. 2017, 10, 11771–11778. [Google Scholar]
- Xie, P.; Cao, H.; Li, Y.; Wang, J.; Cui, Z. Knockdown of lncRNA CCAT2 inhibits endometrial cancer cells growth and metastasis via sponging miR-216b. Cancer Biomark. 2017, 21, 123–133. [Google Scholar] [CrossRef]
- Wu, Z.J.; Li, Y.; Wu, Y.Z.; Wang, Y.; Nian, W.Q.; Wang, L.L.; Li, L.C.; Luo, H.L.; Wang, D.L. Long non-coding RNA CCAT2 promotes the breast cancer growth and metastasis by regulating TGF-beta signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 706–714. [Google Scholar]
- Huang, J.L.; Liao, Y.; Qiu, M.X.; Li, J.; An, Y. Long non-coding RNA CCAT2 promotes cell proliferation and invasion through regulating Wnt/beta-catenin signaling pathway in clear cell renal cell carcinoma. Tumour Biol. 2017, 39, 1010428317711314. [Google Scholar] [CrossRef] [Green Version]
- Hua, F.; Li, C.-H.; Chen, X.; Liu, X.-P. Long Noncoding RNA CCAT2 Knockdown Suppresses Tumorous Progression by Sponging miR-424 in Epithelial Ovarian Cancer. Oncol. Res. 2018, 26, 241–247. [Google Scholar] [CrossRef]
- Sun, S.-L.; Shu, Y.-G.; Tao, M.-Y. LncRNA CCAT2 promotes angiogenesis in glioma through activation of VEGFA signalling by sponging miR-424. Mol. Cell. Biochem. 2020, 468, 69–82. [Google Scholar] [CrossRef]
- You, Z.; Liu, C.; Wang, C.; Ling, Z.; Wang, Y.; Wang, Y.; Zhang, M.; Chen, S.; Xu, B.; Guan, H.; et al. LncRNA CCAT1 Promotes Prostate Cancer Cell Proliferation by Interacting with DDX5 and MIR-28-5P. Mol. Cancer Ther. 2019, 18, 2469–2479. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhou, C.; Chang, Y.; Zhang, Z.; Hu, Y.; Zhang, F.; Lu, Y.; Zheng, L.; Zhang, W.; Li, X. Long non-coding RNA CASC11 interacts with hnRNP-K and activates the WNT/beta-catenin pathway to promote growth and metastasis in colorectal cancer. Cancer Lett. 2016, 376, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.; Liu, L.; Chen, X.; Zhang, Y.; Zhu, W. LncRNA CASC11 promotes the cervical cancer progression by activating Wnt/beta-catenin signaling pathway. Biol. Res. 2019, 52, 33. [Google Scholar] [CrossRef]
- Zhang, L.; Kang, W.; Lu, X.; Ma, S.; Dong, L.; Zou, B. LncRNA CASC11 promoted gastric cancer cell proliferation, migration and invasion in vitro by regulating cell cycle pathway. Cell Cycle 2018, 17, 1886–1900. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Xu, C.; Le, W.; Ge, B.; Wang, T. lncRNA CASC11 promotes cancer cell proliferation in bladder cancer through miR-NA-150. J. Cell Biochem. 2019, 120, 13487–13493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Zhang, P.; Nan, H.; Lu, Y.; Zhao, J.; Yang, M.; Song, Q. LncRNA CASC11 promotes TGF-β1, increases cancer cell stemness and predicts postoperative survival in small cell lung cancer. Gene 2019, 704, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Jiang, Y.; Lai, B.; Lin, Y.; Wen, J. The positive feedback loop FOXO3/CASC11/miR-498 promotes the tumorigenesis of non-small cell lung cancer. Biochem. Biophys. Res. Commun. 2019, 519, 518–524. [Google Scholar] [CrossRef]
- Cheng, N.; Wu, J.; Yin, M.; Xu, J.; Wang, Y.; Chen, X.; Nie, Z.; Yin, J. LncRNA CASC11 promotes cancer cell proliferation in hepatocellular carcinoma by inhibiting miRNA-188-5p. Biosci. Rep. 2019, 39, BSR20190251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.-G.; Wang, C.-H.; He, R.-Q.; Xu, R.-Y.; Ji, C.-B. LncRNA CASC11 promotes the development of esophageal carcinoma by regulating KLF6. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8878–8887. [Google Scholar]
- Yu, Z.; Zhang, J.; Han, J. Silencing CASC11 curbs neonatal neuroblastoma progression through modulating mi-croRNA-676-3p/nucleolar protein 4 like (NOL4L) axis. Pediatric Res. 2020, 87, 662–668. [Google Scholar] [CrossRef]
- Shen, F.; Feng, L.; Zhou, J.; Zhang, H.; Xu, Y.; Jiang, R.; Zhang, H.; Chen, Y. Overexpression of CASC11 in ovarian squamous cell carcinoma mediates the development of cancer cell resistance to chemotherapy. Gene 2019, 710, 363–366. [Google Scholar] [CrossRef]
- Song, K.; Yuan, X.; Li, G.; Ma, M.; Sun, J. Long noncoding RNA CASC11 promotes osteosarcoma metastasis by suppressing degradation of snail mRNA. Am. J. Cancer Res. 2019, 9, 300–311. [Google Scholar] [PubMed]
- Jin, J.; Zhang, S.; Hu, Y.; Zhang, Y.; Guo, C.; Feng, F. SP1 induced lncRNA CASC11 accelerates the glioma tumorigenesis through targeting FOXK1 via sponging miR-498. Biomed. Pharmacother. 2019, 116, 108968. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.; Xu, B.; He, L.; Zhang, G. PVT1 Promotes Angiogenesis by Regulating miR-29c/Vascular Endothelial Growth Factor (VEGF) Signaling Pathway in Non-Small-Cell Lung Cancer (NSCLC). Med. Sci. Monit. 2019, 25, 5418–5425. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhao, J.; He, Y. Long non-coding RNA PVT1 functions as an oncogene in human colon cancer through miR-30d-5p/RUNX2 axis. J. B.U.ON. Off. J. Balk. Union Oncol. 2018, 23, 48–54. [Google Scholar]
- Ding, Y.; Fang, Q.; Li, Y.; Wang, Y. Amplification of lncRNA PVT1 promotes ovarian cancer proliferation by binding to miR-140. Mamm. Genome 2019, 30, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, Y.; Jin, J.; Han, G.; Sun, C.; Pizzi, M.P.; Huo, L.; Scott, A.; Wang, Y.; Ma, L.; et al. LncRNA PVT1 up-regulation is a poor prognosticator and serves as a therapeutic target in esophageal adenocarcinoma. Mol. Cancer 2019, 18, 141. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Jing, Y.; Wei, F.; Tang, Y.; Yang, L.; Luo, J.; Yang, P.; Ni, Q.; Pang, J.; Liao, Q.; et al. Long non-coding RNA PVT1 predicts poor prognosis and induces radioresistance by regulating DNA repair and cell apoptosis in nasopharyngeal carcinoma. Cell Death Dis. 2018, 9, 235. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zou, H.; Yang, H.; Wang, L.; Chu, H.; Jiao, J.; Wang, Y.; Chen, A. Long noncoding RNA plasmacytoma variant trans-location 1 gene promotes the development of cervical cancer via the NFkappaB pathway. Mol. Med. Rep. 2019, 20, 2433–2440. [Google Scholar]
- Fang, J.; Huang, J. Clinical significance of the expression of long non-coding RNA PVT1 in glioma. Cancer Biomark. 2019, 24, 509–513. [Google Scholar] [CrossRef]
- Tian, Z.; Cao, S.; Li, C.; Xu, M.; Wei, H.; Yang, H.; Sun, Q.; Ren, Q.; Zhang, L. LncRNA PVT1 regulates growth, migration, and invasion of bladder cancer by miR-31/ CDK1. J. Cell. Physiol. 2019, 234, 4799–4811. [Google Scholar] [CrossRef]
- Zhang, X.; Feng, W.; Zhang, J.; Ge, L.; Zhang, Y.; Jiang, X.; Peng, W.; Wang, D.; Gong, A.; Xu, M. Long noncoding RNA PVT1 promotes epithelialmesenchymal transition via the TGFbeta/Smad pathway in pancreatic cancer cells. Oncol Rep. 2018, 40, 1093–1102. [Google Scholar] [PubMed]
- Yang, L.; Peng, X.; Jin, H.; Liu, J. Long non-coding RNA PVT1 promotes autophagy as ceRNA to target ATG3 by sponging microRNA-365 in hepatocellular carcinoma. Gene 2019, 697, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Liu, G.; Zhang, X.; Du, N. Long noncoding RNA TMEM75 promotes colorectal cancer progression by activation of SIM2. Gene 2018, 675, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, P.; Mo, W.; Zhang, Y.; Zhou, W.; Deng, T.; Zhou, M.; Chen, X.; Wang, S.; Wang, C. lncRNA-CCDC26, as a novel biomarker, predicts prognosis in acute myeloid leukemia. Oncol Lett. 2019, 18, 2203–2211. [Google Scholar] [CrossRef]
- Hirano, T.; Yoshikawa, R.; Harada, H.; Harada, Y.; Ishida, A.; Yamazaki, T. Long noncoding RNA, CCDC26, controls myeloid leukemia cell growth through regulation of KIT expression. Mol. Cancer 2015, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hui, Y.; Li, X.; Jia, Q. Silencing of lncRNA CCDC26 Restrains the Growth and Migration of Glioma Cells In Vitro and In Vivo via Targeting miR-203. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2018, 26, 1143–1154. [Google Scholar] [CrossRef]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Zhou, K.; Liu, M.; Cao, Y. New Insight into microRNA Functions in Cancer: Oncogene–microRNA–Tumor Suppressor Gene Network. Front. Mol. Biosci. 2017, 4, 46. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Hu, Y. Long Non-coding RNA PVT1 Competitively Binds MicroRNA-424-5p to Regulate CARM1 in Radiosensi-tivity of Non-Small-Cell Lung Cancer. Mol. Ther. Nucleic Acids 2019, 16, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Hu, N.; Wang, C.; Zhao, H.; Gu, Y. Long non-coding RNA CCAT1 promotes multiple myeloma progression by acting as a molecular sponge of miR-181a-5p to modulate HOXA1 expression. Cell Cycle 2018, 17, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Yue, P.; Wang, Y.; Chen, G.; Li, Y. PCAT-1 contributes to cisplatin resistance in gastric cancer through miR-128/ZEB1 axis. Biomed. Pharmacother. 2019, 118, 109255. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kong, D.; Sun, D.; Li, J. Long non-coding RNA CCAT2 acts as an oncogene in osteosarcoma through regulation of miR-200b/VEGF. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2994–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, W.; Han, T.C.; Wang, W.; Zhao, J. LncRNA CASC11 promotes the development of lung cancer through targeting mi-croRNA-302/CDK1 axis. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6539–6547. [Google Scholar] [PubMed]
- Shen, S.; Li, K.; Liu, Y.; Yang, C.-L.; He, C.; Wang, H. Down-regulation of long noncoding RNA PVT1 inhibits esophageal carcinoma cell migration and invasion and promotes cell apoptosis via microRNA-145-mediated inhibition of FSCN1. Mol. Oncol. 2019, 13, 2554–2573. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yu, Y.; Li, H.; Hu, Q.; Chen, X.; He, Y.; Xue, C.; Ren, F.; Ren, Z.; Li, J.; et al. Long non-coding RNA PVT1 promotes tumor progression by regulating the miR-143/HK2 axis in gallbladder cancer. Mol. Cancer 2019, 18, 33. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Li, D.; Zhang, X.; Liu, N.; Chi, G.; Jin, X. LncRNA PVT1 Facilitates Tumorigenesis and Progression of Glioma via Regulation of MiR-128-3p/GREM1 Axis and BMP Signaling Pathway. Neurotherapeutics 2018, 15, 1139–1157. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Cui, J.; Song, Y. Long noncoding RNA PVT1 promotes EMT via mediating microRNA-186 targeting of Twist1 in prostate cancer. Gene 2018, 654, 36–42. [Google Scholar] [CrossRef]
- Chen, L.; Han, X.; Hu, Z.; Chen, L. The PVT1/miR-216b/Beclin-1 regulates cisplatin sensitivity of NSCLC cells via modulating autophagy and apoptosis. Cancer Chemother. Pharmacol. 2019, 83, 921–931. [Google Scholar] [CrossRef]
- Feng, K.; Liu, Y.; Xu, L.-J.; Zhao, L.-F.; Jia, C.-W.; Xu, M.-Y. Long noncoding RNA PVT1 enhances the viability and invasion of papillary thyroid carcinoma cells by functioning as ceRNA of microRNA-30a through mediating expression of insulin like growth factor 1 receptor. Biomed. Pharmacother. 2018, 104, 686–698. [Google Scholar] [CrossRef]
- Huang, F.; Chen, W.; Peng, J.; Li, Y.; Zhuang, Y.; Zhu, Z.; Shao, C.; Yang, W.; Yao, H.; Zhang, S. LncRNA PVT1 triggers Cy-to-protective autophagy and promotes pancreatic ductal adenocarcinoma development via the miR-20a-5p/ULK1 Axis. Mol. Cancer 2018, 17, 98. [Google Scholar] [CrossRef]
- Zheng, J.; Hu, L.; Cheng, J.; Xu, J.; Zhong, Z.; Yang, Y.; Yuan, Z. lncRNA PVT1 promotes the angiogenesis of vascular endothelial cell by targeting miR-26b to activate CTGF/ANGPT2. Int. J. Mol. Med. 2018, 42, 489–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Q.-Q.; Chen, C.-Y.; Chen, Z.; Chang, S. LncRNA PVT1 promotes proliferation and invasion through enhancing Smad3 expression by sponging miR-140-5p in cervical cancer. Radiol. Oncol. 2019, 53, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Luo, X.; He, W.; Chen, G.; Li, Y.; Li, W.; Wang, X.; Lai, Y.; Ye, Y. Long Non-Coding RNA PVT1/miR-150/ HIG2 Axis Regulates the Proliferation, Invasion and the Balance of Iron Metabolism of Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2018, 49, 1403–1419. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Liu, S.; Xing, G.; Wang, F. lncRNA PVT1/MicroRNA-17-5p/PTEN Axis Regulates Secretion of E2 and P4, Proliferation, and Apoptosis of Ovarian Granulosa Cells in PCOS. Mol. Ther. Nucleic Acids 2020, 20, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Longfei, L.; Long, W.; Feng, Z.; Chen, J.; Chao, L.; Peihua, L.; Xiongbing, Z.; Chen, H. LncRNA PVT1 regulates VEGFC through inhibiting miR-128 in bladder cancer cells. J. Cell. Physiol. 2019, 234, 1346–1353. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Chen, Y.; Kong, W.; Fu, L.; Liu, Y.; Yao, Q.; Yuan, Y. PVT1-derived miR-1207-5p promotes breast cancer cell growth by targeting STAT6. Cancer Sci. 2017, 108, 868–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hu, H. Long non-coding RNA CCAT1/miR-218/ZFX axis modulates the progression of laryngeal squamous cell cancer. Tumour Biol. 2017, 39, 1010428317699417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, C.; Sun, L.; Jin, X.; Han, M.; Zhang, B.; Li, T. Long non-coding RNA colon cancer-associated transcript 1 functions as a competing endogenous RNA to regulate cyclin-dependent kinase 1 expression by sponging miR-490-3p in hepatocellular car-cinoma progression. Tumour Biol. 2017, 39, 1010428317697572. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Li, X.; Fan, Q.; Liu, G.; Yin, J. CCAT1 promotes triple-negative breast cancer progression by suppressing miR-218/ZFX signaling. Aging 2019, 11, 4858–4875. [Google Scholar] [CrossRef]
- Xu, W.; Chang, J.; Du, X.; Hou, J. Long non-coding RNA PCAT-1 contributes to tumorigenesis by regulating FSCN1 via miR-145-5p in prostate cancer. Biomed. Pharmacother. 2017, 95, 1112–1118. [Google Scholar] [CrossRef]
- Zhang, D.; Cao, J.; Zhong, Q.; Zeng, L.; Cai, C.; Lei, L.; Zhang, W.; Liu, F. Long noncoding RNA PCAT-1 promotes invasion and metastasis via the miR-129-5p-HMGB1 signaling pathway in hepatocellular carcinoma. Biomed. Pharmacother. 2017, 95, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Wang, C.-X.; Wang, H.; Wang, Y.; Hu, S.; Cao, Z.; Min, B.; Li, L.; Tian, X.; Hu, H.-B. Long noncoding RNA CCAT2 functions as a competitive endogenous RNA to regulate FOXC1 expression by sponging miR-23b-5p in lung adenocarcinoma. J. Cell. Biochem. 2019, 120, 7998–8007. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Jiang, A. Long noncoding RNA CCDC26 as a potential predictor biomarker contributes to tumorigenesis in pan-creatic cancer. Biomed Pharmacother. 2016, 83, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Wijesinghe, S.; Halsall, J.; Kanhere, A. A long intergenic non-coding RNA regulates nuclear localisation of DNA methyl transferase-1. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.-T.; Fang, L.; Cheng, Y.-X.; Sun, Q. LncRNA PVT1 regulates prostate cancer cell growth by inducing the methylation of miR-146a. Cancer Med. 2016, 5, 3512–3519. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, G.; Liu, J. Long noncoding RNA PVT1 promotes cervical cancer progression through epigenetically silencing miR-200b. APMIS 2016, 124, 649–658. [Google Scholar] [CrossRef]
- Zhang, C.; Shao, S.; Zhang, Y.; Wang, L.; Liu, J.; Fang, F.; Li, P.; Wang, B. LncRNA PCAT1 promotes metastasis of endometrial carcinoma through epigenetical downregulation of E-cadherin associated with methyltransferase EZH2. Life Sci. 2020, 243, 117295. [Google Scholar] [CrossRef]
- Huang, J.; Deng, G.; Liu, T.; Chen, W.; Zhou, Y. Long noncoding RNA PCAT-1 acts as an oncogene in osteosarcoma by reducing p21 levels. Biochem. Biophys. Res. Commun. 2018, 495, 2622–2629. [Google Scholar] [CrossRef]
- Shah, M.Y.; Ferracin, M.; Pileczki, V.; Chen, B.; Redis, R.; Fabris, L.; Zhang, X.; Ivan, C.; Shimizu, M.; Rodriguez-Aguayo, C.; et al. Cancer-associated rs6983267 SNP and its accompanying long noncoding RNA CCAT2 induce myeloid malignancies via unique SNP-specific RNA mutations. Genome Res. 2018, 28, 432–447. [Google Scholar] [CrossRef] [Green Version]
- Shtivelman, E.; Bishop, J.M. The PVT gene frequently amplifies with MYC in tumor cells. Mol. Cell. Biol. 1989, 9, 1148–1154. [Google Scholar] [CrossRef]
- Jin, K.; Wang, S.; Zhang, Y.; Xia, M.; Mo, Y.; Li, X.; Li, G.; Zeng, Z.; Xiong, W.; He, Y. Long non-coding RNA PVT1 interacts with MYC and its downstream molecules to synergistically promote tumorigenesis. Cell. Mol. Life Sci. 2019, 76, 4275–4289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.Y.; Moriarity, B.S.; Gong, W.; Akiyama, R.; Tiwari, A.; Kawakami, H.; Ronning, P.; Reuland, B.; Guenther, K.; Beadnell, T.C.; et al. PVT1 dependence in cancer with MYC copy-number increase. Nature 2014, 512, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, P.; Morris, K.V. Expanding the functional role of long noncoding RNAs. Cell Res. 2014, 24, 1284–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, E.; Cunningham, M.; Arnold, H.; Chasse, D.; Monteith, T.; Ivaldi, G.; Hahn, W.C.; Stukenberg, P.T.; Shenolikar, S.; Uchida, T.; et al. A signalling pathway controlling c-Myc degradation that impacts oncogenic trans-formation of human cells. Nat. Cell Biol. 2004, 6, 308–318. [Google Scholar] [CrossRef]
- Cho, S.W.; Xu, J.; Sun, R.; Mumbach, M.R.; Carter, A.C.; Chen, Y.G.; Yost, K.E.; Kim, J.; He, J.; Nevins, S.A.; et al. Promoter of lncRNA Gene PVT1 Is a Tumor-Suppressor DNA Boundary Element. Cell 2018, 173, 1398–1412.e22. [Google Scholar] [CrossRef] [Green Version]
- Prensner, J.R.; Chen, W.; Han, S.; Iyer, M.K.; Cao, Q.; Kothari, V.; Evans, J.R.; Knudsen, K.E.; Paulsen, M.T.; Ljungman, M.; et al. The long non-coding RNA PCAT-1 promotes prostate cancer cell proliferation through cMyc. Neoplasia 2014, 16, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Kajino, T.; Shimamura, T.; Gong, S.; Yanagisawa, K.; Ida, L.; Nakatochi, M.; Griesing, S.; Shimada, Y.; Kano, K.; Suzuki, M.; et al. Divergent lncRNA MYMLR regulates MYC by eliciting DNA looping and promoter-enhancer interaction. EMBO J. 2019, 38, e98441. [Google Scholar] [CrossRef]
- Xiang, J.-F.; Yin, Q.-F.; Chen, T.; Zhang, Y.; Zhang, X.-O.; Wu, Z.; Zhang, S.; Wang, H.-B.; Ge, J.; Lu, X.; et al. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014, 24, 513–531. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, N.; Sugai, T.; Habano, W.; Eizuka, M.; Sugimoto, R.; Akasaka, R.; Toya, Y.; Yamamoto, E.; Koeda, K.; Sasaki, A.; et al. Genome-wide analysis of DNA copy number alterations in early and advanced gastric cancers. Mol. Carcinog. 2016, 56, 527–537. [Google Scholar] [CrossRef]
- Naidu, R.; Ching, H.C.; Seong, M.K.; Har, Y.C.; Taib, N.A.M. Integrated analysis of copy number and loss of heterozygosity in primary breast carcinomas using high-density SNP array. Int. J. Oncol. 2011, 39, 621–633. [Google Scholar] [CrossRef] [Green Version]
- Alberto, L.; Tolomeo, D.; Cifola, I.; Severgnini, M.; Turchiano, A.; Augello, B.; Squeo, G.; Pietro, D.; Traversa, D.; Daniele, G.; et al. MYC-containing amplicons in acute myeloid leukemia: Genomic structures, evolution, and transcriptional consequences. Leukemia 2018, 32, 2152–2166. [Google Scholar]
- Haverty, P.M.; Hon, L.S.; Kaminker, J.S.; Chant, J.; Zhang, Z. High-resolution analysis of copy number alterations and associated expression changes in ovarian tumors. BMC Med Genom. 2009, 2, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radtke, I.; Mullighan, C.G.; Ishii, M.; Su, X.; Cheng, J.; Ma, J.; Ganti, R.; Cai, Z.; Goorha, S.; Pounds, S.B.; et al. Genomic analysis reveals few genetic alterations in pediatric acute myeloid leukemia. Proc. Natl. Acad. Sci. USA 2009, 106, 12944–12949. [Google Scholar] [CrossRef] [Green Version]
- Quigley, D.A.; Dang, H.X.; Zhao, S.G.; Lloyd, P.; Aggarwal, R.; Alumkal, J.J.; Foye, A.; Kothari, V.; Perry, M.D.; Bailey, A.M.; et al. Genomic Hallmarks and Structural Variation in Meta-static Prostate Cancer. Cell 2018, 174, 758–769.e9. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Rondon, N.; Villegas, V.E.; Rondon-Lagos, M. The Role of Chromosomal Instability in Cancer and Therapeutic Re-sponses. Cancers 2017, 10, 4. [Google Scholar] [CrossRef] [Green Version]
- Cory, S.; Graham, M.; Webb, E.; Corcoran, L.; Adams, J.M. Variant (6;15) translocations in murine plasmacytomas involve a chromosome 15 locus at least 72 kb from the c-myc oncogene. EMBO J. 1985, 4, 675–681. [Google Scholar] [CrossRef]
- Graham, M.; Adams, J.M. Chromosome 8 breakpoint far 3’ of the c-myc oncogene in a Burkitt’s lymphoma 2;8 variant translocation is equivalent to the murine pvt-1 locus. EMBO J. 1986, 5, 2845–2851. [Google Scholar] [CrossRef] [PubMed]
- Chinen, Y.; Sakamoto-Inada, N.; Nagoshi, H.; Taki, T.; Maegawa, S.; Tatekawa, S.; Tsukamoto, T.; Mizutani, S.; Shimura, Y.; Yamamoto-Sugitani, M.; et al. 8q24 amplified segments involve novel fusion genes between NSMCE2 and long noncoding RNAs in acute myelogenous leukemia. J. Hematol. Oncol. 2014, 7, 68. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, H.; Taki, T.; Hanamura, I.; Nitta, M.; Otsuki, T.; Nishida, K.; Okuda, K.; Sakamoto, N.; Kobayashi, S.; Yamamoto-Sugitani, M.; et al. Frequent PVT1 rearrangement and novel chi-meric genes PVT1-NBEA and PVT1-WWOX occur in multiple myeloma with 8q24 abnormality. Cancer Res. 2012, 72, 4954–4962. [Google Scholar] [CrossRef] [Green Version]
- Taniwaki, M. Recent advancements in molecular cytogenetics for hematological malignancies: Identification of novel PVT1 fusion genes. Rinsho Ketsueki 2015, 56, 2056–2065. [Google Scholar]
- Northcott, P.A.; Shih, D.J.H.; Peacock, J.; Garzia, L.; Morrissy, A.S.; Zichner, T.; Stuetz, A.M.; Korshunov, A.; Reimand, J.; Schumacher, S.E.; et al. Subgroup-specific structural variation across 1000 medulloblastoma genomes. Nat. Cell Biol. 2012, 488, 49–56. [Google Scholar]
- Sarver, A.L.; Murray, C.D.; Temiz, N.A.; Tseng, Y.Y.; Bagchi, A. MYC and PVT1 synergize to regulate RSPO1 levels in breast cancer. Cell Cycle 2016, 15, 881–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Ding, L.; Chang, T.-C.; Shao, Y.; Chiang, J.; Mulder, H.; Wang, S.; Shaw, T.I.; Wen, J.; Hover, L.; et al. Structure and evolution of double minutes in diagnosis and relapse brain tumors. Acta Neuropathol. 2019, 137, 123–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storlazzi, C.T.; Fioretos, T.; Surace, C.; Lonoce, A.; Mastrorilli, A.; Strömbeck, B.; D’Addabbo, P.; Iacovelli, F.; Minervini, C.; Aventin, A.; et al. MYC-containing double minutes in hematologic malignancies: Evidence in favor of the episome model and exclusion of MYC as the target gene. Hum. Mol. Genet. 2006, 15, 933–942. [Google Scholar] [CrossRef]
- Hirano, T.; Ike, F.; Murata, T.; Obata, Y.; Utiyama, H.; Yokoyama, K.K. Genes encoded within 8q24 on the amplicon of a large extrachromosomal element are selectively repressed during the terminal differentiation of HL-60 cells. Mutat. Res. Mol. Mech. Mutagen. 2008, 640, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Redis, R.S.; Vela, L.E.; Lu, W.; de Oliveira, J.F.; Ivan, C.; Rodriguez-Aguayo, C.; Adamoski, D.; Pasculli, B.; Taguchi, A.; Chen, Y.; et al. Allele-Specific Reprogramming of Cancer Metabolism by the Long Non-coding RNA CCAT2. Mol. Cell. 2016, 61, 520–534. [Google Scholar] [CrossRef] [Green Version]
- Jendrzejewski, J.; He, H.; Radomska, H.S.; Li, W.; Tomsic, J.; Liyanarachchi, S.; Davuluri, R.V.; Nagy, R.; De La Chapelle, A. The polymorphism rs944289 predisposes to papillary thyroid carcinoma through a large intergenic noncoding RNA gene of tumor suppressor type. Proc. Natl. Acad. Sci. USA 2012, 109, 8646–8651. [Google Scholar] [CrossRef] [Green Version]
- Hua, J.T.; Ahmed, M.; Guo, H.; Zhang, Y.; Chen, S.; Soares, F.; Lu, J.; Zhou, S.; Wang, M.; Li, H.; et al. Risk SNP-Mediated Promoter-Enhancer Switching Drives Prostate Cancer through lncRNA PCAT19. Cell 2018, 174, 564–575.e18. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Liu, H.; Liu, Z.; Owzar, K.; Han, Y.; Su, L.; Wei, Y.; Hung, R.J.; McLaughlin, J.; Brhane, Y.; et al. A Novel Genetic Variant in Long Non-coding RNA Gene NEXN-AS1 is Associated with Risk of Lung Cancer. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef]
- Taheri, M.; Habibi, M.; Noroozi, R.; Rakhshan, A.; Sarrafzadeh, S.; Sayad, A.; Omrani, M.D.; Ghafouri-Fard, S. HOTAIR genetic variants are associated with prostate cancer and benign prostate hyperplasia in an Iranian population. Gene 2017, 613, 20–24. [Google Scholar] [CrossRef]
- Miao, Y.-R.; Liu, W.; Zhang, Q.; Guo, A.-Y. lncRNASNP2: An updated database of functional SNPs and mutations in human and mouse lncRNAs. Nucleic Acids Res. 2018, 46, D276–D280. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Sun, H.; Wang, Y.; Yang, X.; Meng, Q.; Yang, H.; Zhu, H.; Tang, W.; Li, X.; Aschner, M.; et al. MALAT1 rs664589 Poly-morphism Inhibits Binding to miR-194-5p, Contributing to Colorectal Cancer Risk, Growth, and Metastasis. Cancer Res. 2019, 79, 5432–5441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, C.; Yan, T.; Wang, Z.; Su, H.-C.; Zhu, X.; Tian, X.; Fang, J.-Y.; Chen, H.; Hong, J. Variant of SNP rs1317082 at CCSlnc362 (RP11-362K14.5) creates a binding site for miR-4658 and diminishes the susceptibility to CRC. Cell Death Dis. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onay, U.V.; Briollais, L.; Knight, J.A.; Shi, E.; Wang, Y.; Wells, S.; Li, H.; Rajendram, I.; Andrulis, I.L.; Ozcelik, H. SNP-SNP interactions in breast cancer susceptibility. BMC Cancer 2006, 6, 114. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-Y.; Callan, C.Y.; Fang, Z.; Tung, H.-Y.; Park, J.Y. Interactions of PVT1 and CASC11 on Prostate Cancer Risk in African Americans. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1067–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tseng, Y.Y.; Bagchi, A. The PVT1-MYC duet in cancer. Mol. Cell. Oncol. 2015, 2, e974467. [Google Scholar] [CrossRef] [Green Version]
- Homer-Bouthiette, C.; Zhao, Y.; Shunkwiler, L.; Van Peel, B.; Garrett-Mayer, E.; Baird, R.C.; Rissman, A.I.; Guest, S.T.; Ethier, S.P.; John, M.C.; et al. Deletion of the murine ortholog of the 8q24 gene desert has anti-cancer effects in transgenic mammary cancer models. BMC Cancer 2018, 18, 1233. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, L. New Insights into Long Non-Coding RNA MALAT1 in Cancer and Metastasis. Cancers 2019, 11, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Mechanisms of lncRNAs. LncRNAs (in red) function in the nucleus and cytoplasm to act on transcriptional, post-transcriptional and translational levels. Within the nucleus, lncRNAs can regulate transcription through (A) recruiting chromatin modifying complexes to supress or activate transcriptional start sites (TSS) or (B) modulating transcription factor activity. LncRNAs can also (C) interact with mRNA to alter splicing events and (D) act as a scaffold and bind numerous proteins to form a ribonucleoprotein complex which can regulate transcription within the nucleus and (G) translation in the cytoplasm. In the cytoplasm, lncRNAs can also regulate gene expression post-transcriptionally by (E) acting as a microRNA (miRNA) sponge or interact with mRNA to (F) promote degradation or (G) regulate translation [19].
Figure 1.
Mechanisms of lncRNAs. LncRNAs (in red) function in the nucleus and cytoplasm to act on transcriptional, post-transcriptional and translational levels. Within the nucleus, lncRNAs can regulate transcription through (A) recruiting chromatin modifying complexes to supress or activate transcriptional start sites (TSS) or (B) modulating transcription factor activity. LncRNAs can also (C) interact with mRNA to alter splicing events and (D) act as a scaffold and bind numerous proteins to form a ribonucleoprotein complex which can regulate transcription within the nucleus and (G) translation in the cytoplasm. In the cytoplasm, lncRNAs can also regulate gene expression post-transcriptionally by (E) acting as a microRNA (miRNA) sponge or interact with mRNA to (F) promote degradation or (G) regulate translation [19].

Figure 2.
Map of the 8q24.21 region of the human genome. The 8q24.21 region (chr8:126,300,001–130,400,000) is predominately a non-coding region containing numerous lncRNAs (in red), with some microRNAs (in green) and few protein-coding genes (in blue). The direction of arrows indicates the direction in which the genes are transcribed.
Figure 2.
Map of the 8q24.21 region of the human genome. The 8q24.21 region (chr8:126,300,001–130,400,000) is predominately a non-coding region containing numerous lncRNAs (in red), with some microRNAs (in green) and few protein-coding genes (in blue). The direction of arrows indicates the direction in which the genes are transcribed.

Figure 3.
Potential effects of genetic variants on 8q24.21 lncRNA expression and function. (A) Copy number gains or single-nucleotide polymorphisms (SNPs) within the promoter region can alter the expression of lncRNAs (in red). (B) Chromosomal translocations can lead to the formation fusion genes containing 8q24.21 lncRNAs. (C) An intronic SNP can result in different lncRNA isoforms being expression including short and long versions of the lncRNA. SNPs located in intronic or exonic regions can result in changes in the structure, therefore (D) preventing binding to proteins, or (E) alterations in the miRNA binding domains resulting in either new binding sites or preventing binding to target miRNAs.
Figure 3.
Potential effects of genetic variants on 8q24.21 lncRNA expression and function. (A) Copy number gains or single-nucleotide polymorphisms (SNPs) within the promoter region can alter the expression of lncRNAs (in red). (B) Chromosomal translocations can lead to the formation fusion genes containing 8q24.21 lncRNAs. (C) An intronic SNP can result in different lncRNA isoforms being expression including short and long versions of the lncRNA. SNPs located in intronic or exonic regions can result in changes in the structure, therefore (D) preventing binding to proteins, or (E) alterations in the miRNA binding domains resulting in either new binding sites or preventing binding to target miRNAs.
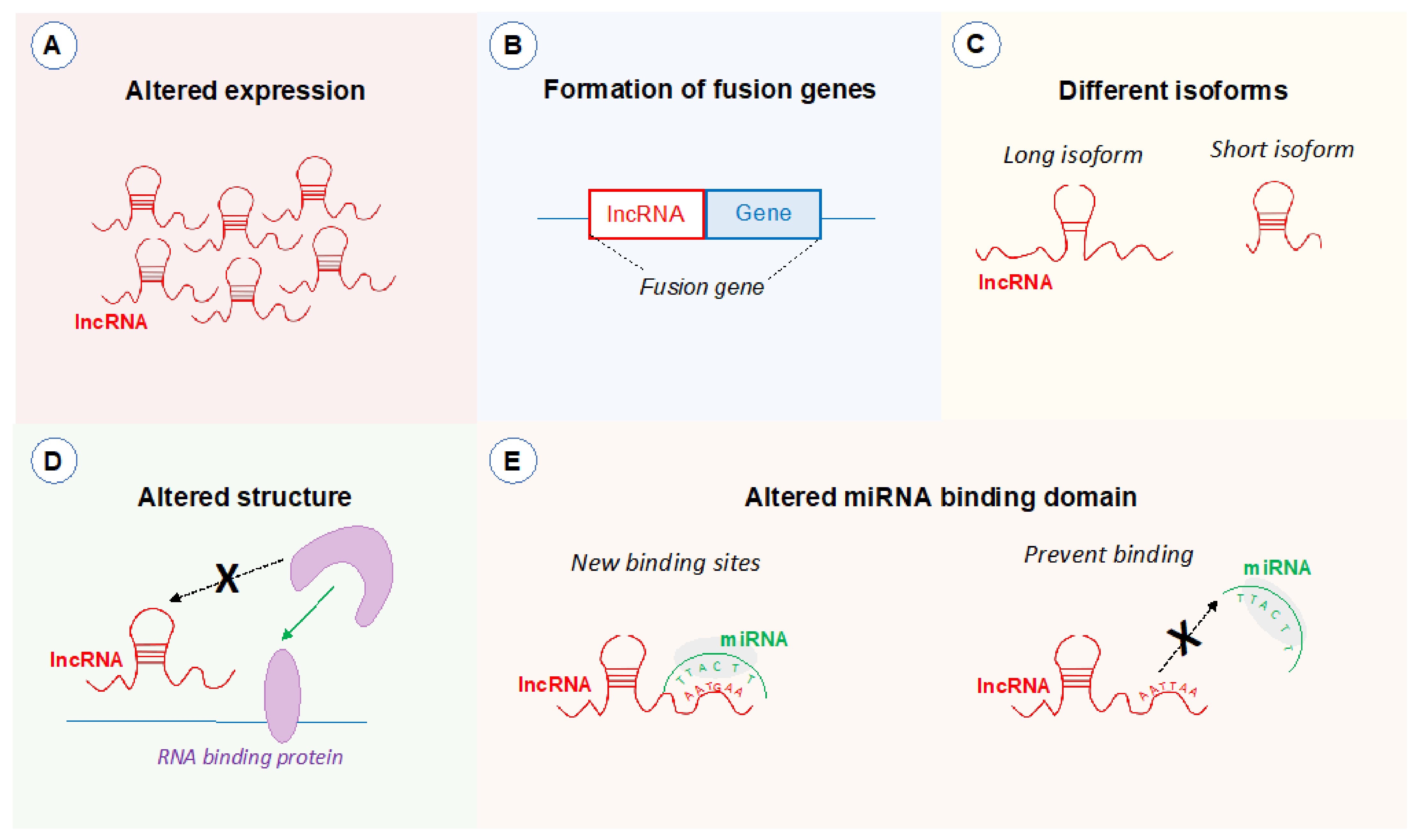

{kind=link}
{kind=link}
{kind=link}
Table 1.
Summary of cancers associated with lncRNAs located in the 8q24.21 region.
| 8q24.21 LncRNA | Cancers in Which the lncRNA Has Been Studied |
|---|---|
| PCAT1 | Colorectal cancer [40], hepatocellular carcinoma [41], extrahepatic cholangiocarcinoma [42], oesophageal squamous carcinoma [43], breast cancer [44], head and neck squamous carcinoma [45], laryngeal cancer [46], osteosarcoma [47], bladder cancer [48], glioma [49] |
| CASC19 | Colorectal cancer [50], cervical cancer [51] |
| PRNCR1 | Non-small cell lung cancer [52], tongue squamous cell carcinoma [53] |
| CCAT1 | Colon cancer [54], endometrial cancer [55], prostate cancer [56], ovarian cancer [57], gastric cancer [58], hepatocellular carcinoma [59], breast cancer [60], cervical cancer [61], melanoma [62], laryngeal squamous cell carcinoma [63], glioma [64], oral squamous cell carcinoma [65], retinoblastoma [66], gallbladder cancer [67], oesophageal cancer [68], acute myeloid leukaemia [69], non-small cell lung cancer [70], osteosarcoma [71], nasopharynx cancer [72] |
| CASC8 | Colorectal cancer [73], bladder cancer [74], lung cancer [75], hepatocellular carcinoma [76] |
| CCAT2 | Colon cancer [77], pancreatic ductal adenocarcinoma [78], osteosarcoma [79], oesophageal carcinoma [80], hepatocellular carcinoma [76], acute myeloid leukaemia [81], non-small cell lung cancer [82], lung adenocarcinoma [83], ovarian carcinoma [84], endometrial cancer [85], breast cancer [86], renal cell carcinoma [87], epithelial ovarian cancer [88], glioma [89], prostate cancer [90] |
| CASC11 | Colorectal cancer [91], cervical cancer [92], gastric cancer [93], bladder cancer [94], small cell lung cancer [95], non-small cell lung cancer [96], hepatocellular carcinoma [97], oesophageal carcinoma [98], neonatal neuroblastoma [99], ovarian squamous cell carcinoma [100], osteosarcoma [101], glioma [102] |
| PVT1 | Non-small cell lung cancer [103], colon cancer [104], ovarian cancer [105], oesophageal adenocarcinoma [106], nasopharyngeal carcinoma [107], cervical cancer [108], glioma [109], bladder cancer [110], pancreatic cancer [111], hepatocellular carcinoma [112] |
| TMEM75 | Colorectal cancer [113] |
| CCDC26 | Acute myeloid leukaemia [114], acute monocytic leukaemia [115], chronic myeloid leukaemia [115], glioma [116] |
Table 2.
Summary of microRNAs for which lincRNAs in the 8q24.21 region act as a competitive endogenous RNA.
Table 2.
Summary of microRNAs for which lincRNAs in the 8q24.21 region act as a competitive endogenous RNA.
| 8q24.21 LncRNA | miRNAs and Affected Target Genes |
|---|---|
| PVT1 | miR-424-5p (CARM1) [119], miR-145 (FSCN1) [124], miR-143 (HK2) [125], miR-128-3p (GREM1) [126], miR-186 (Twist1) [127], miR-216b (Beclin-1) [128], miR-30a (IGF1) [129], miR-20a-5p (ULK1) [130], miR-29c (VEGF) [103], miR-26b (CTGF) [131], miR-140-5p (SMAD3) [132], miR-150 (HIG2) [133], miR-365 (ATG3) [112], miR-17-5p (PTEN) [134], miR-31 (CDK1) [110], miR-128 (VEGF) [135], miR-1207-5p (STAT6) [136] |
| CCAT1 | miR-181a-5p (HOXA1) [120], miR-143 (PLK1) [68], miR-148a (PKC) [56], miR-130a-3p (SOX4) [70], miR-218 (ZFX) [137], miR-148 (PIK3IP1) [71], miR-490-3p (CDK1) [138], miR-181a (CPEB2) [72], miR-218 (ZFX) [139] |
| PCAT1 | miR-128 (ZEB1) [121], miR-145-5p (FSCN1) [140], miR-129-5p (HMGB1) [141], miR-122 (WNT1) [42] |
| CCAT2 | miR-200b (VEGF) [122], miR-23b-5p (FOXC1) [142], miR-424 (VEGFA) [89] |
| CASC11 | miR-302 (CDK1) [123], miR-676-3p (NOL4L) [99], miR-498 (FOXK1) [102] |
| PRNCR1 | miR-944 (HOXB5) [53], miR-126-5p (MTDH) [52] |
| CASC19 | miR-140-5p (CEMIP) [50] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wilson, C.; Kanhere, A. 8q24.21 Locus: A Paradigm to Link Non-Coding RNAs, Genome Polymorphisms and Cancer. Int. J. Mol. Sci. 2021, 22, 1094. https://doi.org/10.3390/ijms22031094
AMA Style
Wilson C, Kanhere A. 8q24.21 Locus: A Paradigm to Link Non-Coding RNAs, Genome Polymorphisms and Cancer. International Journal of Molecular Sciences. 2021; 22(3):1094. https://doi.org/10.3390/ijms22031094
Chicago/Turabian StyleWilson, Claire, and Aditi Kanhere. 2021. "8q24.21 Locus: A Paradigm to Link Non-Coding RNAs, Genome Polymorphisms and Cancer" International Journal of Molecular Sciences 22, no. 3: 1094. https://doi.org/10.3390/ijms22031094
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.