
The Construction and Validation of a Novel Ferroptosis-Related Gene Signature in Parkinson’s Disease


Abstract
:1. Introduction
2. Results
2.1. Identification of Differential Genes in PD
2.2. Immune Infiltration Analysis and GSEA
2.3. WGCNA Identification of DEGs in PD
2.4. Identification of DEFRGs in PD
2.5. Functional Enrichment Analysis of DEFRGs
2.6. Prognostic Analysis
2.7. Single-Cell Sequencing Analysis of DEFRGs
2.8. Construction of the miRNA–Gene Network
2.9. In Vivo Experiment Verification
2.10. Prediction of Drug- and Protein-Binding Pockets
3. Materials and Methods
3.1. Data Acquisition
3.2. Immune Infiltration and GSEA
3.3. WGCNA Identification of DEGs in PD
3.4. Identification of DEFRGs in PD
3.5. Identification of Diagnostic Genes
3.6. Prediction of DEFRGs-Related miRNA and Protein-Binding Sites
3.7. In Vivo Experiment Verification
3.7.1. Animal
3.7.2. Western Blot and Immunohistochemistry Analysis
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolters, E.C. PD-related psychosis: Pathophysiology with therapeutical strategies. J. Neural Transm. Suppl. 2006, 71, 31–37. [Google Scholar] [CrossRef]
- Zhang, Y.-F.; Cragg, S.J. Revisiting dopamine-acetylcholine imbalance in Parkinson’s disease: Glutamate co-transmission as an exciting partner in crime. Neuron 2021, 109, 1070–1071. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Dhorajia, V.V.; Kim, J.; Kim, Y. Mitochondrial iron metabolism and neurodegenerative diseases. NeuroToxicology 2022, 88, 88–101. [Google Scholar] [CrossRef]
- Biondetti, E.; Santin, M.D.; Valabrègue, R.; Mangone, G.; Gaurav, R.; Pyatigorskaya, N.; Hutchison, M.; Yahia-Cherif, L.; Villain, N.; Habert, M.-O.; et al. The spatiotemporal changes in dopamine, neuromelanin and iron characterizing Parkinson’s disease. Brain 2021, 144, 3114–3125. [Google Scholar] [CrossRef] [PubMed]
- Bergsland, N.; Zivadinov, R.; Schweser, F.; Hagemeier, J.; Lichter, D.; Guttuso, T., Jr. Ventral posterior substantia nigra iron increases over 3 years in Parkinson’s disease. Mov. Disord. 2019, 34, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Langley, J.; Huddleston, D.E.; Chen, S.; Huang, P.; Ling, H.; Yan, F.; Hu, X. Increased iron-deposition in lateral-ventral substantia nigra pars compacta: A promising neuroimaging marker for Parkinson’s disease. NeuroImage Clin. 2020, 28, 102391. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-J.; Liu, X.-Y.; Xing, L.; Wan, X.; Chang, X.; Jiang, H.-L. Fenton reaction-independent ferroptosis therapy via glutathione and iron redox couple sequentially triggered lipid peroxide generator. Biomaterials 2020, 241, 119911. [Google Scholar] [CrossRef]
- Zuo, Y.; Xie, J.; Li, X.; Li, Y.; Thirupathi, A.; Zhang, J.; Yu, P.; Gao, G.; Chang, Y.; Shi, Z. Ferritinophagy-Mediated Ferroptosis Involved in Paraquat-Induced Neurotoxicity of Dopaminergic Neurons: Implication for Neurotoxicity in PD. Oxidative Med. Cell. Longev. 2021, 2021, 9961628. [Google Scholar] [CrossRef]
- Bisaglia, M.; Bubacco, L. Copper Ions and Parkinson’s Disease: Why Is Homeostasis So Relevant? Biomolecules 2020, 10, 195. [Google Scholar] [CrossRef]
- D’Ambrosi, N.; Rossi, L. Copper at synapse: Release, binding and modulation of neurotransmission. Neurochem. Int. 2015, 90, 36–45. [Google Scholar] [CrossRef]
- Dodani, S.C.; Domaille, D.W.; Nam, C.I.; Miller, E.W.; Finney, L.A.; Vogt, S.; Chang, C.J. Calcium-dependent copper redistributions in neuronal cells revealed by a fluorescent copper sensor and X-ray fluorescence microscopy. Proc. Natl. Acad. Sci. USA 2011, 108, 5980–5985. [Google Scholar] [CrossRef] [PubMed]
- Valensin, D.; Dell’Acqua, S.; Kozlowski, H.; Casella, L. Coordination and redox properties of copper interaction with α-synuclein. J. Inorg. Biochem. 2016, 163, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Davies, K.M.; Mercer, J.F.; Chen, N.; Double, K.L. Copper dyshomoeostasis in Parkinson’s disease: Implications for pathogenesis and indications for novel therapeutics. Clin. Sci. 2016, 130, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Huang, S.; Liu, Y.; Wan, C.; Gu, Y.; Wang, D.; Yu, H. Manganese promotes α-synuclein amyloid aggregation through the induction of protein phase transition. J. Biol. Chem. 2021, 298, 101469. [Google Scholar] [CrossRef] [PubMed]
- Clough, E.; Barrett, T. The Gene Expression Omnibus Database. Methods Mol. Biol. 2016, 1418, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Ren, J.; Pan, C.; Li, Y.; Xu, J.; Dong, H.; Chen, Y.; Liu, W. Serum miR-214 Serves as a Biomarker for Prodromal Parkinson’s Disease. Front. Aging Neurosci. 2021, 13, 700959. [Google Scholar] [CrossRef] [PubMed]
- Talepoor Ardakani, M.; Delavar, M.R.; Baghi, M.; Nasr-Esfahani, M.H.; Kiani-Esfahani, A.; Ghaedi, K. Upregulation of miR-200a and miR-204 in MPP+-treated differentiated PC12 cells as a model of Parkinson’s disease. Mol. Genet. Genom. Med. 2019, 7, e548. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, P.; Biswas, A.; Biswas, S.C. Brain-enriched miR-128: Reduced in exosomes from Parkinson’s patient plasma, improves synaptic integrity, and prevents 6-OHDA mediated neuronal apoptosis. Front. Cell. Neurosci. 2023, 16, 1037903. [Google Scholar] [CrossRef]
- Nordström, E.; Eriksson, F.; Sigvardson, J.; Johannesson, M.; Kasrayan, A.; Jones-Kostalla, M.; Appelkvist, P.; Söderberg, L.; Nygren, P.; Blom, M.; et al. ABBV-0805, a novel antibody selective for soluble aggregated α-synuclein, prolongs lifespan and prevents buildup of α-synuclein pathology in mouse models of Parkinson’s disease. Neurobiol. Dis. 2021, 161, 105543. [Google Scholar] [CrossRef]
- Pagano, G.; Taylor, K.I.; Anzures-Cabrera, J.; Marchesi, M.; Simuni, T.; Marek, K.; Postuma, R.B.; Pavese, N.; Stocchi, F.; Azulay, J.-P.; et al. Trial of Prasinezumab in Early-Stage Parkinson’s Disease. N. Engl. J. Med. 2022, 387, 421–432. [Google Scholar] [CrossRef]
- Lang, A.E.; Siderowf, A.D.; Macklin, E.A.; Poewe, W.; Brooks, D.J.; Fernandez, H.H.; Rascol, O.; Giladi, N.; Stocchi, F.; Tanner, C.M.; et al. Trial of Cinpanemab in Early Parkinson’s Disease. N. Engl. J. Med. 2022, 387, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Tosolini, M.; Kirilovsky, A.; Waldner, M.; Obenauf, A.C.; Angell, H.; Fredriksen, T.; Lafontaine, L.; Berger, A.; et al. Spatiotemporal Dynamics of Intratumoral Immune Cells Reveal the Immune Landscape in Human Cancer. Immunity 2013, 39, 782–795. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, L.; Wang, J.; Zhang, M.; Song, Z.; Ni, B.; You, Y. Identification of key biomarkers and immune infiltration in systemic lupus erythematosus by integrated bioinformatics analysis. J. Transl. Med. 2021, 19, 35, Erratum in J. Transl. Med. 2021, 19, 64. [Google Scholar] [CrossRef] [PubMed]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef] [PubMed]
- Elfil, M.; Kamel, S.; Kandil, M.; Koo, B.B.; Schaefer, S.M. Implications of the Gut Microbiome in Parkinson’s Disease. Mov. Disord. 2020, 35, 921–933. [Google Scholar] [CrossRef]
- Coles, L.D.; Tuite, P.J.; Öz, G.; Mishra, U.R.; Kartha, R.V.; Sullivan, K.M.; Cloyd, J.C.; Terpstra, M. Repeated-Dose Oral N-Acetylcysteine in Parkinson’s Disease: Pharmacokinetics and Effect on Brain Glutathione and Oxidative Stress. J. Clin. Pharmacol. 2018, 58, 158–167. [Google Scholar] [CrossRef]
- Grolez, G.; Moreau, C.; Sablonnière, B.; Garçon, G.; Devedjian, J.-C.; Meguig, S.; Gelé, P.; Delmaire, C.; Bordet, R.; Defebvre, L.; et al. Ceruloplasmin activity and iron chelation treatment of patients with Parkinson’s disease. BMC Neurol. 2015, 15, 74. [Google Scholar] [CrossRef]
- Wang, Z.-L.; Yuan, L.; Li, W.; Li, J.-Y. Ferroptosis in Parkinson’s disease: Glia–neuron crosstalk. Trends Mol. Med. 2022, 28, 258–269. [Google Scholar] [CrossRef]
- Ko, C.-J.; Gao, S.-L.; Lin, T.-K.; Chu, P.-Y.; Lin, H.-Y. Ferroptosis as a Major Factor and Therapeutic Target for Neuroinflammation in Parkinson’s Disease. Biomedicines 2021, 9, 1679. [Google Scholar] [CrossRef]
- Magalhaes, J.; Tresse, E.; Ejlerskov, P.; Hu, E.; Liu, Y.; Marin, A.; Montalant, A.; Satriano, L.; Rundsten, C.F.; Carlsen, E.M.M.; et al. PIAS2-mediated blockade of IFN-β signaling: A basis for sporadic Parkinson disease dementia. Mol. Psychiatry 2021, 26, 6083–6099. [Google Scholar] [CrossRef]
- Du, J.; Wang, T.; Li, Y.; Zhou, Y.; Wang, X.; Yu, X.; Ren, X.; An, Y.; Wu, Y.; Sun, W.; et al. DHA inhibits proliferation and induces ferroptosis of leukemia cells through autophagy dependent degradation of ferritin. Free Radic. Biol. Med. 2019, 131, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Nead, K.T.; Sinha, S.; Nguyen, P.L. Androgen deprivation therapy for prostate cancer and dementia risk: A systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2017, 20, 259–264. [Google Scholar] [CrossRef]
- Jeong, H.; Kim, M.-S.; Kwon, J.; Kim, K.-S.; Seol, W. Regulation of the transcriptional activity of the tyrosine hydroxylase gene by androgen receptor. Neurosci. Lett. 2006, 396, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Kelava, I.; Chiaradia, I.; Pellegrini, L.; Kalinka, A.T.; Lancaster, M.A. Androgens increase excitatory neurogenic potential in human brain organoids. Nature 2022, 602, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Mahoney-Sánchez, L.; Bouchaoui, H.; Ayton, S.; Devos, D.; Duce, J.A.; Devedjian, J.-C. Ferroptosis and its potential role in the physiopathology of Parkinson’s Disease. Prog. Neurobiol. 2020, 196, 101890. [Google Scholar] [CrossRef] [PubMed]
- Benskey, M.J.; Perez, R.G.; Manfredsson, F.P. The contribution of alpha synuclein to neuronal survival and function—Implications for Parkinson’s disease. J. Neurochem. 2016, 137, 331–359. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, J.; Kuang, F.; Chen, X.; Zeh, H.J.; Kang, R.; Kroemer, G.; Xie, Y.; Tang, D. PDK4 dictates metabolic resistance to ferroptosis by suppressing pyruvate oxidation and fatty acid synthesis. Cell Rep. 2021, 34, 108767. [Google Scholar] [CrossRef]
- Rahman, H.; Jha, M.K.; Kim, J.-H.; Nam, Y.; Lee, M.G.; Go, Y.; Harris, R.A.; Park, D.H.; Kook, H.; Lee, I.-K.; et al. Pyruvate Dehydrogenase Kinase-mediated Glycolytic Metabolic Shift in the Dorsal Root Ganglion Drives Painful Diabetic Neuropathy. J. Biol. Chem. 2016, 291, 6011–6025. [Google Scholar] [CrossRef]
- Weisberg, L.S. Lactic Acidosis in a Patient with Type 2 Diabetes Mellitus. Clin. J. Am. Soc. Nephrol. 2015, 10, 1476–1483. [Google Scholar] [CrossRef]
- Sotelo-Hitschfeld, T.; Niemeyer, M.I.; Mächler, P.; Ruminot, I.; Lerchundi, R.; Wyss, M.T.; Stobart, J.; Fernández-Moncada, I.; Valdebenito, R.; Garrido-Gerter, P.; et al. Channel-Mediated Lactate Release by K+-Stimulated Astrocytes. J. Neurosci. 2015, 35, 4168–4178. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Name | Biological Function | Expression Variance | Main Location |
|---|---|---|---|---|
| BEX1 | Brain-Expressed X-Linked Protein 1 | BEX1 is involved in the regulation of neuronal differentiation and apoptosis through the P75 neurotrophin receptor pathway. | Down |  |
| CIRBP | Cold-Inducible RNA-Binding Protein | CIRBP regulates cell proliferation, development, apoptosis, differentiation, biological rhythm regulation, and inflammation. | Down |  |
| CP | Ceruloplasmin | It exhibits ferroxidase activity and oxidizes Fe2+ to Fe3+ without releasing free radical oxygen. | Up | - |
| DNAJB6 | DnaJ Heat Shock Protein Family (HSP40) Member B6 | Having stimulatory effects on the atpase activity of HSP70 in a dose-dependent and time-dependent manner and therefore acting as cochaperones of HSP70, members of this family play a specific role in neuronal aggregation of polyglutamine. | Down |  |
| GCH1 | GTP Cyclohydrolase 1 | GCH1 is the most important rate-limiting enzyme in BH4 synthesis, and the decrease in GCH1 activity leads to a decrease in BH4 production. It is also a major regulator of peripheral neuropathic and inflammatory pain, and gene mutations may reduce pain sensitivity. It is involved in dopamine synthesis. | Down |  |
| HSPB1 | Heat shock protein family B | It is transported from the cytoplasm to the nucleus upon stress induction and acts as a molecular chaperone to promote the proper folding of other proteins. It can regulate many biological processes through its molecular chaperone activity, including phosphorylation of neurofilament proteins and axonal transport. | Up |  |
| LAMP2 | Lysosomal-Associated Membrane Protein 2 | LAMP2 plays a role in lysosomal protection, maintenance, and adhesion, regulating chaperone-mediated autophagy. | Up |  |
| MT1G | Metallothionein 1G | MT1G is involved in the response of cells to metal ions. Cellular response to vascular endothelial growth factor stimulation, negative regulation of growth. | Up | - |
| TF | Transferrin | Responsible for transporting iron from sites of absorption and heme degradation to sites of storage and utilization, stimulating cell proliferation. | Up | - |
| YTHDC2 | YTH N6-Methyladenosine RNA Binding Protein C2 | RNA helicases play a role in the efficiency and stability of RNA processing by specifically recognizing and binding N6-methyladenosine (m6A)-containing RNA by facilitating the transition from mitosis to meiosis in stem cells. | Down | 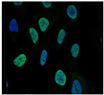 |
| GFRA1 | Glial cell line-derived neurotrophic factor Family Receptor Alpha 1 | GFRA1 encodes a member of the glial cell line-derived neurotrophic factor receptor (GDNFR) family of proteins. Glial cell line-derived neurotrophic factor (GDNF) and neurotrophin (NTN) are two structurally related potent neurotrophic factors that play a key role in controlling neuronal survival and differentiation. | Down | 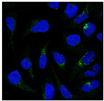 |
| GSK3B | Glycogen Synthase Kinase 3 Beta | Negative regulator of glucose homeostasis involved in energy metabolism, inflammation, endoplasmic reticulum stress, mitochondrial dysfunction, and apoptotic pathways. GSK3B deficiency has been implicated in PD and Alzheimer’s disease. | Down |  |
| LIFR | LIF Receptor Subunit Alpha | LIFR encodes a protein belonging to the type I cytokine receptor family that mediates the action of leukemia-inhibitory factors, which are involved in cell differentiation, proliferation, and survival in adults and embryos. | Up | - |
| ADAM23 | ADAM Metallopeptidase Domain 23 | Members of this family are membrane-anchored proteins that are structurally related to snake venom disintegrin and are implicated in various biological processes involving cell–cell and cell–matrix interactions, including fertilization, muscle development, and neurogenesis. | Down | - |
| PDK4 | Pyruvate Dehydrogenase Kinase 4 | The activity of pyruvate dehydrogenase complex (PDC) is inactivated and tricarboxylic acid cycle and lipid biosynthesis are blocked. | Up | - |
| SNCA | Alpha-Synuclein | Regulates synaptic vesicle trafficking and subsequent neurotransmitter release. SNCA can be used to integrate presynaptic signaling and membrane trafficking. SNCA deficiency has been implicated in the pathogenesis of PD. | Down | - |
| CYB5R1 | Cytochrome B5 Reductase 1 | REDOX proteins catalyze the peroxidation of lipids containing long-chain phospholipids of unsaturated fatty acids, causing oxidative damage and leakage of liposome membranes. They can transfer electrons from NADPH/NADH to downstream proteins to mediate REDOX reactions. | Down |  |
| ALOX15 | Arachidonate 15-Lipoxygenase | ALOX15 encodes a member of the lipoxygenase protein family. The encoded enzymes act on various polyunsaturated fatty acid substrates to produce various bioactive lipid mediators that can regulate inflammation and immunity. | Up | - |
| MAPK1 | Mitogen-Activated Protein Kinase 1 | It serves as an integration point for a variety of biochemical signals and is involved in various cellular processes such as proliferation, differentiation, transcriptional regulation, and development. | Down |  |
| KDM5A | Lysine Demethylase 5A | As a histone demethylase, the encoded protein demethylates lysine 4 of histone H3 through histone coding, thus playing a role in gene regulation and closely related to tumor proliferation. | Up |  |
| TYRO3 | Tyrosine-Protein Kinase Receptor | TYRO3 is involved in the control of cell survival, migration and differentiation, spermatogenesis, immune regulation, and phagocytosis. | Up | - |
| YAP1 | Yes1-Associated Transcriptional Regulator | Transcriptional regulators, key downstream regulatory targets in the Hippo signaling pathway, play a key role in organ size control and tumor suppression by limiting proliferation and promoting apoptosis. | Up |  |
| CDO1 | Cysteine Dioxygenase Type 1 | Taurine is a key enzyme that is synthesized by the organism itself. Taurine is abundant and widely distributed in the brain, which can significantly promote the growth and development of the nervous system and cell proliferation and differentiation, and it plays an important role in the development of brain nerve cells. It is involved in L-cysteine catabolism. | Down |  |
| ISCU | Iron-Sulfur Cluster Assembly Enzyme | Enzymes that regulate metabolism, iron homeostasis, and oxidative stress responses. | Down |  |
| PANX1 | Pannexin 1 | Structural components of gap junctions and hemichannes, involved in ATP release and nucleotide permeation. It can act as a Ca2+ leakage channel to regulate endoplasmic reticulum Ca2+ homeostasis. | Down |  |
| AR | Androgen Receptor | Ligand-activated transcription factors that regulate eukaryotic gene expression and affect cell proliferation and differentiation in target tissues, physiological concentrations of testosterone induce neuroprotective effects through AR. | Down |  |
| Protein Positive Degree Proportion (%) | High Positive | Positive | Low Positive | Negative | |
|---|---|---|---|---|---|
| Protein | |||||
| AR | Saline | 51.51 ± 0.65 | 45.70 ± 0.87 | 2.25 ± 0.11 | 0.54 ± 0.19 |
| MPTP | 38.54 ± 1.12 *** | 55.31 ± 1.19 ** | 3.21 ± 0.11 ** | 2.93 ± 0.37 ** | |
| ISCU | Saline | 52.26 ± 1.16 | 43.52 ± 1.22 | 3.20 ± 0.10 | 1.02 ± 0.24 |
| MPTP | 10.02 ± 0.29 *** | 80.33 ± 0.48 *** | 9.41 ± 0.34 ** | 0.25 ± 0.075 * | |
| SNCA | Saline | 27.60 ± 0.66 | 60.50 ± 0.40 | 10.65 ± 0.29 | 1.24 ± 0.28 |
| MPTP | 47.00 ± 0.84 *** | 46.26 ± 0.89 *** | 6.32 ± 0.13 *** | 0.42 ± 0.10 * | |
| PDK4 | Saline | 16.11 ± 0.33 | 79.56 ± 0.47 | 3.75 ± 0.48 | 0.59 ± 0.31 |
| MPTP | 31.07 ± 0.20 *** | 63.51 ± 0.62 *** | 5.01 ± 0.54 | 0.41 ± 0.17 | |
| Protein | Name | Volume Å3 | Surface Å2 | Drug Score | Simple Score |
|---|---|---|---|---|---|
| PDK4 (2zkj) | P_3 | 535.61 | 618.17 | 0.89 | 0.25 |
| P_9 | 301.66 | 393.37 | 0.84 | 0.15 | |
| P_0 | 939.76 | 1168.93 | 0.83 | 0.64 | |
| P_1 | 841.81 | 920.6 | 0.81 | 0.57 | |
| P_2 | 551.09 | 639.63 | 0.81 | 0.29 | |
| P_6 | 387.17 | 498.25 | 0.79 | 0.19 | |
| P_7 | 330.28 | 444.89 | 0.71 | 0.06 | |
| P_5 | 388.33 | 549.11 | 0.67 | 0.17 | |
| P_4 | 392.29 | 612.63 | 0.65 | 0.15 | |
| P_8 | 310.97 | 447.95 | 0.55 | 0.29 | |
| SNCA (1xq8) | P_2 | 130.6 | 321.36 | 0.2 | 0.04 |
| P_1 | 321.36 | 762.65 | 0.37 | 0.22 | |
| P_0 | 406.65 | 887.92 | 0.55 | 0.32 | |
| ISCU (7c8m) | P_0 | 1158.91 | 1285.98 | 0.81 | 0.6 |
| P_1 | 383.3 | 674.44 | 0.81 | 0.24 | |
| P_2 | 367.74 | 669.91 | 0.74 | 0.22 | |
| P_3 | 314.82 | 427.98 | 0.52 | 0.23 | |
| P_4 | 141.31 | 341.21 | 0.35 | 0.0 | |
| P_5 | 140.03 | 305.42 | 0.34 | 0.0 | |
| P_6 | 109.18 | 284.76 | 0.24 | 0.0 | |
| AR (1r4i) | P_0 | 848.19 | 1180.27 | 0.79 | 0.57 |
| P_1 | 130.43 | 290.87 | 0.26 | 0 | |
| P_2 | 112.51 | 212.86 | 0.17 | 0 | |
| P_3 | 106.56 | 327.51 | 0.22 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Wu, H.; Wei, J. The Construction and Validation of a Novel Ferroptosis-Related Gene Signature in Parkinson’s Disease. Int. J. Mol. Sci. 2023, 24, 17203. https://doi.org/10.3390/ijms242417203
Liu T, Wu H, Wei J. The Construction and Validation of a Novel Ferroptosis-Related Gene Signature in Parkinson’s Disease. International Journal of Molecular Sciences. 2023; 24(24):17203. https://doi.org/10.3390/ijms242417203
Chicago/Turabian StyleLiu, Tingting, Haojie Wu, and Jianshe Wei. 2023. "The Construction and Validation of a Novel Ferroptosis-Related Gene Signature in Parkinson’s Disease" International Journal of Molecular Sciences 24, no. 24: 17203. https://doi.org/10.3390/ijms242417203
APA StyleLiu, T., Wu, H., & Wei, J. (2023). The Construction and Validation of a Novel Ferroptosis-Related Gene Signature in Parkinson’s Disease. International Journal of Molecular Sciences, 24(24), 17203. https://doi.org/10.3390/ijms242417203