
Diversity of Bacterial Communities in Sediment in Inland Water Bodies in Relation to Environmental Factors and Human Impacts: A Case Study on Typical Regions in Vietnam
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Sample Processing and DNA Extraction
2.3. 16S rRNA Gene Targeted Sequencing
2.4. Bacterial Diversity Analyses
2.5. Analyses of the Correlations between Bacterial Diversity and Environmental and Anthropogenic Factors
3. Results
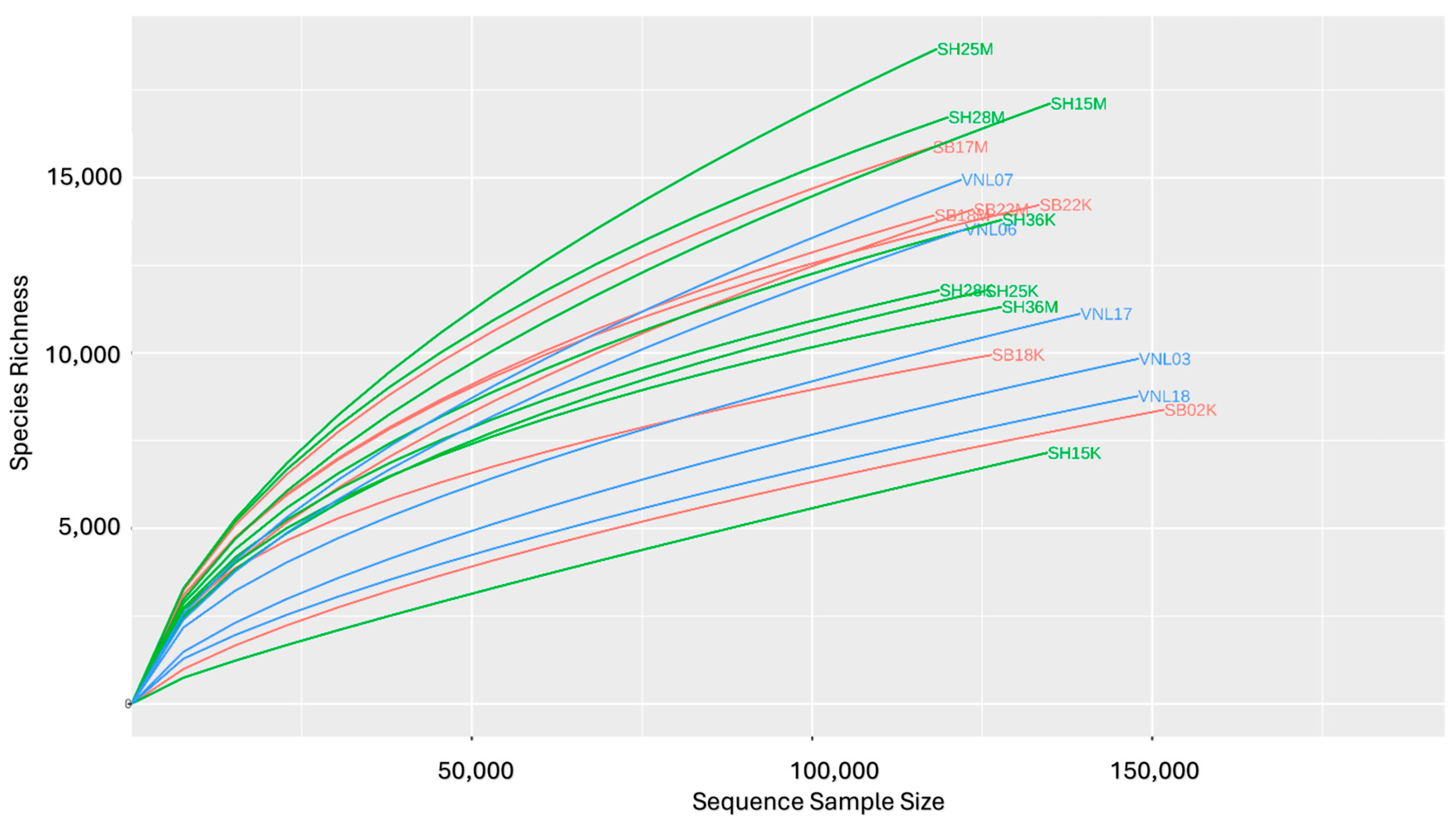
3.1. Sequencing Data Quality
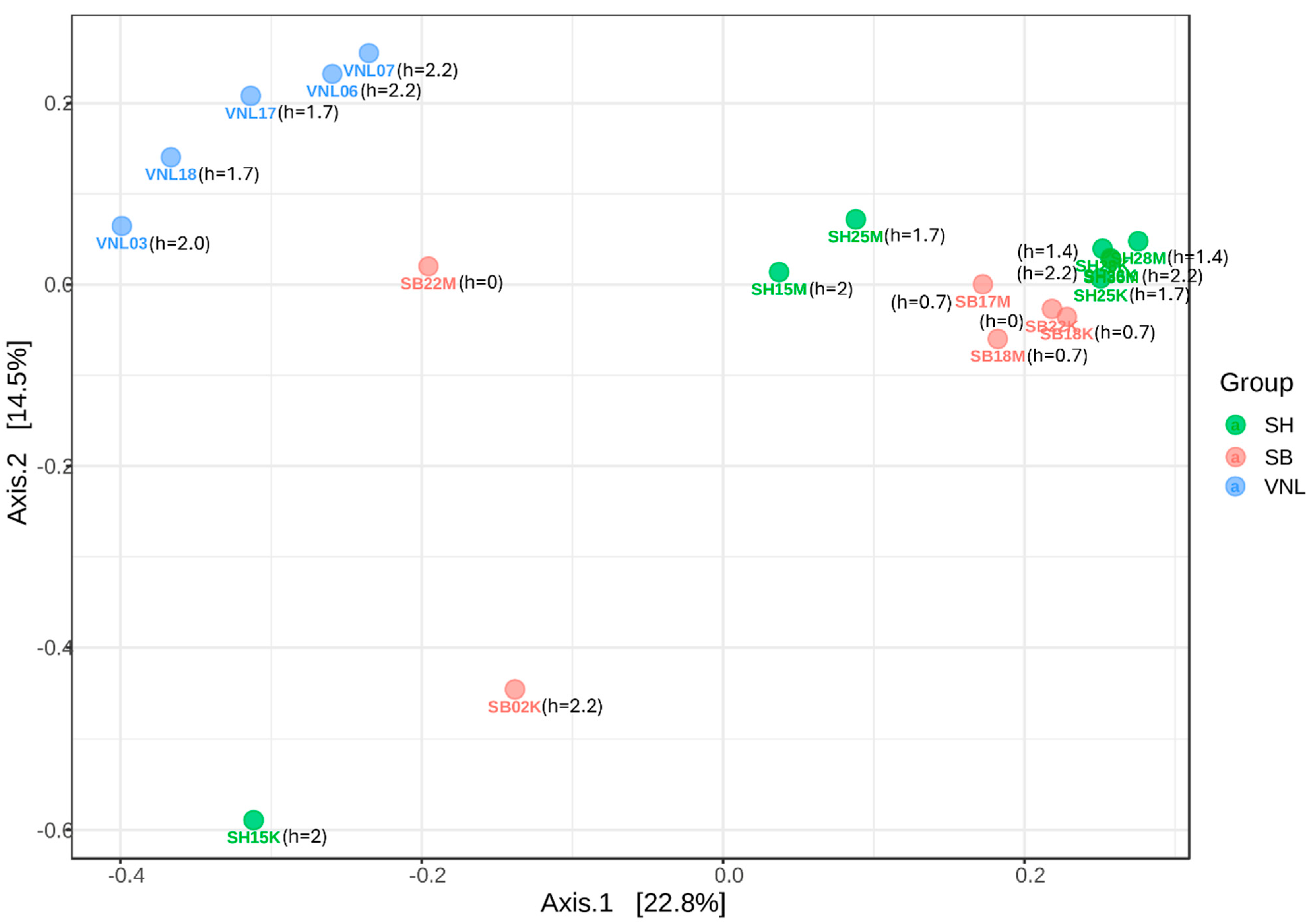
3.2. Bacterial Diversity
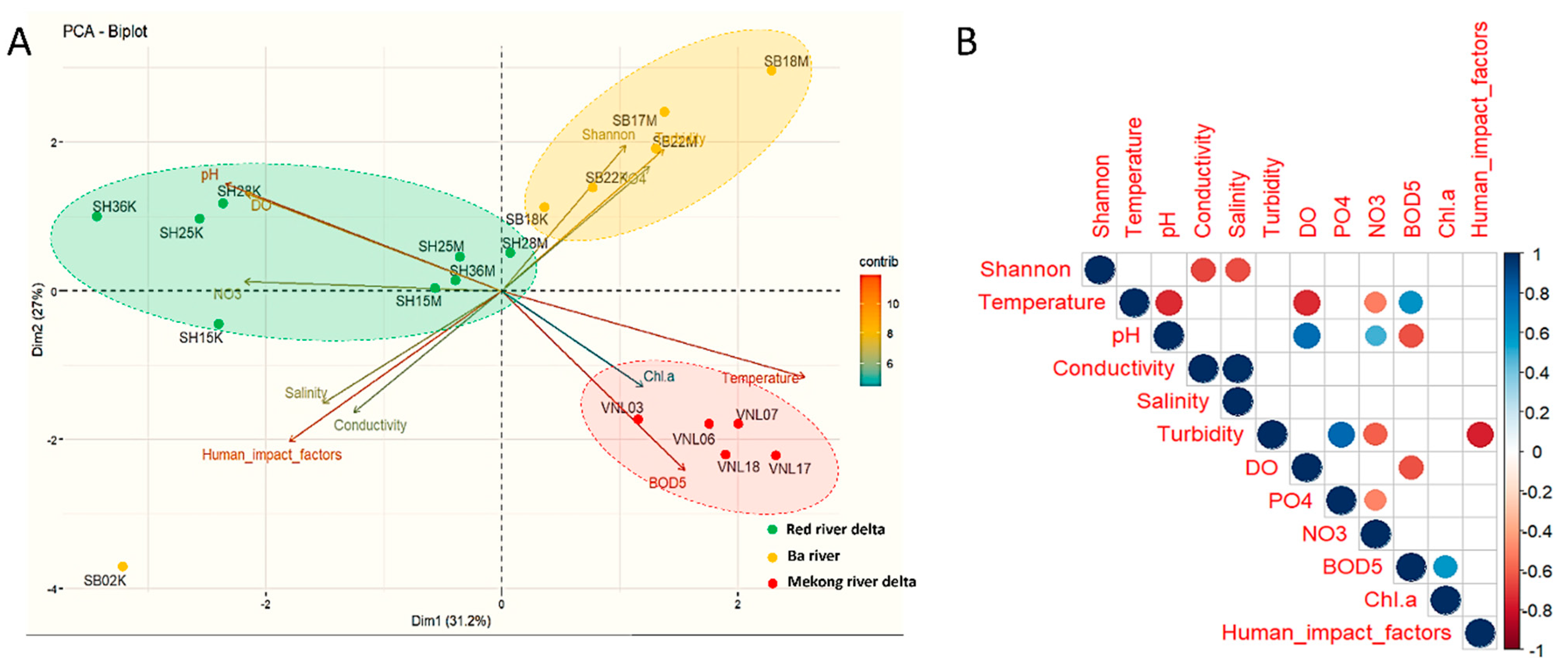
3.3. Bacterial Diversity vs. Environmental Factors
3.4. Bacterial Diversity vs. Human Impact
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bandh, S.A.; Parray, J.A.; Shameem, N. Climate Change and Microbial Diversity: Advances and Challenges; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Zhang, Y.; Dong, S.; Gao, Q.; Liu, S.; Zhou, H.; Ganjurjav, H.; Wang, X. Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total. Environ. 2016, 562, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.K.; Purkayastha, S.D.; De Mandal, S.; Passari, A.K.; Govindarajan, R.K. Effect of climate change on microbial diversity and its functional attributes. In Recent Advancements in Microbial Diversity; Elsevier: Amsterdam, The Netherlands, 2020; pp. 315–331. [Google Scholar]
- Jansson, J.K.; Hofmockel, K.S. Soil microbiomes and climate change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhang, Y.; Guo, X.; Ning, D.; Zhou, X.; Feng, J.; Yuan, M.M.; Liu, S.; Guo, J.; Gao, Z.; et al. Reduction of microbial diversity in grassland soil is driven by long-term climate warming. Nat. Microbiol. 2022, 7, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, T.; Wang, Y.; Cheng, H.; Chang, S.X.; Liang, C.; An, S. Negative effects of multiple global change factors on soil microbial diversity. Soil Biol. Biochem. 2021, 156, 108229. [Google Scholar] [CrossRef]
- Pold, G.; DeAngelis, K.M. Up against the wall: The effects of climate warming on soil microbial diversity and the potential for feedbacks to the carbon cycle. Diversity 2013, 5, 409–425. [Google Scholar] [CrossRef]
- Blaud, A.; Lerch, T.Z.; Phoenix, G.K.; Osborn, A.M. Arctic soil microbial diversity in a changing world. Res. Microbiol. 2015, 166, 796–813. [Google Scholar] [CrossRef] [PubMed]
- Makhalanyane, T.P.; Van Goethem, M.W.; Cowan, D.A. Microbial diversity and functional capacity in polar soils. Curr. Opin. Biotechnol. 2016, 38, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Moreno, J.; Harrison, I.J.; Dudgeon, D.; Clausnitzer, V.; Darwall, W.; Farrell, T.; Savy, C.; Tockner, K.; Tubbs, N. Sustaining Freshwater Biodiversity in the Anthropocene; Springer: Berlin/Heidelberg, Germany, 2014; pp. 247–270. [Google Scholar]
- Capon, S.J.; Chambers, L.E.; Mac Nally, R.; Naiman, R.J.; Davies, P.; Marshall, N.; Pittock, J.; Reid, M.; Capon, T.; Douglas, M.; et al. Riparian ecosystems in the 21st century: Hotspots for climate change adaptation? Ecosystems 2013, 16, 359–381. [Google Scholar] [CrossRef]
- Dinesen, L.; Joosten, H.; Rochefort, L.; Lindsay, R.; Glatzel, S. Ramsar Policy Brief No. 5. Restoring drained peatlands: A necessary step to achieve global climate goals. In Convention on Wetlands; Secretariat of the Convention on Wetlands: Gland, Switzerland, 2021; Available online: https://repository.uel.ac.uk/download/15eac57c406a86031ea0a7545bd4631f8f70921f1f0d1a057b443f5bc7d69c42/1267307/rpb5_restoring_drained_peatlands_e.pdf (accessed on 15 February 2024).
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.; Cumberlidge, N.; Darwall, W.R.; Pollock, C.; Richman, N.I.; Soulsby, A.M.; Böhm, M.; et al. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef]
- Thieme, M.L.; Tickner, D.; Grill, G.; Carvallo, J.P.; Goichot, M.; Hartmann, J.; Higgins, J.; Lehner, B.; Mulligan, M.; Nilsson, C. Navigating trade-offs between dams and river conservation. Glob. Sustain. 2021, 4, e17. [Google Scholar] [CrossRef]
- Esdaile, L.J.; Chalker, J.M. The mercury problem in artisanal and small-scale gold mining. Chemistry 2018, 24, 6905–6916. [Google Scholar] [CrossRef] [PubMed]
- Chagnon, M.; Kreutzweiser, D.; Mitchell, E.A.; Morrissey, C.A.; Noome, D.A.; Van der Sluijs, J.P. Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ. Sci. Pollut. Res. 2015, 22, 119–134. [Google Scholar] [CrossRef] [PubMed]
- Zeglin, L.H. Stream microbial diversity in response to environmental changes: Review and synthesis of existing research. Front. Microbiol. 2015, 6, 128252. [Google Scholar] [CrossRef] [PubMed]
- Retter, A.; Karwautz, C.; Griebler, C. Groundwater microbial communities in times of climate change. Curr. Issues Mol. Biol. 2021, 41, 509–538. [Google Scholar] [CrossRef] [PubMed]
- Muyzer, G. DGGE/TGGE a method for identifying genes from natural ecosystems. Curr. Opin. Microbiol. 1999, 2, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Nam, N.N.; Do, H.D.K.; Loan Trinh, K.T.; Lee, N.Y. Metagenomics: An effective approach for exploring microbial diversity and functions. Foods 2023, 12, 2140. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, N.S.; Posa, M.R.C.; Lee, T.M.; Bickford, D.; Koh, L.P.; Brook, B.W. The state and conservation of Southeast Asian biodiversity. Biodivers. Conserv. 2010, 19, 317–328. [Google Scholar] [CrossRef]
- Nakayama, T.; Hoa, T.T.T.; Harada, K.; Warisaya, M.; Asayama, M.; Hinenoya, A.; Lee, J.W.; Phu, T.M.; Ueda, S.; Sumimura, Y.; et al. Water metagenomic analysis reveals low bacterial diversity and the presence of antimicrobial residues and resistance genes in a river containing wastewater from backyard aquacultures in the Mekong Delta, Vietnam. Environ. Pollut. 2017, 222, 294–306. [Google Scholar] [CrossRef]
- Wada, M.; Mori, F.; Yokouchi, K.; Yagi, M.; Takita, T.; Ishimatsu, A.; Iwataki, M.; Takahashi, K.; Van Mai, H.; Vo, T.T. Comparison of planktonic microbial abundance and dissolved oxygen consumption between the aquaculture ponds of mudskippers and shrimps in the Mekong Delta, southern Vietnam. Fish Sci. 2016, 82, 787–797. [Google Scholar] [CrossRef]
- Wilbers, G.-J.; Becker, M.; Sebesvari, Z.; Renaud, F.G. Spatial and temporal variability of surface water pollution in the Mekong Delta, Vietnam. Sci. Total. Environ. 2014, 485, 653–665. [Google Scholar] [CrossRef]
- Pham, T.-L.; Dao, T.-S.; Tran, N.-D.; Nimptsch, J.; Wiegand, C.; Motoo, U. Influence of environmental factors on cyanobacterial biomass and microcystin concentration in the Dau Tieng Reservoir, a tropical eutrophic water body in Vietnam. Ann. Limnol.-Int. J. Limnol. 2017, 53, 89–100. [Google Scholar] [CrossRef]
- Petro, C.; Starnawski, P.; Schramm, A.; Kjeldsen, K.U. Microbial community assembly in marine sediments. Aquat. Microb. Ecol. 2017, 79, 177–195. [Google Scholar] [CrossRef]
- Mac, T.M.T. A Review on Stream and River Ecosystems in Vietnam: Common and Specific Characteristics by Their Natural Geographical Conditions (in Vietnamese). 2008. Available online: http://thongtinkhcn.vinhlong.gov.vn:81/kqncvn2012/Bao_ve_moi_truong/Toan_van/7629-3.pdf (accessed on 15 February 2024).
- Li, Z.; Saito, Y.; Matsumoto, E.; Wang, Y.; Tanabe, S.; Vu, Q.L. Climate change and human impact on the Song Hong (Red River) Delta, Vietnam, during the Holocene. Quat. Int. 2006, 144, 4–28. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Dang, X.P.; Eiji, M.; Jiang, H. Characteristics of vegetation types and distributions around the Red River, Vietnam. Guihaia 2009, 29, 744–750. [Google Scholar]
- Pham, Q.S. Assessment of the Impacts of the Downstream Flow Rate of Ba River on the Accretion in the Estuary Area of Da Dien River, Phu Yen Province (in Vietnamese). Master’s Thesis, VNU University of Science—Vietnam National University in Hanoi, Hanoi, Vietnam, 2018. [Google Scholar]
- Xuan, N.H.; Van, N.K.; Oanh, H.T.K.; Van Vu, V. The creation of bioclimatic vegetation map to develop sustainable agro forestry in ba and kone river basin, vietnam. Ukr. Geogr. J. 2021, N1, 54–62. [Google Scholar] [CrossRef]
- Le, T.; Cần, T.; Du, L.; Ngọc, P.; Thường, V.; Toan, T.; Lợi, T. Summary of Several Acts to Cope with Clime Change in Mekong Delta Area (in Vietnamese); Project Report of Sustainable Rural Development Centre and Climate Change Research Institute at Can Tho University: Can Tho, Vietnam, 2013. [Google Scholar]
- Dang, A.T.; Kumar, L.; Reid, M.; Anh, L.N. Modelling the susceptibility of wetland plant species under climate change in the Mekong Delta, Vietnam. Ecol. Informatics 2021, 64, 101358. [Google Scholar] [CrossRef]
- Greenberg, A.; Clesceri, L.S.; Eaton, A.D. Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Trinh, H.P.; Lee, S.-H.; Kim, N.-K.; Yoon, H.; Jeong, G.; Jung, Y.-J.; Hur, M.; Lee, B.-H.; Park, H.-D. Enrichment of Ca. Jettenia in sequencing batch reactors operated with low nitrogen loading rate and high influent nitrogen concentration. Bioresour. Technol. 2022, 352, 127079. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Kassambara, A. Factoextra: Extract and visualize the results of multivariate data analyses [R package factoextra version 1.0. 7]. Comprehensive R Archive Network (CRAN), 2020.
- Islam, M.; Shafi, S.; Bandh, S.A.; Shameem, N. Impact of environmental changes and human activities on bacterial diversity of lakes. In Freshwater Microbiology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 105–136. [Google Scholar]
- Vineis, P.; Chan, Q.; Khan, A. Climate change impacts on water salinity and health. J. Epidemiol. Glob. Health 2011, 1, 5–10. [Google Scholar] [CrossRef]
- Olson, J.R. Predicting combined effects of land use and climate change on river and stream salinity. Philos. Trans. R. Soc. Lond. 2019, 374, 20180005. [Google Scholar] [CrossRef]
- Vu, D.T.; Yamada, T.; Ishidaira, H. Assessing the impact of sea level rise due to climate change on seawater intrusion in Mekong Delta, Vietnam. Water Sci. Technol. 2018, 77, 1632–1639. [Google Scholar] [CrossRef]
- Huang, L.; Bai, J.; Wen, X.; Zhang, G.; Zhang, C.; Cui, B.; Liu, X. Microbial resistance and resilience in response to environmental changes under the higher intensity of human activities than global average level. Glob. Chang. Biol. 2020, 26, 2377–2389. [Google Scholar] [CrossRef]
- Antiqueira, P.A.P.; Petchey, O.L.; Dos Santos, V.P.; de Oliveira, V.M.; Romero, G.Q. Environmental change and predator diversity drive alpha and beta diversity in freshwater macro and microorganisms. Glob. Chang. Biol. 2018, 24, 3715–3728. [Google Scholar] [CrossRef]
- Sun, W.; Xia, C.; Xu, M.; Guo, J.; Sun, G. Seasonality affects the diversity and composition of bacterioplankton communities in Dongjiang River, a drinking water source of Hong Kong. Front. Microbiol. 2017, 8, 1644. [Google Scholar] [CrossRef]
- Ortiz-Vera, M.P.; Olchanheski, L.R.; da Silva, E.G.; de Lima, F.R.; Martinez, L.R.d.P.R.; Sato, M.I.Z.; Jaffé, R.; Alves, R.; Ichiwaki, S.; Padilla, G.; et al. Influence of water quality on diversity and composition of fungal communities in a tropical river. Sci. Rep. 2018, 8, 14799. [Google Scholar] [CrossRef]
- Feng, B.-W.; Li, X.-R.; Wang, J.-H.; Hu, Z.-Y.; Meng, H.; Xiang, L.-Y.; Quan, Z.-X. Bacterial diversity of water and sediment in the Changjiang estuary and coastal area of the East China Sea. FEMS Microbiol. Ecol. 2009, 70, 236–248. [Google Scholar] [CrossRef]
- Campbell, B.J.; Kirchman, D.L. Bacterial diversity, community structure and potential growth rates along an estuarine salinity gradient. ISME J. 2013, 7, 210–220. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Hou, Y. A new cold-adapted and salt-tolerant glutathione reductase from Antarctic psychrophilic bacterium Psychrobacter sp. and its resistance to oxidation. Int. J. Mol. Sci. 2020, 21, 420. [Google Scholar] [CrossRef]
- Gan, L.; Li, X.; Tian, Y.; Peng, B. Genomic insights into the salt tolerance and cold adaptation of Planococcus halotolerans SCU63 T. Arch. Microbiol. 2020, 202, 2841–2847. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Region | Season | Temperature (°C) | pH | Conductivity (mS m−1) | Salinity (%) | Turbidity (NTU) | [DO] (mg L−1) | [PO43−] (mg L−1) | [NO3−] (mg L−1) | BOD5 (mg L−1) | Chl.a (mg L−1) | H. Impact Score |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SH15K | Red river | dry | 23.1 | 7.85 | 20 | 0.01 | 19 | 5.65 | 0.027 | 0.747 | 1.5 | 0.218 | 2.0 |
| SH15M | Red river | rainy | 28.78 | 7.17 | 19.6 | 0.01 | 90.4 | 4.58 | 0.093 | 1.032 | 2.2 | 0.193 | 2.0 |
| SH25K | Red river | dry | 21.5 | 7.99 | 21 | 0.01 | 26 | 6.23 | 0.069 | 0.988 | 1.1 | 0.155 | 1.7 |
| SH25M | Red river | rainy | 30.28 | 7.78 | 19.3 | 0.01 | 60.1 | 4.05 | 0.057 | 0.704 | 0.6 | 0.089 | 1.7 |
| SH28K | Red river | dry | 20.5 | 8.01 | 20 | 0.01 | 59 | 6.15 | 0.05 | 0.923 | 1.5 | 0.171 | 1.4 |
| SH28M | Red river | rainy | 28.72 | 7.05 | 23.5 | 0.01 | 84.7 | 4.17 | 0.057 | 0.749 | 0.5 | 0.133 | 1.4 |
| SH36K | Red river | dry | 20 | 8.21 | 22 | 0.1 | 37 | 6.73 | 0.108 | 1.07 | 0.7 | 0.228 | 2.2 |
| SH36M | Red river | rainy | 27.93 | 7.07 | 24.1 | 0.01 | 70 | 5.19 | 0.098 | 0.729 | 1.5 | 1.962 | 2.2 |
| SB02K | Ba river | dry | 24.9 | 7.41 | 1200 | 0.5 | 103 | 5.3 | 0.044 | 0.462 | 4.2 | 0.415 | 2.2 |
| SB17M | Ba river | rainy | 28.92 | 7.59 | 12 | 0.01 | 524 | 6.49 | 0.332 | 0.089 | 1.1 | 4.294 | 0.7 |
| SB18K | Ba river | dry | 25.1 | 6.83 | 11 | 0.01 | 240 | 4.66 | 0.058 | 0.447 | 0.05 | 0.472 | 0.7 |
| SB18M | Ba river | rainy | 27.8 | 7.44 | 10 | 0 | 581 | 4.5 | 0.965 | 0.128 | 2.05 | 0.2 | 0.7 |
| SB22K | Ba river | dry | 24.5 | 6.83 | 10 | 0.01 | 252 | 4.52 | 0.104 | 0.402 | 0.95 | 0.259 | 0 |
| SB22M | Ba river | rainy | 27.43 | 7.3 | 11.1 | 0.01 | 522 | 3.23 | 0.301 | 0.652 | 1.6 | 0.12 | 0 |
| VNL03 | MD flood zone | 29.6 | 7.17 | 16 | 0.01 | 243 | 5.7 | 0.1 | 0.4 | 6.2 | 26.5 | 2.0 | |
| VNL06 | MD flood zone | 31.4 | 6.6 | 33 | 0.01 | 41 | 1.65 | 0.11 | 0.5 | 6 | 4.5 | 2.2 | |
| VNL07 | MD flood zone | 31.6 | 6.68 | 34 | 0.01 | 40 | 1.65 | 0.06 | 0.49 | 6.8 | 6.35 | 2.2 | |
| VNL17 | MD flood zone | 31.2 | 6.6 | 34 | 0.01 | 41 | 1.82 | 0.08 | 0.51 | 9.4 | 8.55 | 1.7 | |
| VNL18 | MD flood zone | 31.3 | 6.3 | 25 | 0.01 | 43 | 3.35 | 0.09 | 0.33 | 7.9 | 4.5 | 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bui, H.T.; Nguyen, T.T.T.; Le, H.T.; Pham, D.T.; Nguyen, H.Q.; Nguyen, N.T.; Nguyen, T.D.; Nguyen, T.T.; Trinh, H.P.; Duong, C.V.; et al. Diversity of Bacterial Communities in Sediment in Inland Water Bodies in Relation to Environmental Factors and Human Impacts: A Case Study on Typical Regions in Vietnam. Microbiol. Res. 2024, 15, 864-879. https://doi.org/10.3390/microbiolres15020056
Bui HT, Nguyen TTT, Le HT, Pham DT, Nguyen HQ, Nguyen NT, Nguyen TD, Nguyen TT, Trinh HP, Duong CV, et al. Diversity of Bacterial Communities in Sediment in Inland Water Bodies in Relation to Environmental Factors and Human Impacts: A Case Study on Typical Regions in Vietnam. Microbiology Research. 2024; 15(2):864-879. https://doi.org/10.3390/microbiolres15020056
Chicago/Turabian StyleBui, Hoa Thi, Thuy Thi Thu Nguyen, Ha Thu Le, Dau Thi Pham, Huy Quang Nguyen, Nam Thanh Nguyen, Tam Duc Nguyen, Tam Thi Nguyen, Hoang Phuc Trinh, Cuong Van Duong, and et al. 2024. "Diversity of Bacterial Communities in Sediment in Inland Water Bodies in Relation to Environmental Factors and Human Impacts: A Case Study on Typical Regions in Vietnam" Microbiology Research 15, no. 2: 864-879. https://doi.org/10.3390/microbiolres15020056