
Deflamin Attenuated Lung Tissue Damage in an Ozone-Induced COPD Murine Model by Regulating MMP-9 Catalytic Activity

 , , , , , , , , and
, , , , , , , , and
Abstract
:1. Introduction
2. Results
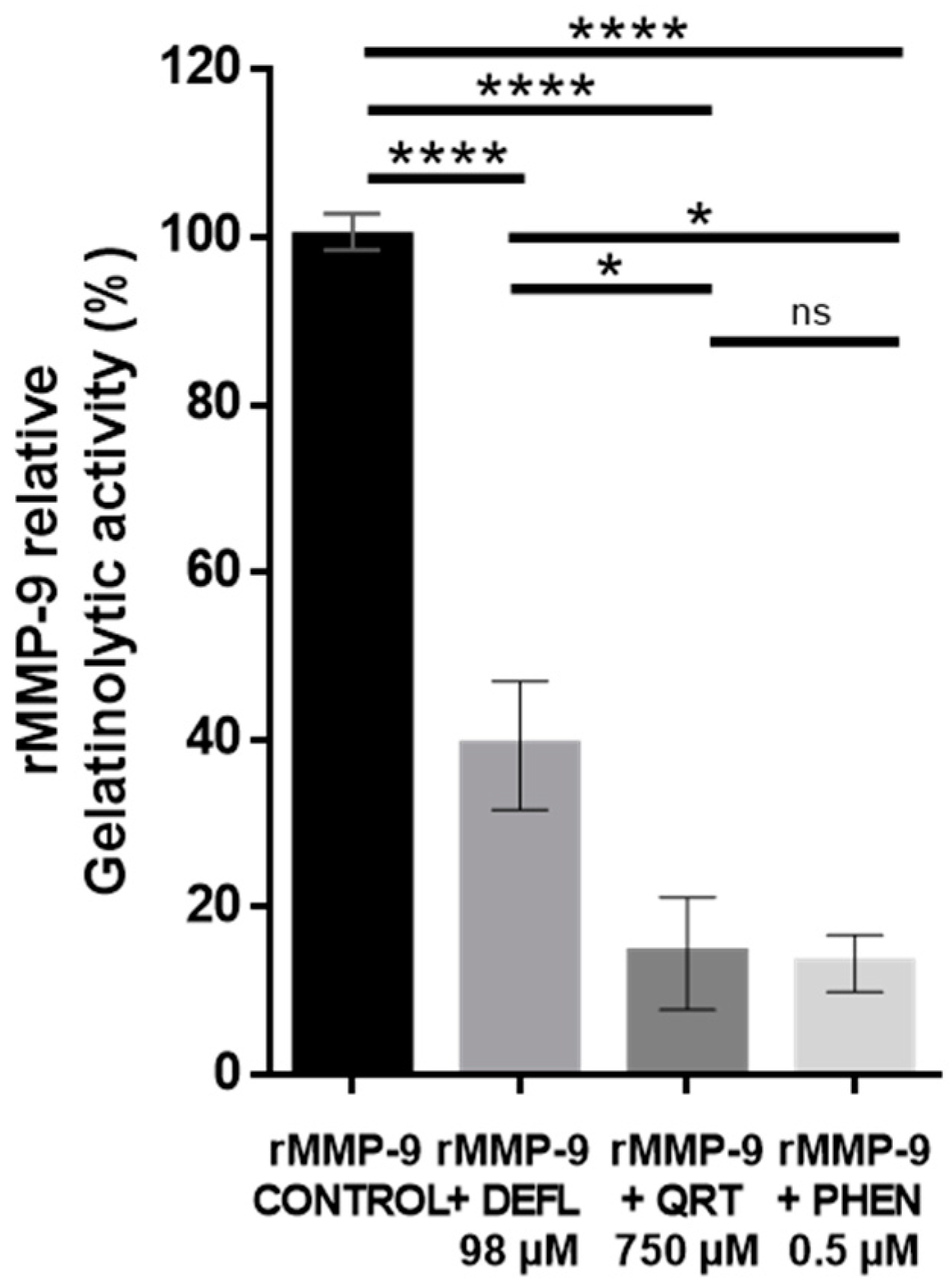
2.1. In Vitro Inhibitory Effect of Deflamin on MMP-9 Activity Using the DQ-Gelatin Assay
2.2. Attenuation of Lung Damage after Deflamin Treatment in Chronic Ozone-Exposed Mice
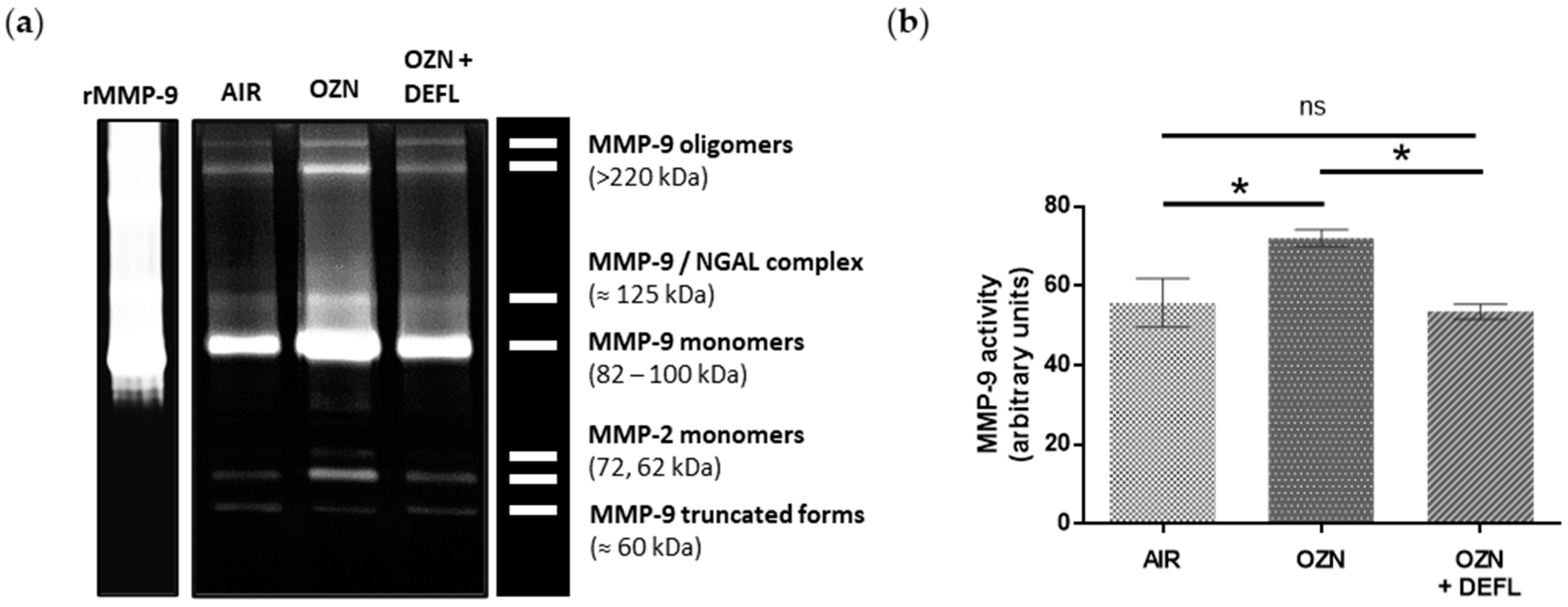
2.3. Restoration of MMP-9 Protein Expression in Lung Tissue from Treated Animals
2.4. Inhibition of the MMP-9 Activity in Deflamin-Treated Mice
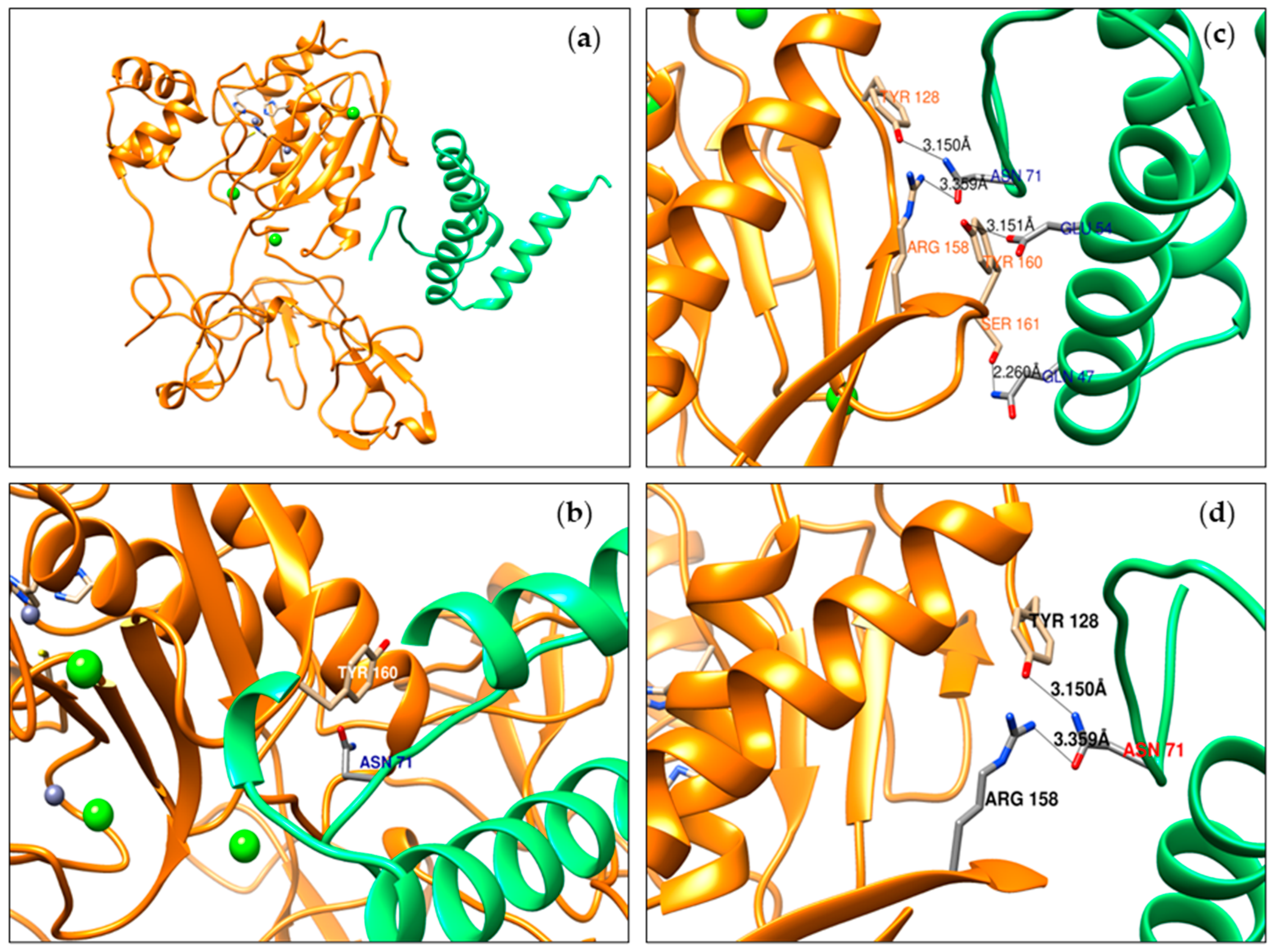
2.5. Prediction of Deflamin Fragments and MMP-9 Interaction
3. Discussion
4. Materials and Methods
4.1. Plant Material and Isolation of Deflamin
4.2. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis
4.3. Fluorogenic DQ™-Gelatin Assay
4.4. Animals and Experimental Design
4.5. Tissue Samples
4.6. Lung Histological Assessment
4.7. Western Blot Analysis
4.8. Gelatin Zymography
4.9. Docking of Deflamin Fragments and MMP-9
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Initiative for Chronic Obstructive Lung Disease Global Strategy for Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease (2023 Report). Available online: https://goldcopd.org/2023-gold-report-2/ (accessed on 5 December 2023).
- GBD 2019 Chronic Respiratory Disease Collaborators. Global burden of chronic respiratory diseases and risk factors, 1990–2019: An update from the Global Burden of Disease Study 2019. EClinicalMedicine 2023, 59, 101936. [Google Scholar] [CrossRef] [PubMed]
- Mumby, S.; Chung, K.F.; Adcock, I.M. Transcriptional Effects of Ozone and Impact on Airway Inflammation. Front. Immunol. 2019, 10, 1610. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Hanaoka, M.; Droma, Y.; Chen, P.; Voelkel, N.F.; Kubo, K. Endothelin-1 receptor antagonists prevent the development of pulmonary emphysema in rats. Eur. Respir. J. 2010, 35, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, P.; Zhang, M.; Liang, L.; Sun, X.; Li, M.; Tang, Y.; Bao, A.; Gong, J.; Zhang, J.; et al. Hydrogen Sulfide Prevents and Partially Reverses Ozone-Induced Features of Lung Inflammation and Emphysema in Mice. Am. J. Respir. Cell Mol. Biol. 2016, 55, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Maeno, T.; Ota, F.; Ueno, M.; Korekane, H.; Takamatsu, S.; Shirato, K.; Matsumoto, A.; Kobayashi, S.; Yoshida, K.; et al. Sensitivity of heterozygous α1,6-fucosyltransferase knock-out mice to cigarette smoke-induced emphysema: Implication of aberrant transforming growth factor-β signaling and matrix metalloproteinase gene expression. J. Biol. Chem. 2012, 287, 16699–16708. [Google Scholar] [CrossRef] [PubMed]
- Araya, J.; Tsubouchi, K.; Sato, N.; Ito, S.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; Kadota, T.; et al. PRKN-regulated mitophagy and cellular senescence during COPD pathogenesis. Autophagy 2019, 15, 510–526. [Google Scholar] [CrossRef] [PubMed]
- Woldhuis, R.R.; de Vries, M.; Timens, W.; van den Berge, M.; Demaria, M.; Oliver, B.G.G.; Heijink, I.H.; Brandsma, C.A. Link between increased cellular senescence and extracellular matrix changes in COPD. Am. J. Physiol. Lung Cell. Mol. Physiol. 2020, 319, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.; Guan, Y.; Shen, H.J.; Zhang, L.H.; Jiang, J.X.; Dong, X.W.; Shen, H.H.; Xie, Q.M. Akt/PKB signaling regulates cigarette smoke-induced pulmonary epithelial-mesenchymal transition. Lung Cancer 2018, 122, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.; Yang, Y.; Zhou, X.; Zhang, J.; Li, B.; Wang, X.; Luo, X. Overexpression of the hyperplasia suppressor gene inactivates airway fibroblasts obtained from a rat model of chronic obstructive pulmonary disease by inhibiting the Wnt signaling pathway. Mol. Med. Rep. 2019, 20, 2754–2762. [Google Scholar] [CrossRef] [PubMed]
- Houghton, A.M. Matrix metalloproteinases in destructive lung disease. Matrix Biol. 2015, 44–46, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Vlahos, R.; Wark, P.A.B.; Anderson, G.P.; Bozinovski, S. Glucocorticosteroids differentially regulate MMP-9 and neutrophil elastase in COPD. PLoS ONE 2012, 7, e33277. [Google Scholar] [CrossRef] [PubMed]
- Perotin, J.M.; Adam, D.; Vella-Boucaud, J.; Delepine, G.; Sandu, S.; Jonvel, A.C.; Prevost, A.; Berthiot, G.; Pison, C.; Lebargy, F.; et al. Delay of airway epithelial wound repair in COPD is associated with airflow obstruction severity. Respir. Res. 2014, 15, 151. [Google Scholar] [CrossRef] [PubMed]
- Linder, R.; Rönmark, E.; Pourazar, J.; Behndig, A.; Blomberg, A.; Lindberg, A. Serum metalloproteinase-9 is related to COPD severity and symptoms-cross-sectional data from a population based cohort-study. Respir. Res. 2015, 16, 28. [Google Scholar] [CrossRef]
- Bchir, S.; Nasr, H.B.; Bouchet, S.; Benzarti, M.; Garrouch, A.; Tabka, Z.; Susin, S.; Chahed, K.; Bauvois, B. Concomitant elevations of MMP-9, NGAL, proMMP-9/NGAL and neutrophil elastase in serum of smokers with chronic obstructive pulmonary disease. J. Cell. Mol. Med. 2017, 21, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Sundar, I.K.; Hwang, J.W.; Wu, S.; Sun, J.; Rahman, I. Deletion of vitamin D receptor leads to premature emphysema/COPD by increased matrix metalloproteinases and lymphoid aggregates formation. Biochem. Biophys. Res. Commun. 2011, 406, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chen, H.; Xu, W.; Zhang, W.; Buckley, S.; Zheng, S.G.; Warburton, D.; Kolb, M.; Gauldie, J.; Shi, W. Molecular mechanisms of MMP9 overexpression and its role in emphysema pathogenesis of Smad3-deficient mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Hwang, J.W.; Sundar, I.K.; Friedman, A.E.; McBurney, M.W.; Guarente, L.; Gu, W.; Kinnula, V.L.; Rahman, I. SIRT1 redresses the imbalance of tissue inhibitor of matrix metalloproteinase-1 and matrix metalloproteinase-9 in the development of mouse emphysema and human COPD. Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Li, S.; Zou, W.; Hu, G.; Zhou, Y.; Peng, G.; He, F.; Li, B.; Ran, P. Upregulation of gelatinases and epithelial-mesenchymal transition in small airway remodeling associated with chronic exposure to wood smoke. PLoS ONE 2014, 9, e96708. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Xie, G.; Ding, F.; Zhou, X. LPS-induced MMP-9 expression is mediated through the MAPKs-AP-1 dependent mechanism in BEAS-2B and U937 cells. Exp. Lung Res. 2018, 44, 217–225. [Google Scholar] [CrossRef]
- Zhou, L.; Le, Y.; Tian, J.; Yang, X.; Jin, R.; Gai, X.; Sun, Y. Cigarette smoke-induced RANKL expression enhances MMP-9 production by alveolar macrophages. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 14, 81–91. [Google Scholar] [CrossRef]
- Bidan, C.M.; Veldsink, A.C.; Meurs, H.; Gosens, R. Airway and Extracellular Matrix Mechanics in COPD. Front. Physiol. 2015, 6, 346. [Google Scholar] [CrossRef] [PubMed]
- Swetha, R.; Gayen, C.; Kumar, D.; Singh, T.D.; Modi, G.; Singh, S.K. Biomolecular basis of matrix metallo proteinase-9 activity. Future Med. Chem. 2018, 10, 1093–1112. [Google Scholar] [CrossRef] [PubMed]
- Gearing, A.J.; Beckett, P.; Christodoulou, M.; Churchill, M.; Clements, J.; Davidson, A.H.; Drummond, A.H.; Galloway, W.A.; Gilbert, R.; Gordon, J.L.; et al. Processing of tumour necrosis factor-alpha precursor by metalloproteinases. Nature 1994, 370, 555–557. [Google Scholar] [CrossRef] [PubMed]
- Schönbeck, U.; Mach, F.; Libby, P. Generation of biologically active IL-1 beta by matrix metalloproteinases: A novel caspase-1-independent pathway of IL-1 beta processing. J. Immunol. 1998, 161, 3340–3346. [Google Scholar] [CrossRef]
- Perng, D.W.; Chang, K.T.; Su, K.C.; Wu, Y.C.; Chen, C.S.; Hsu, W.H.; Tsai, C.M.; Lee, Y.C. Matrix metalloprotease-9 induces transforming growth factor-β(1) production in airway epithelium via activation of epidermal growth factor receptors. Life Sci. 2011, 89, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Noerager, B.D.; Xu, X.; Davis, V.A.; Jones, C.W.; Okafor, S.; Whitehead, A.; Blalock, J.E.; Jackson, P.L. A Potential Role for Acrolein in Neutrophil-Mediated Chronic Inflammation. Inflammation 2015, 38, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Karakiulakis, G.; Batzios, S.; Savic, S.; Roth, M.; Tamm, M.; Stolz, D. Acute exacerbations of COPD are associated with significant activation of matrix metalloproteinase 9 irrespectively of airway obstruction, emphysema and infection. Respir. Res. 2015, 16, 78. [Google Scholar] [CrossRef]
- Wells, J.M.; Parker, M.M.; Oster, R.A.; Bowler, R.P.; Dransfield, M.T.; Bhatt, S.P.; Cho, M.H.; Kim, V.; Curtis, J.L.; Martinez, F.J.; et al. Elevated circulating MMP-9 is linked to increased COPD exacerbation risk in SPIROMICS and COPDGene. JCI Insight 2018, 3, e123614. [Google Scholar] [CrossRef]
- Lima, A.I.G.; Mota, J.; Monteiro, S.A.V.S.; Ferreira, R.M.S.B. Legume seeds and colorectal cancer revisited: Protease inhibitors reduce MMP-9 activity and colon cancer cell migration. Food Chem. 2016, 197, 30–38. [Google Scholar] [CrossRef]
- Mota, J.; Direito, R.; Rocha, J.; Fernandes, J.; Sepodes, B.; Figueira, M.E.; Raymundo, A.; Lima, A.; Ferreira, R.B. Lupinus albus Protein Components Inhibit MMP-2 and MMP-9 Gelatinolytic Activity In Vitro and In Vivo. Int. J. Mol. Sci. 2021, 22, 13286. [Google Scholar] [CrossRef] [PubMed]
- Mota, J.; Casimiro, S.; Fernandes, J.; Hartmann, R.M.; Schemitt, E.; Picada, J.; Costa, L.; Marroni, N.; Raymundo, A.; Lima, A. Lupin Protein Concentrate as a Novel Functional Food Additive That Can Reduce Colitis-Induced Inflammation and Oxidative Stress. Nutrients 2022, 14, 2102. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Cavaco, A.; Basso, B.; Mota, J.; Cruz-Duarte, R.; Costa, M.; Carvalho, L.; Lima, A.; Costa, L.; Ferreira, R.; et al. Therapeutic Potential of Deflamin against Colorectal Cancer Development and Progression. Cancers 2022, 14, 6182. [Google Scholar] [CrossRef] [PubMed]
- Mota, J.; Figueira, M.E.; Ferreira, R.B.; Lima, A. An Up-Scalable and Cost-Effective Methodology for Isolating a Polypeptide Matrix Metalloproteinase-9 Inhibitor from Lupinus albus Seeds. Foods 2021, 10, 1663. [Google Scholar] [CrossRef]
- Triantaphyllopoulos, K.; Hussain, F.; Pinart, M.; Zhang, M.; Li, F.; Adcock, I.; Kirkham, P.; Zhu, J.; Chung, K.F. A model of chronic inflammation and pulmonary emphysema after multiple ozone exposures in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2011, 300, 691–700. [Google Scholar] [CrossRef]
- Michaudel, C.; Fauconnier, L.; Julé, Y.; Ryffel, B. Functional and morphological differences of the lung upon acute and chronic ozone exposure in mice. Sci. Rep. 2018, 8, 10611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bao, W.; Fei, X.; Zhang, Y.; Zhang, G.; Zhou, X.; Zhang, M. Progesterone attenuates airway remodeling and glucocorticoid resistance in a murine model of exposing to ozone. Mol. Immunol. 2018, 96, 69–77. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhao, Y.; Wang, Q.; Chen, H.; Zhou, X. Fine particulate matter exposure promotes M2 macrophage polarization through inhibiting histone deacetylase 2 in the pathogenesis of chronic obstructive pulmonary disease. Ann. Transl. Med. 2020, 8, 1303. [Google Scholar] [CrossRef] [PubMed]
- Vandooren, J.; Van den Steen, P.E.; Opdenakker, G. Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): The next decade. Crit. Rev. Biochem. Mol. Biol. 2013, 48, 222–272. [Google Scholar] [CrossRef]
- Begum, H.; Murugesan, P.; Tangutur, A.D. Western blotting: A powerful staple in scientific and biomedical research. Biotechniques 2022, 73, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.K.; Cho, H.Y.; Kleeberger, S.R. Protective role of matrix metalloproteinase-9 in ozone-induced airway inflammation. Environ. Health Perspect. 2007, 115, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.; Liu, Q.; Han, F.; Gu, W.; Song, L.; Zhang, Y.; Guo, X.; Xu, W. Ginsenoside Rg1 Ameliorates Cigarette Smoke-Induced Airway Fibrosis by Suppressing the TGF-β1/Smad Pathway In Vivo and In Vitro. Biomed. Res. Int. 2017, 2017, 6510198. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.W.; Seo, C.S.; Shin, N.R.; Kim, J.S.; Lee, S.I.; Kim, J.C.; Kim, S.H.; Shin, I.S. Modificated Mahuang-Tang, a traditional herbal medicine suppresses inflammatory responses induced by cigarette smoke in human airway epithelial cell and mice. Phytomedicine 2019, 59, 152777. [Google Scholar] [CrossRef] [PubMed]
- Higham, A.; Rattray, N.J.W.; Dewhurst, J.A.; Trivedi, D.K.; Fowler, S.J.; Goodacre, R.; Singh, D. Electronic cigarette exposure triggers neutrophil inflammatory responses. Respir. Res. 2016, 17, 56. [Google Scholar] [CrossRef] [PubMed]
- Paemen, L.; Jansen, P.M.; Proost, P.; Van Damme, J.; Opdenakker, G.; Hack, E.; Taylor, F.B. Induction of gelatinase B and MCP-2 in baboons during sublethal and lethal bacteraemia. Cytokine 1997, 9, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.; Ng, T.S.Y.; Rezaeizadeh, A.; Aamidor, S.; Twigg, S.M.; Min, D.; McLennan, S.V. Insulin treatment prevents wounding associated changes in tissue and circulating neutrophil MMP-9 and NGAL in diabetic rats. PLoS ONE 2017, 12, e0170951. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, S.P.; Li, H.; Taniguchi, G.T. Zymography and reverse zymography for detecting MMPs and TIMPs. Methods Mol. Biol. 2010, 622, 257–269. [Google Scholar] [PubMed]
- Wojtowicz-Praga, S.; Torri, J.; Johnson, M.; Steen, V.; Marshall, J.; Ness, E.; Dickson, R.; Sale, M.; Rasmussen, H.S.; Chiodo, T.A.; et al. Phase I trial of Marimastat, a novel matrix metalloproteinase inhibitor, administered orally to patients with advanced lung cancer. J. Clin. Oncol. 1998, 16, 2150–2156. [Google Scholar] [CrossRef]
- Amigo, L.; Hernández-Ledesma, B. Current Evidence on the Bioavailability of Food Bioactive Peptides. Molecules 2020, 25, 4479. [Google Scholar] [CrossRef]
- Wells, J.M.; Gaggar, A.; Blalock, J.E. MMP generated matrikines. Matrix Biol. 2015, 44–46, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Saragusti, A.C.; Ortega, M.G.; Cabrera, J.L.; Estrin, D.A.; Marti, M.A.; Chiabrando, G.A. Inhibitory effect of quercetin on matrix metalloproteinase 9 activity molecular mechanism and structure-activity relationship of the flavonoid-enzyme interaction. Eur. J. Pharmacol. 2010, 644, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Vandooren, J.; Geurts, N.; Martens, E.; Van den Steen, P.E.; Jonghe, S.D.; Herdewijn, P.; Opdenakker, G. Gelatin degradation assay reveals MMP-9 inhibitors and function of O-glycosylated domain. World J. Biol. Chem. 2011, 2, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Ringland, C.; Schweig, J.E.; Eisenbaum, M.; Paris, D.; Ait-Ghezala, G.; Mullan, M.; Crawford, F.; Abdullah, L.; Bachmeier, C. MMP9 modulation improves specific neurobehavioral deficits in a mouse model of Alzheimer’s disease. BMC Neurosci. 2021, 22, 39. [Google Scholar] [CrossRef] [PubMed]
- Elkins, P.A.; Ho, Y.S.; Smith, W.W.; Janson, C.A.; D’Alessio, K.J.; McQueney, M.S.; Cummings, M.D.; Romanic, A.M. Structure of the C-terminally truncated human ProMMP9, a gelatin-binding matrix metalloproteinase. Acta Crystallogr. D Biol. Crystallogr. 2002, 58, 1182–1192. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nandy, S.K.; Chowdhury, A.; Chakraborti, T.; Chakraborti, S. Inhibition of MMP-9 by green tea catechins and prediction of their interaction by molecular docking analysis. Biomed. Pharmacother. 2016, 84, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.B.; Wang, W.; Gao, J.M.; Li, F.; Shi, J.S.; Gong, Q.H. Icariside II attenuates cerebral ischemia/reperfusion-induced blood-brain barrier dysfunction in rats via regulating the balance of MMP9/TIMP1. Acta Pharmacol. Sin. 2020, 41, 1547–1556. [Google Scholar] [CrossRef] [PubMed]
- Pluda, S.; Mazzocato, Y.; Angelini, A. Peptide-Based Inhibitors of ADAM and ADAMTS Metalloproteinases. Front. Mol. Biosci. 2021, 8, 703715. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Nery-Flores, S.D.; Mendoza-Magaña, M.L.; Ramírez-Herrera, M.A.; Ramírez-Vázquez, J.J.; Romero-Prado, M.M.J.; Cortez-Álvarez, C.R.; Ramírez-Mendoza, A.A. Curcumin Exerted Neuroprotection against Ozone-Induced Oxidative Damage and Decreased NF-κB Activation in Rat Hippocampus and Serum Levels of Inflammatory Cytokines. Oxid. Med. Cell. Longev. 2018, 2018, 9620684. [Google Scholar] [CrossRef] [PubMed]
- Marcos, J.V.; Muñoz-Barrutia, A.; Ortiz-de-Solórzano, C.; Cristóbal, G. Quantitative Assessment of Emphysema Severity in Histological Lung Analysis. Ann. Biomed. Eng. 2015, 43, 2515–2529. [Google Scholar] [CrossRef] [PubMed]
- Alzoubi, A.; Ghazwi, R.; Alzoubi, K.; Alqudah, M.; Kheirallah, K.; Khabour, O.; Allouh, M. Vascular endothelial growth factor receptor inhibition enhances chronic obstructive pulmonary disease picture in mice exposed to waterpipe smoke. Folia Morphol. 2018, 77, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Eurlings, I.M.; Dentener, M.A.; Mercken, E.M.; de Cabo, R.; Bracke, K.R.; Vernooy, J.H.; Wouters, E.F.M.; Reynaert, N. A comparative study of matrix remodeling in chronic models for COPD; mechanistic insights into the role of TNF-α. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Chen, J.; Khalil, R.A. Zymography as a Research Tool in the Study of Matrix Metalloproteinase Inhibitors. Methods Mol. Biol. 2017, 1626, 79–102. [Google Scholar] [PubMed]
- Wilkesman, J.; Kurz, L. Zymography Principles. Methods Mol. Biol. 2017, 1626, 3–10. [Google Scholar]
- Yang, J.; Zhang, Y. I-TASSER server: New development for protein structure and function predictions. Nucleic Acids Res. 2015, 43, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Freddolino, P.L.; Zhang, Y. COFACTOR: Improved protein function prediction by combining structure, sequence and protein-protein interaction information. Nucleic Acids Res. 2017, 45, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Zhang, C.; Li, Y.; Pearce, R.; Bell, E.W.; Zhang, Y. Folding non-homologous proteins by coupling deep-learning contact maps with I-TASSER assembly simulations. Cell Rep. Methods 2021, 1, 100014. [Google Scholar] [CrossRef] [PubMed]
- Heo, L.; Park, H.; Seok, C. GalaxyRefine: Protein structure refinement driven by side-chain repacking. Nucleic Acids Res. 2013, 41, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.R.; Heo, L.; Seok, C. Effective protein model structure refinement by loop modeling and overall relaxation. Proteins 2016, 84, 293–301. [Google Scholar] [CrossRef]
- Lovell, S.C.; Davis, I.W.; Arendall, W.B., 3rd; de Bakker, P.I.; Word, J.M.; Prisant, M.G.; Richardson, J.S.; Richardson, D.C. Structure validation by Calpha geometry: Phi, psi and Cbeta deviation. Proteins 2003, 50, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Tao, H.; He, J.; Huang, S.Y. The HDOCK server for integrated protein-protein docking. Nat. Protoc. 2020, 15, 1829–1852. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MMP-9 | δ-Conglutin Large Subunit | Distance (Å) |
|---|---|---|
| TYR 128 (C) | ASN 71 | 3.150 |
| ARG 158 (C) | ASN 71 | 3.359 |
| TYR 160 (C) | GLU 54 | 3.151 |
| SER 161 (C) | GLN 47 | 2.260 |
| ARG 162 (C) | GLU 46 | 2.772 |
| ASP 163 (C) | ARG 43 | 2.863 |
| ASP 280 (FN) | ARG 27 | 2.677 |
| ASP 284 (FN) | ARG 23 | 2.938 |
| TYR 311 (FN) | GLU 75 | 2.478 |
| MMP-9 | δ-Conglutin Large Subunit | Distance (Å) |
|---|---|---|
| ARG 134 (5) | ASN 71 (7) | 4.832 |
| ARG 134 (5) | ASP 73 (4) | 4.086 |
| ARG 134 (5) | GLU 75 (4) | 2.626 |
| TYR 160 (11) | GLN 47 (5) | 4.842 |
| TYR 160 (11) | ASN 69 (5) | 3.149 |
| TYR 160 (11) | ASN 71 (7) | 2.797 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltazar-García, E.A.; Vargas-Guerrero, B.; Lima, A.; Boavida Ferreira, R.; Mendoza-Magaña, M.L.; Ramírez-Herrera, M.A.; Baltazar-Díaz, T.A.; Domínguez-Rosales, J.A.; Salazar-Montes, A.M.; Gurrola-Díaz, C.M. Deflamin Attenuated Lung Tissue Damage in an Ozone-Induced COPD Murine Model by Regulating MMP-9 Catalytic Activity. Int. J. Mol. Sci. 2024, 25, 5063. https://doi.org/10.3390/ijms25105063
Baltazar-García EA, Vargas-Guerrero B, Lima A, Boavida Ferreira R, Mendoza-Magaña ML, Ramírez-Herrera MA, Baltazar-Díaz TA, Domínguez-Rosales JA, Salazar-Montes AM, Gurrola-Díaz CM. Deflamin Attenuated Lung Tissue Damage in an Ozone-Induced COPD Murine Model by Regulating MMP-9 Catalytic Activity. International Journal of Molecular Sciences. 2024; 25(10):5063. https://doi.org/10.3390/ijms25105063
Chicago/Turabian StyleBaltazar-García, Elia Ana, Belinda Vargas-Guerrero, Ana Lima, Ricardo Boavida Ferreira, María Luisa Mendoza-Magaña, Mario Alberto Ramírez-Herrera, Tonatiuh Abimael Baltazar-Díaz, José Alfredo Domínguez-Rosales, Adriana María Salazar-Montes, and Carmen Magdalena Gurrola-Díaz. 2024. "Deflamin Attenuated Lung Tissue Damage in an Ozone-Induced COPD Murine Model by Regulating MMP-9 Catalytic Activity" International Journal of Molecular Sciences 25, no. 10: 5063. https://doi.org/10.3390/ijms25105063