
Soybean CEP6 Signaling Peptides Positively Regulate Nodulation
by
 ,
,
Shuai Wu
†,
Xiaoli Wang
†,
Jie Qin
†,
Wenqing Tian
,
Min Wang
,
Aiqin Yue
,
Lixiang Wang
,
Weijun Du
* and
Jinzhong Zhao
Houji Labortary in Shanxi Province, College of Agriculture, Shanxi Agricultural University, Jinzhong 030801, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Agronomy 2024, 14(5), 988; https://doi.org/10.3390/agronomy14050988
Submission received: 5 March 2024
/
Revised: 11 April 2024
/
Accepted: 4 May 2024
/
Published: 8 May 2024
(This article belongs to the Section Plant-Crop Biology and Biochemistry)
Abstract
:Nodulation is the most efficient nitrate assimilation system in the ecosystem, while excessive fertilization has an increased nitrate inhibition effect; deciphering the nitrate signal transduction mechanism in the process is of the utmost importance. In this study, genome-wide analyses of the GmCEP genes were applied to identify nodulation-related CEP genes; 22 GmCEP family members were identified, while GmCEP6 was mainly expressed in nodules and significantly responded to nitrate treatment and rhizobium infection, especially in later stages. Overexpression and CRISPR-Cas9 were used to validate its role in nodulation. We found that GmCEP6 overexpression significantly increased the nodule number, while GmCEP6 knock-out significantly decreased the nodule number, which suggests that GmCEP6 functions as a positive regulator in soybean nodulation. qRT-PCR showed that alterations in the expression of GmCEP6 affected the expression of marker genes in the Nod factor signaling pathway. Lastly, the function of GmCEP6 in nitrate inhibition of nodulation was analyzed; nodule numbers in the GmCEP6-overexpressed roots significantly increased under nitrogen treatments, which suggests that GmCEP6 functions in the resistance to nitrate inhibition. The study helps us understand that GmCEP6 promotes nodulation and participates in the regulation of nitrate inhibition of nodulation, which is of great significance for high efficiency utilization of nitrogen in soybeans.
1. Introduction
Nitrogen is one of the essential macroelements for plant growth, development, yield, and quality formation [1]. Therefore, improving nitrogen utilization efficiency is an important guarantee for high and stable soybean yields [2]. Legumes can not only absorb nitrogen from nitrogen-containing compounds such as ammonium and nitrate contained in the soil but also provide nitrogen by reducing free nitrogen to ammonia through symbiotic nitrogen fixation with rhizobia. The soybean, as an important symbiotic nitrogen fixation food crop, needs the rhizobia-soybean symbiotic system to fix the 50~90% nitrogen nutrition required for its growth [3]. Previous studies have shown that applying an appropriate amount of nitrogen fertilizer before sowing soybean can promote root nodule primordium formation and nodule organogenesis, improve the growth performance of rhizobia, promote plant growth, and provide effective carbon sinks and energy sinks for symbiotic nitrogen fixation [4]. Therefore, the symbiotic nitrogen fixation system between legume crops and rhizobia plays a very important role in nitrogen cycling.
Nitrogen uptake by plants from the soil is mainly in the form of nitrate [5]. Nitrate, however, tends to be unevenly distributed in soils. Thus, plants have evolved a systematic long-distance signaling pathway (CEP-CEPR module) for compensatory nitrate uptake in a N-starvation side of the root system [6]. The CEP polypeptide is one of the largest peptide signal groups secreted by plants; biochemical and functional evidence suggests that 15 amino acid peptides derived from the C-terminal region of precursor peptides act as ligands to regulate various stages of plant growth and development. Although CEP peptides have long been known to play a role in local cell-to-cell communication within specific tissues, recent advances indicate their new role as long-distance mobile signals required for systemic nutritional responses [7,8].
The Arabidopsis genome contains 15 CEP genes [9], of which seven are upregulated about 10 times in response to N starvation [10]. They are expressed specifically in the stele of lateral roots and are loaded into xylem vessels for transportation to the shoots [11]. The CEP family peptides are then recognized by receptor kinase CEP receptor 1 (CEPR1), which is expressed in leaf vascular tissue and induces the production of shoot secondary signals that up-regulate nitrate transport genes, such as NRT2.1, at the distal end of the root to compensate for local N starvation [6]. Because CEP family peptides and CEPR1 are widely presented in seed plants, the CEP-CEPR signaling module appears to be evolutionarily conserved [6,8]. In Medicago truncatula, MtCEP1 is the homologous of AtCEP9, but there are two CEP domains in MtCEP1, which are mainly expressed in the root tip, root vascular tissue, and lateral root meristem and are induced by different levels of nitrogen treatments [11]. However, the developmental role of CEP polypeptides in soybeans is not clear.
Here, 22 CEP family members were identified in soybeans through systematic bioinformatic study. We found that the expression of GmCEP6 was higher in roots and nodules, and histochemical staining was applied to validate this result. The effect of GmCEP6 on nodule development was evaluated, and we found that GmCEP6 functioned as a positive regulator and was partially tolerant to nitrate inhibition of nodulation. The results of this study may be utilized for high-nitrogen-fixation-efficiency soybean breeding in future.
2. Materials and Methods
2.1. Identification and Bioinformatic Analysis of GmCEP Genes
AtCEPs (Arabidopsis thaliana CEP genes family, AtCEPs) protein sequences were obtained from the Arabidopsis Information Resource database (https://www.arabidopsis.org/, (accessed on 2 January 2022)) [12]. The genome sequence, gff3 file, and protein sequence of soybean (Glycine max) were downloaded from the Ensembl database (http://plants.ensembl.org/index.html, (accessed on 4 January 2022)) [13]. The Blast wrapper tool in the bioinformatic analysis software TBtools v2.056 was applied to retrieve the GmCEPs based on the AtCEP protein sequence. After removing the duplicates of the GmCEP sequences, the amino acid sequences of remaining GmCEPs were submitted to the InterPro database (https://www.ebi.ac.uk/interpro/, (accessed on 14 January 2022)) [14] for protein domain prediction. Conserved CEP (C-terminal Encoded peptide) domains containing GmCEPs were screened for further analysis. The ProtParam (https://web.expasy.org/protparam/, (accessed on 11 February 2022)) [15] database was used for the physical and chemical properties analysis, including the number of amino acids, theoretical isoelectric point (pI), and relative molecular mass. The online website MEME (https://meme-suite.org/meme/, (accessed on 17 February 2022)) [16] was used for characteristic analysis of the GmCEPs motifs. The Muscle program of MEGA-X was applied to construct the CEPs’ phylogenetic tree; the NJ (neighbor-joining) [17] adjacency method was used to analyze the evolution distance.
2.2. Plant Materials and Growth Conditions
Soybean (G. max L. cv. Williams 82) seeds (kindly provided by Professor Xia Li from Huazhong agriculture university for research only) were surface-sterilized in 95% alcohol for 1 min and in 5% NaClO for 5 min, then washed several times using ddH2O water. The basic nutrient solution was referred to in a previous publication [18]. KNO3 was selected as a nitrogen source to set different nitrate concentrations, with no nitrogen (0 N, 0 mM), low nitrogen (LN, 4 mM), and high nitrogen (HN, 16 mM). Soyabeans were planted in 12 cm square boxes with a common vermiculite substrate. One soyabean plant was planted in each box and cultured in a growth house with photoperiod cycle (light/dark: 14 h/10 h) at 25 °C cultivation temperature, under the light intensity 10,000 lx and 70%. On the 10th day after seeding, the seedlings developed into the cotyledon stage and were inoculated with rhizobium bacteria, and the roots were harvested on the 7th day after seeding. After inoculation with rhizobia, HAI was the duration of different short-term treatments within 24 h, and DPI was the number of days of treatment. Agrobacterium rhizogenes strain K599 was used for the hairy-root transformation. The hairy-root transformation procedure was as previously described, with some modifications [14]. Soybeans sown with common vermiculite for 3 days were selected for hairy-root transformation and were inoculated with rhizobia on the 7th day after planting, and the roots were harvested on the 14th and 28th day after treatment, respectively. During the whole plant-growth period, nitrogen-free nutrient solution and water were required to be irrigated in rotation. The 14th and 28th days after inoculating the soybean seedlings with rhizobium were two important early nodulation development periods. For the nodulation assay, the plants were inoculated with a suspension of B. japonicum strain USDA110 (30 mL, OD600 = 0.08).
2.3. Vector Construction
For the GmCEP6 promoter, a GUS reporter fusion construct, 2369 bp upstream ATG of GmCEP6 region, was selected and amplified from cv. Williams 82 genomic DNA and cloned into pMDC162 though a gateway system. The GmCEP6 full-length coding sequence was cloned into pB7RWG2.0 using the same strategy for the overexpression construction. For the CRISPR-Cas9 construction, the top two reliable sgRNAs (small guide RNA, sgRNA), CATGAACTACTCGGTAGTGAGGG and CCGTAGCATTAGAAGCCT AGGG were selected. Then, vector pCBC-DT1T2 was used as a template to clone the two CRISPR fragments, and the two obtained products were inserted into vector pKSE401-GFP.
2.4. RNA Extraction and Expression Analysis
RNAprep Pure Plant plus Trizol Kit was used to extract RNA from collected transgenic hairy roots, soybean leaves, roots, and nodules, and the first-strand cDNA was synthesized using a super Mix Kit (Hifair II 1 strand cDNA Synthesis SuperMix, gDNA digester plus) (Yeasen Biotech Co. Ltd., Shanghai, China). qPCR was performed using SYBR Green JumpStart Taq ReadyMix (Sigma-Aldrich, St. Louis, MO, USA). GmCYP2 was used as an internal control [19]. (The primers used in this study are shown in Table S1).
2.5. Histochemical Analysis of GmCEP6 Expression
Composite transgenic roots expressing GmCEP6pro:GUS were inoculated with B. japonicum strain USDA110. The transformed hairy roots at different infection and nodulation stages were stained with X-Gluc at 37 °C for 8 h to test for β-glucuronidase activity. GUS activity was observed with a light microscope (OLYMPUS U-TV0.5XC-3).
2.6. Statistical Analysis
One-way analysis of variance (ANOVA) and Student’s t-test were used to performed p values. The gene expression and nodule numbers were analyzed using IBM SPSS 22.0 and GraphPad Prism 5 (GraphPad Software). The data are means with ±SE (Standard Error) from three independent replicates. The statistical differences are marked as follows: * p < 0.05; ** p < 0.01; *** p < 0.001.
3. Results
3.1. Identification and Physicochemical Properties of the Soybean CEP Family Gene
22 soybean CEP gene family members were obtained from the soybean genome by using BLAST and HMMER search and were named GmCEP1-22 according to their chromosomal positions; the amino acids residues encoded by them ranged from 80 (GmCEP6) to 163 (GmCEP20), and the molecular weights of the 22 GmCEPs ranged from 8740.03 (GmCEP6) to 17,533.65 (GmCEP20) Da. Theoretical isoelectric points of 22 GmCEPs family members ranged from 6.26 (GmCEP22) to 10.60 (GmCEP19) and belonged to alkaline proteins (Table 1). Six members of the GmCEPs family contained two CEP motifs, while 16 contained one. Conserved domains analysis of GmCEP family members showed that Motif 1 and Motif 2 were present in all CEP proteins. Motif 5 is the second motif in the GmCEP family, with 16 CEP proteins containing this motif. The least contained motif is Motif 9, with only two CEP proteins containing this motif (Table 1; Figure S1). We found there were no introns in all 22 GmCEP gene family members, according to the gene structure analysis (Figure S2).
3.2. Phylogenetic Analysis of CEPs in Soybeans
In order to study the evolutionary relationships of soybeans with other plants, 22 soybean CEP amino acids sequences were aligned with 15 Arabidopsis thaliana (At) AtCEP proteins, 11 Medicago truncatula (Mt) MtCEP proteins, and 15 Oryza sativa (Os) OsCEP sequences, collected for analysis. As shown in Figure 1, the CEP family members of the four species were divided into three subfamilies (I, II, and III). There were 24 gene family members in group I that contained 14 rice CEP family members. The remaining family members were from Arabidopsis (five), soybean (four), and one from M. truncatula. Except AtCEP16, the CEP family members in Group II were from soybean and M. truncatula, which suggests it may be a dicots- or even legume-specific group CEP. Ten Arabidopsis and eleven soybean CEP family members were distributed in Group III.
3.3. Digital Expression Pattern of CEPs in Soybeans
A dynamic expression Heatmap was constructed to dissect the soybean symbiotic related GmCEP genes; the online transcriptome data of nine soybean tissues covered most organs, and the developmental stages were analyzed. As shown in Figure 2, the results show that CEP genes had diverse expression patterns in different developmental stages of organs and tissues; for instance, most GmCEPs had similar expression levels in various tissues, while GmCEP22, GmCEP15, and GmCEP5 had extremely low expression in other tissues, except for a higher expression in one or two special tissues, which suggests that they might play a special role in their corresponding biological processes. In particular, we found that GmCEP6 was the highest expressed gene in root nodules among the 22 soybean CEPs, indicating that it might be involved in soybean and rhizobia interaction. Nitrogen, including nitrate, ammonia, and urea, had a significant effect on rhizobium infection and nodule initiation; the expression changes of the above three nitrogen treatments of roots and leaves were compared to standard values. We found that GmCEP6 was the only gene which was dramatically affected by the three kinds of nitrogen treatment of both the leaf and root.
3.4. GmCEP6 Is Preferentially Expressed in Soybean Nodules
Bioinformatics analysis revealed that the full length transcript of GmCEP6 was 730 bp with an entire exon, the cDNA contained a 5′ untranslated region (UTR) of 101 nucleotides and 3′ UTR of 386 bp, and the gene contained a 243 bp open reading frame (ORF) (Figure 3A), encoding a predicted 80 amino acid residues proteins with conserved C-terminally encoded peptides (CEP) (66-80aa) (Figure 3B), similar to its homologs in other plant species. A comparative analysis of GmCEP6 transcript levels was performed. Firstly, the relative expression level of GmCEP6 in nodules, root, stem, and leaves was determined using qRT-PCR, as shown in Figure 3C; GmCEP6 was mainly expressed in roots and nodules, which indicates a possible role of GmCEP6 in nodulation (Figure 3C). To further check the GmCEP6 expression in response to rhizobium infection, soybean seedlings were inoculated with Bradyrhizobium japonicum USDA110, and the transcript abundance of GmCEP6 at different stages was confirmed, as shown in Figure 3D; GmCEP6 was weakly induced by USDA110 treatment and peaked at 16 DPI in infected roots, while dramatically decreased at 28 DPI. In addition, GmCEP6 was markedly higher expressed in nodules than in roots at the checked time points (Figure 3D). Finally, to visually determine the expression profile of GmCEP6 in soybean nodulation, transgenic hairy roots harboring the -2kb promoter region upstream of GmCEP6 ATG were fused to the β-glucuronidase (GUS) reporter (pCEP6:GUS). Histochemical GUS staining was performed in transgenic hairy roots inoculated with rhizobia at 10 DPI. We found that GmCEP6 was mainly expressed in the pericycle, nodule primordium, lateral root primordium, and root nodule (Figure 3E–I). In addition, GUS signaling was mainly detected in the infection zone of mature nodules (28 DPI). These results suggest that GmCEP6 may play a vital role in soybean nodulation and nitrogen fixation.
3.5. GmCEP6 Plays a Key Role in Soybean Nodulation
To determine the roles of GmCEP6 in soybean nodulation, we generated transgenic hairy roots carrying GmCEP6 overexpressing (OE) or GmCEP6-CRISPR cas9 (KO). As shown in Figure 4, both GmCEP6 overexpressing (OE) and GmCEP6-CRISPR cas9 (KO) significantly affected soybean nodulation. In GmCEP6 overexpressing roots, qRT-PCR was applied to check the overexpression efficiency; the result showed that the transcript of GmCEP6 was about 20-fold in the GmCEP6-OE roots that in the control roots (Figure 4D). Then, the nodule numbers per root was quantified at 14 and 28 days after inoculation; we found, in GmCEP6 overexpressing hairy roots, the nodule number increased by 2.875 times (14 DPI) and four times (28 DPI), respectively. These data suggested that GmCEP6 plays a positive role in regulating soybean nodulation.
To further check this, the effect of GmCEP6 on soybean nodulation was evaluated in GmCEP6-CRISPR cas9 transgenic roots; gene editing and knock-out efficiency were validated by sequencing, and deletions and mutations can be detected in GmCEP6-KO roots (Figure S3). As shown in Figure 4E,F, in GmCEP6-KO root lines, nodule numbers at 14 and 28 days were decreased by 17 times and 44 times, respectively, compared with the control. Combined with the overexpression results, it is suggested that GmCEP6 is critical for the regulation of soybean nodulation.
3.6. GmCEP6 Affects the Expression of Related Genes in Nodulation Signal Pathway
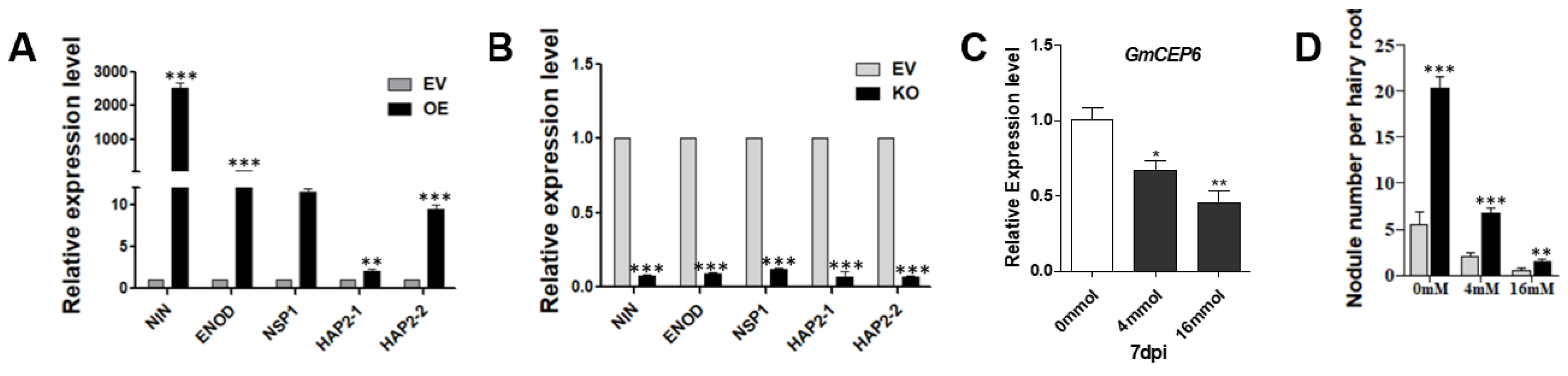
As soybean nodule numbers were significantly affected by GmCEP6 overexpression and GmCEP6 knock-out, we questioned whether GmCEP6 regulates soybean nodulation through the NF (Nodular Factor, NF) signaling pathway. To this end, we examined the expression pattern of several NF pathway marker genes in GmCEP6 overexpressed or knocked-out soybean roots. We selected GmENOD40 (Early nodulin), GmNINa (Nodule Inception), GmNSP1 (Nodulation Signaling Pathway 1), NF-YA1 (GmHAP2-1), and NF-YA2 (GmHAP2-2) to verify this [21,22,23,24]. As shown in Figure 5, the expression of GmNINa, GmENOD40, GmNSP1, GmHAP2-1, and GmHAP2-2 in GmCEP6-OE roots was significantly increased compared with that in empty vector control roots at 7 DPI. Meanwhile, we found the expression levels of these genes in GmCEP6-KO hairy roots were markedly reduced. These results suggest that GmCEP6 regulates soybean nodulation and nodule number controlling via modulating these symbiosis-related Nod factor signaling pathway genes.
Nodule number is regulated by an autoregulatory mechanism and by the nitrogen state of roots; a previous study on Medicago truncatula has shown that MtCEP1 increased nodulation and promoted nodule development at different nitrate concentrations. In order to investigate the response degree of CEP6 to nitrogen, we selected a medium concentration (4 mM) of nitrogen and high concentration (16 mM) of nitrate for treatment. It was found that CEP6 decreased with the increase of nitrogen concentration. However, we observed GmCEP6 overexpression in the hairy roots of 35s where CEP6 was treated with a high concentration of nitrate. Compared with WT, the number of overexpressed nodules was significantly higher. These results indicated that soybean nodulation is enhanced by overexpression of GmCEP6, and this tolerance to nitrogen inhibition for nodulation engaged by GmCEP6 could have beneficial outcomes in soybean breeding.
4. Discussion
CEP peptides play multiple roles in various plant biological processes. The first identified C-Terminally encoded secreted peptide AtCEP1 significantly arrests root growth [25]. The following reports proved that CEP genes responded to nitrogen deficiency, drought stress, and salt stress [26]. The CEP peptides were percepted by shoot expressed LRR-RLK CEPR, to mediate a systematic regulating of nitrogen deficiency [6]. Moreover, the CEP peptides and cytokinin converge on CEPD glutaredoxins to inhibit root growth through a local system [27]. The CEP peptides family number varied in different plants; there were 15 CEP peptides in A. thaliana, 11 in M. truncatula, 15 in O. sativa, 6 in C. sativus, and 21 in P. sativum. The function of several CEPs in the above plants have been identified. However, little is known about the CEP peptide family in soybeans. In this study, a systematic bioinformatics analysis was applied to identify soybean CEP peptides. A total of 22 GmCEPs were characterized from the soybean genome; the GmCEP proteins showed similar features to the previously discovered CEP family (Figure 1; Table 1). On analyzing the expression patterns of GmCEPs in the transcriptome, there were diverse expression patterns of GmCEPs in different developmental stages of organs and tissues, implying multiple roles of GmCEPs in regulating different biological processes in soybeans (Figure 2).
Legumes can specifically interact with their compatible rhizobia in the surrounding soil to form nodules. However, nodulation and nitrogen fixation in mature nodules is an high energy consumption process; thus, host legumes have evolved a root–shoot–root long-distance auto-regulation of nodulation (AON) system to refine the number of nodules [28,29]. NODULE INCEPTION (NIN) induced the expression of CLE ROOT SIGNAL1 (CLE-RS1) and CLE-RS2 to activate AON [30]. Another phenomenon in legume nodulation is sensitivity to soil nitrogen content; interestingly, recent studies have shown plants to also have a long-distance system (CEP-CEPR) in the nitrogen assimilation signaling pathway [6]. In legumes, the key transcriptional factor NIN coordinates CEP and CLE signaling peptides, combining these two long-distance signaling pathways to balance nitrogen absorption and symbiotic nitrogen fixation in order to meet high nitrogen demands [31]. In this study, another symbiosis-related CEP gene was characterized; we first identified a nodulation-related GmCEP6 as regulating soybean nodulation (Figure 3 and Figure 4) and as sharing a phenotype with reported legume CEP overexpression; the nodule number of its overexpression being close to the super-noduling phenotype of the NARK mutant in soybeans [18,30]. Moreover, MtCEP1 promoted MtNRT2.1 expression and nodulation dependent on compact root architecture 2 (MtCRA2) in low nitrate conditions [32]; MtCEP1/MtCRA2 balances root and nodule development by reducing auxin and ethylene responses [33]. Another study reports that MtCEP1, 2, and 12 redundantly regulate lateral root number and nodulation. Further study is needed to clarify the function diversification of GmCEPs family members, to construct the relationship between cytokinin and GmCEP, and to determine the relationship between the AON shoot-center component and CEPR [34]. We also need to clarify the roles of carbon signals in balancing the AON pathway and CEP-CEPR pathways that regulate nodule numbers.
5. Conclusions
In this study, a comprehensive analysis of the GmCEP genes was conducted at the whole-genome level; a total of 22 members in the CEP gene family were identified, and the structural features and evolutionary relationships of the GmCEPs were systematically analyzed. Further expression pattern analysis found that GmCEP6 was mainly expressed in nodules and showed significant responses to nitrate treatment and rhizobial infection, indicating its involvement in nodulation-related processes. Both overexpression and CRISPR-Cas9 were used to validate its role in nodulation. The results demonstrated that overexpression of GmCEP6 significantly increased nodule numbers, while knock-out of GmCEP6 led to a significant decrease, suggesting that GmCEP6 acts as a positive regulator in soybean nodulation. qRT-PCR results showed that changes in GmCEP6 expression positively influenced the expression of marker genes in the Nod factor signaling pathway. Finally, an analysis of the role of GmCEP6 in nitrate inhibition of nodulation revealed that, under nitrogen treatment, overexpression of GmCEP6 significantly increased nodule numbers, indicating its involvement in resistance to nitrate inhibition.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy14050988/s1, Figure S1: Motifs analysis of GmCEPs. Each block represented the position and strength of a motif. The blocks of GmCEPs motif were predicted using MEME. The motif sequence were listed in lower panel; Figure S2: Gene structure of 22 GmCEP genes. The gene structure information of GmCEPs were obtained through gff file of soybean, the diagram was constructed by Tbtools software; Figure S3: Validation of the mutation of GmCEP6-edited hairy roots. (A) Sequence of a region of soybean GmCEP6 with two target sites indicated. (B) Alignment of sequences of target-1 mutated alleles identified from cloned PCR fragments from crispr cas9 GmCEP6 (KO) transgenic root lines. Highlighting blue denotes the degree of homology of the aligned fragments, and only aligned regions of interest are displayed. Each trait represents a different mutation type. The most mutation was a base shift, represented by a green triangle, with a total of 5 (n = 13); Table S1: Primer sequences used in this study.
Author Contributions
L.W., W.D. and J.Z., conceived the project; X.W., J.Q., W.T. and M.W. performed the most experiments; X.W., A.Y. and L.W. prepared the original draft; S.W. supplemented the experimental data; L.W. and W.D. reviewed and finalized the manuscript, L.W. and W.D. supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Central Guidance for Local Science and Technology Development Projects (grant number YDZJSX2022A035), Technological Innovation 2030 Major Project (grant number 2023ZD040350404), Natural Science Foundation of Henan Province (Grant No. 202300410056), Basic research program of Shanxi Province (20210302123365, 202103021224146), National Natural Science Foundation of China (Grant No. 32241046), National Key Research and Development Program (Grant No. 2021YFD1600605-10), Shanxi Breeding Innovation Joint research and development projects (Grant No.2023xczx03), and National Laboratory of Minor Crops Germplasm Innovation and Molecular Breeding (in preparation) (Grant No. 202105D121010-23, 202204010910001-33).
Data Availability Statement
The RNA-seq data used in this study were download from the Phytozome database (https://phytozome-next.jgi.doe.gov/, (accessed on 1 March 2022)). Sequence data from this article can be found in the GenBank/EMBL or Glycine max Wm82.a4.v1 database, with the following entry number shown in Table 1. AtCEPs protein sequences were obtained from the Arabidopsis Information Resource database (https://www.arabidopsis.org/, (accessed on 6 March 2022)). The genome sequence, gff3 file, and protein sequence of soybean (Glycine max) were downloaded from the Ensembl database (http://plants.ensembl.org/index.html, (accessed on 9 March 2022)). The Muscle program of MEGA-X was applied to construct the CEPs’ phylogenetic tree; the NJ (neighbor-joining) adjacency method was used to analyze the evolution distance. The data that support the findings of this study are available from the corresponding author, L.W., upon reasonable request.
Acknowledgments
We thank Xia Li and Zhaosheng Kong for technical support and valuable suggestions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Kim, D.G.; Isaac, M.E. Nitrogen dynamics in agroforestry systems. A review. Agron. Sustain. Dev. 2022, 42, 60. [Google Scholar] [CrossRef]
- Terpolilli, J.J.; Hood, G.A.; Poole, P.S. Chapter 5. What Determines the Efficiency of N2-Fixing Rhizobium-Legume Symbioses. Adv. Microb. Physiol. 2012, 60, 325–389. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef]
- Murray, J.D.; Liu, C.-W.; Chen, Y.; Miller, A.J. Nitrogen sensing in legumes. J. Exp. Bot. 2016, 68, 1919–1926. [Google Scholar] [CrossRef]
- Tabata, R.; Sumida, K.; Yoshii, T.; Ohyama, K.; Shinohara, H.; Matsubayashi, Y. Perception of root-derived peptides by shoot LRR-RKs mediates systemic N-demand signaling. Science 2014, 346, 343–346. [Google Scholar] [CrossRef]
- Taleski, M.; Imin, N.; Djordjevic, M.A. CEP peptide hormones: Key players in orchestrating nitrogen-demand signalling, root nodulation, and lateral root development. J. Exp. Bot. 2018, 69, 1829–1836. [Google Scholar] [CrossRef]
- Roberts, I.; Smith, S.; Stes, E.; De Rybel, B.; Staes, A.; van de Cotte, B.; Njo, M.F.; Dedeyne, L.; Demol, H.; Lavenus, J.; et al. CEP5 and XIP1/CEPR1 regulate lateral root initiation in Arabidopsis. J. Exp. Bot. 2016, 67, 4889–4899. [Google Scholar] [CrossRef]
- Ogilvie, H.A.; Imin, N.; Djordjevic, M.A. Diversification of the C-TERMINALLY ENCODED PEPTIDE (CEP) gene family in angiosperms, and evolution of plant-family specific CEP genes. BMC Genom. 2014, 15, 870. [Google Scholar] [CrossRef]
- Roberts, I.; Smith, S.; De Rybel, B.; Broeke, J.V.D.; Smet, W.; De Cokere, S.; Mispelaere, M.; De Smet, I.; Beeckman, T. The CEP family in land plants: Evolutionary analyses, expression studies, and role in Arabidopsis shoot development. J. Exp. Bot. 2013, 64, 5371–5381. [Google Scholar] [CrossRef]
- Zhu, F.; Ye, Q.; Chen, H.; Dong, J.; Wang, T. Multigene editing reveals that MtCEP1/2/12 redundantly regulate lateral root and nodule number in Medicago truncatula. J. Exp. Bot. 2021, 72, 3661–3676. [Google Scholar] [CrossRef]
- Huala, E.; Dickerman, A.W.; Garcia-Hernandez, M.; Weems, D.; Reiser, L.; LaFond, F. The Arabidopsis Information Resource (TAIR): A comprehensive database and web-based information retrieval, analysis, and visualization system for a model plant. Nucleic Acids Res. 2001, 29, 102–105. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2018, 47, D351–D360. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, w202–w208. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Imin, N.; Mohd-Radzman, N.A.; Ogilvie, H.A.; Djordjevic, M.A. The peptide-encoding CEP1 gene modulates lateral root and nodule numbers in Medicago truncatula. J. Exp. Bot. 2013, 64, 5395–5409. [Google Scholar] [CrossRef]
- Jian, B.; Liu, B.; Bi, Y.; Hou, W.; Wu, C.; Han, T. Validation of internal control for gene expression study in soybean by quantitative real-time PCR. BMC Mol. Biol. 2008, 9, 59. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Franssen, H.; Yang, W.C.; Katinakis, P.; Bisseling, T. Characterization of GmENOD40, a Gene Expressed in Soybean Nodule Primordia; Springer: Amsterdam, The Netherlands, 1993. [Google Scholar] [CrossRef]
- Wang, L.; Sun, Z.; Su, C.; Wang, Y.; Yan, Q.; Chen, J.; Ott, T.; Li, X. A GmNINa-miR172c-NNC1 Regulatory Network Coordinates the Nodulation and Autoregulation of Nodulation Pathways in Soybean. Mol. Plant 2019, 12, 1211–1226. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, L.; Wang, Y.; Li, X. The NMN Module Conducts Nodule Number Orchestra. iScience 2020, 23, 100825. [Google Scholar] [CrossRef]
- He, C.; Gao, H.; Wang, H.; Guo, Y.; He, M.; Peng, Y.; Wang, X. GSK3-mediated stress signaling inhibits legume-rhizobium symbiosis by phosphorylating GmNSP1 in soybean. Mol. Plant 2021, 14, 488–502. [Google Scholar] [CrossRef]
- Ohyama, K.; Ogawa, M.; Matsubayashi, Y. Identification of a biologically active, small, secreted peptide in Arabidopsis by in silico gene screening, followed by LC-MS-based structure analysis. Plant J. 2010, 55, 152–160. [Google Scholar] [CrossRef]
- Aggarwal, S.; Kumar, A.; Jain, M.; Sudan, J.; Singh, K.; Kumari, S.; Mustafiz, A. C-terminally encoded peptides (CEPs) are potential mediators of abiotic stress response in plants. Physiol. Mol. Biol. Plants 2020, 26, 2019–2033. [Google Scholar] [CrossRef]
- Taleski, M.; Chapman, K.; Novák, O.; Schmülling, T.; Frank, M.; Djordjevic, M.A. CEP peptide and cytokinin pathways converge on CEPD glutaredoxins to inhibit root growth. Nat. Commun. 2023, 14, 1683. [Google Scholar] [CrossRef]
- Krusell, L.; Madsen, L.H.; Sato, S.; Aubert, G.; Genua, A.; Szczyglowski, K.; Duc, G.; Kaneko, T.; Tabata, S.; de Bruijn, F.; et al. Shoot control of root development and nodulation is mediated by a receptor-like kinase. Nature 2002, 420, 422–426. [Google Scholar] [CrossRef]
- Okamoto, S.; Kawaguchi, M. Shoot HAR1 mediates nitrate inhibition of nodulation in Lotus japonicus. Plant Signal. Behav. 2015, 10, e1000138. [Google Scholar] [CrossRef]
- Soyano, T.; Hirakawa, H.; Sato, S.; Hayashi, M.; Kawaguchi, M. NODULE INCEPTION creates a long-distance negative feedback loop involved in homeostatic regulation of nodule organ production. Proc. Natl. Acad. Sci. USA 2014, 111, 14607–14612. [Google Scholar] [CrossRef]
- Laffont, C.; Ivanovici, A.; Gautrat, P.; Brault, M.; Djordjevic, M.A.; Frugier, F. The NIN transcription factor coordinates CEP and CLE signaling peptides that regulate nodulation antagonistically. Nat. Commun. 2020, 11, 3167. [Google Scholar] [CrossRef]
- Luo, Z.; Wang, J.; Li, F.; Lu, Y.; Fang, Z.; Fu, M.; Mysore, K.S.; Wen, J.; Gong, J.; Murray, J.D.; et al. The small peptide CEP1 and the NIN-like protein NLP1 regulate NRT2.1 to mediate root nodule formation across nitrate concentrations. Plant Cell 2023, 35, 776–794. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Deng, J.; Chen, H.; Liu, P.; Zheng, L.; Ye, Q.; Li, R.; Brault, M.; Wen, J.; Frugier, F.; et al. A CEP Peptide Receptor-Like Kinase Regulates Auxin Biosynthesis and Ethylene Signaling to Coordinate Root Growth and Symbiotic Nodulation in Medicago truncatula. Plant Cell 2020, 32, 2855–2877. [Google Scholar] [CrossRef] [PubMed]
- Magori, S.; Kawaguchi, M. Long-distance control of nodulation: Molecules and models. Mol. Cells 2009, 27, 129–134. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Phylogenetic analysis of CEPs in G. max (Gm), O. sativa (Os), A. thaliana (At), and M. truncatula (Mt). The amino acid sequences of GmCEPs, OsCEPs, AtCEPs, and MtCEPs were downloaded and submitted into MEGA-X v10.0.0 software for alignment and phylogenetic tree construction; the phylogenetic tree was constructed using the NJ (neighbor-joining) adjacency method with 1000 bootstrap replicates.
Figure 1.
Phylogenetic analysis of CEPs in G. max (Gm), O. sativa (Os), A. thaliana (At), and M. truncatula (Mt). The amino acid sequences of GmCEPs, OsCEPs, AtCEPs, and MtCEPs were downloaded and submitted into MEGA-X v10.0.0 software for alignment and phylogenetic tree construction; the phylogenetic tree was constructed using the NJ (neighbor-joining) adjacency method with 1000 bootstrap replicates.
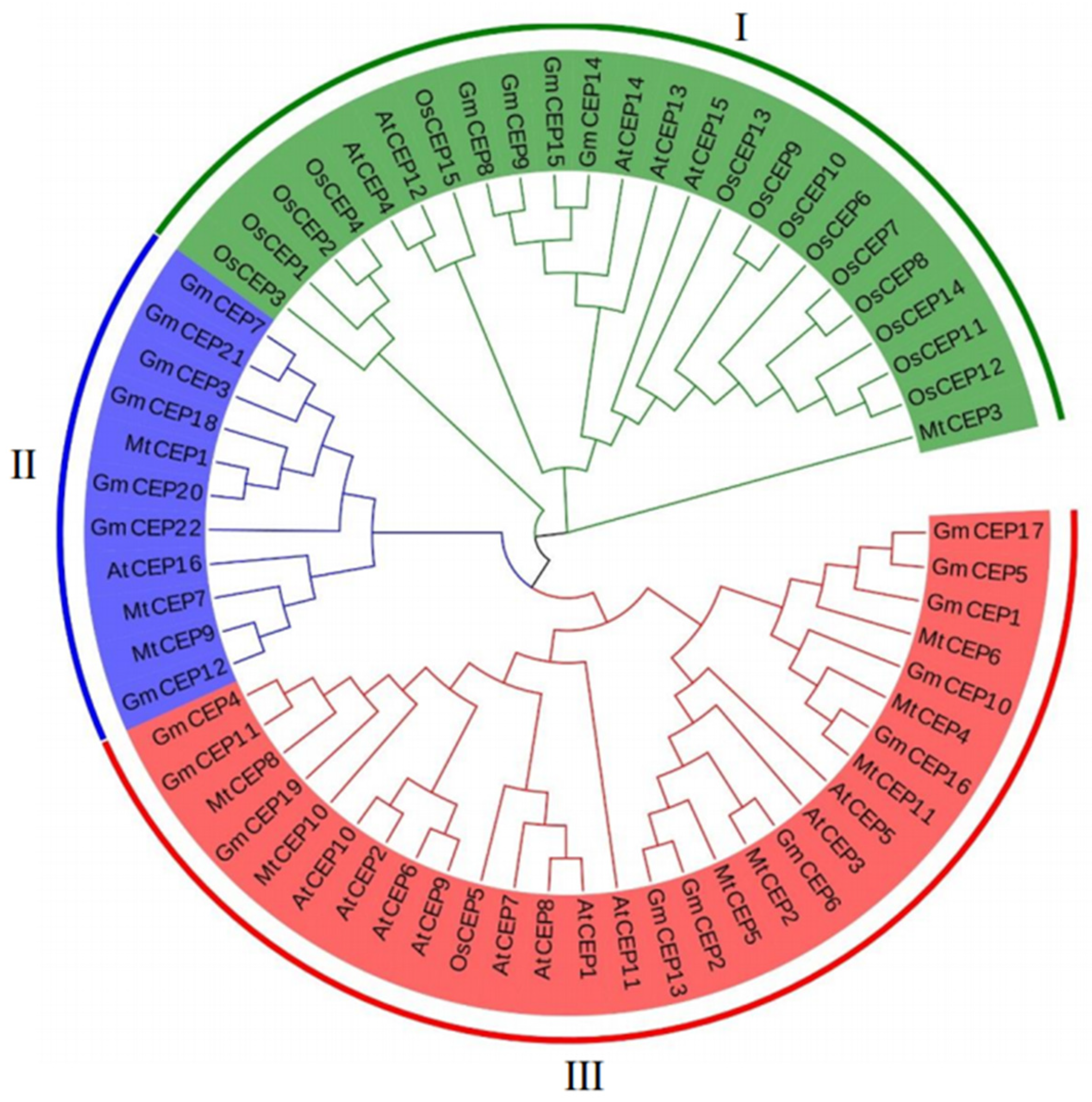
Figure 2.
The GmCEP expression profiles in different organs with or without treatments. The transcriptome data of GmCEP genes in nine tissues (root, stem, bud, leaf, flower, nodule, root tip, pod, and seed) and three different treatment development stages (ammonia treatment, nitrogen treatment, and urea treatment of root and leaf) were obtained from the phytozome database. The Heatmap package in Tbtools [20] software was applied to show the expressed FPKM value in different tissues.
Figure 2.
The GmCEP expression profiles in different organs with or without treatments. The transcriptome data of GmCEP genes in nine tissues (root, stem, bud, leaf, flower, nodule, root tip, pod, and seed) and three different treatment development stages (ammonia treatment, nitrogen treatment, and urea treatment of root and leaf) were obtained from the phytozome database. The Heatmap package in Tbtools [20] software was applied to show the expressed FPKM value in different tissues.

Figure 3.
Expression pattern of GmCEP6 in soybean nodulation. (A) Schematic gene structure of GmCEP6. (B) Domain analysis of GmCEP6. The conserved C-terminally encoded peptide (CEP) (66–80 aa) is shown in red. (C) Relative expression of GmCEP6 in soybean root, nodule, stem and leaf at 28 DPI. (D) Relative expression of GmCEP6 in inoculated soybean roots (0, 0.5 HAI and 1, 4, 16, 28 DPI) and 16, 28 DPI nodules. Gene expression level was normalized based on the expression of housekeeping gene GmCYP2. Error bar represents the mean of four biological replicates with ±SE (Standard Error); the different letters indicate significant differences, p < 0.05. Asterisks in (D) indicate significant difference within a p level in t-tests. ** p < 0.01; *** p < 0.001. (E–I) Histochemical analysis of GmCEP6 expression in transgenic composite soybean roots and nodules: root tip region (E), emerged pericycle (F), lateral root primordium (G), nodule primordium (H), and nodule (I). Scale bar in (E–I) = 1 cm.
Figure 3.
Expression pattern of GmCEP6 in soybean nodulation. (A) Schematic gene structure of GmCEP6. (B) Domain analysis of GmCEP6. The conserved C-terminally encoded peptide (CEP) (66–80 aa) is shown in red. (C) Relative expression of GmCEP6 in soybean root, nodule, stem and leaf at 28 DPI. (D) Relative expression of GmCEP6 in inoculated soybean roots (0, 0.5 HAI and 1, 4, 16, 28 DPI) and 16, 28 DPI nodules. Gene expression level was normalized based on the expression of housekeeping gene GmCYP2. Error bar represents the mean of four biological replicates with ±SE (Standard Error); the different letters indicate significant differences, p < 0.05. Asterisks in (D) indicate significant difference within a p level in t-tests. ** p < 0.01; *** p < 0.001. (E–I) Histochemical analysis of GmCEP6 expression in transgenic composite soybean roots and nodules: root tip region (E), emerged pericycle (F), lateral root primordium (G), nodule primordium (H), and nodule (I). Scale bar in (E–I) = 1 cm.

Figure 4.
GmCEP6 regulates soybean symbiotic nodulation. (A) Soybean growth status of composite plants harboring empty vector, GmCEP6-OE, and GmCEP6-KO. (B) Nodulation performance of transgenic hairy root harboring empty vector, GmCEP6-OE, and GmCEP6-KO at 28 DPI. (C) Transgenic validation of hairy root harboring empty vector, GmCEP6-OE, and GmCEP6-KO using LUYOR-3415RG Hand-Held Lamp; GFP-positive roots were selected for further phenotype analysis. Scale bar in (B,C) = 1 cm. (D) Relative expression level of GmCEP6 in empty vector and GmCEP6-OE transgenic roots at 28 DPI were conducted to check the overexpression ratio; the expression value was normalized based on the expression of reference gene GmCYP2. (E) Quantitative data of nodule number per hairy root at 14 DPI. EV (empty vector for overexpressed transgenic roots) is the control. pKSE401 was the empty vector for CRISPR cas9 (KO). (F) Quantitative data of nodule number per hairy root at 28 DPI. EV: empty vector for overexpressed transgenic roots. pKSE401 was the empty vector for CRISPR cas9 (KO). These experiments were conducted on more than three dependent biological replicates. Data are means with ±SE (Standard Error) from three independent replicates (n = 12). Asterisks indicate significant difference within a p level in t-tests, ** p < 0.01; *** p < 0.001.
Figure 4.
GmCEP6 regulates soybean symbiotic nodulation. (A) Soybean growth status of composite plants harboring empty vector, GmCEP6-OE, and GmCEP6-KO. (B) Nodulation performance of transgenic hairy root harboring empty vector, GmCEP6-OE, and GmCEP6-KO at 28 DPI. (C) Transgenic validation of hairy root harboring empty vector, GmCEP6-OE, and GmCEP6-KO using LUYOR-3415RG Hand-Held Lamp; GFP-positive roots were selected for further phenotype analysis. Scale bar in (B,C) = 1 cm. (D) Relative expression level of GmCEP6 in empty vector and GmCEP6-OE transgenic roots at 28 DPI were conducted to check the overexpression ratio; the expression value was normalized based on the expression of reference gene GmCYP2. (E) Quantitative data of nodule number per hairy root at 14 DPI. EV (empty vector for overexpressed transgenic roots) is the control. pKSE401 was the empty vector for CRISPR cas9 (KO). (F) Quantitative data of nodule number per hairy root at 28 DPI. EV: empty vector for overexpressed transgenic roots. pKSE401 was the empty vector for CRISPR cas9 (KO). These experiments were conducted on more than three dependent biological replicates. Data are means with ±SE (Standard Error) from three independent replicates (n = 12). Asterisks indicate significant difference within a p level in t-tests, ** p < 0.01; *** p < 0.001.

Figure 5.
GmCEP6 regulates Nod factor signaling pathway genes and response to nitrate. (A,B) GmCEP6 regulates soybean symbiotic nodulation through Nod factor signaling pathway genes. (A) qRT-PCR analysis of the expression of GmNINa, GmENOD40, GmNSP1, GmHAP2-1, and GmHAP2-2 in roots carrying EV and GmCEP6-OE at 2 DAI (n = 10). (B) qRT-PCR analysis of GmNINa, GmENOD40, GmNSP1, and GmHAP2-2 in roots harboring pKSE401 and GmCEP6-KO at 28 DAI (n = 10). We set all of the transcript profiles of these genes in EV hairy roots at 28 DAI as “1”. The transcript amounts in each sample were normalized to the expression of reference gene GmCYP2. The expression levels are means ±SE. Asterisks indicate significant difference within a P level in t-tests. ** p < 0.01; *** p < 0.001. (C) qRT-PCR analysis of the expression of GmCEP6 in different nitrogen concentrations. (D) Quantification of nodule number at 28 DAI under different nitrate concentrations. Asterisks indicate significant difference within a P level in t-tests. * p < 0.05; ** p < 0.01; *** p < 0.001.
Figure 5.
GmCEP6 regulates Nod factor signaling pathway genes and response to nitrate. (A,B) GmCEP6 regulates soybean symbiotic nodulation through Nod factor signaling pathway genes. (A) qRT-PCR analysis of the expression of GmNINa, GmENOD40, GmNSP1, GmHAP2-1, and GmHAP2-2 in roots carrying EV and GmCEP6-OE at 2 DAI (n = 10). (B) qRT-PCR analysis of GmNINa, GmENOD40, GmNSP1, and GmHAP2-2 in roots harboring pKSE401 and GmCEP6-KO at 28 DAI (n = 10). We set all of the transcript profiles of these genes in EV hairy roots at 28 DAI as “1”. The transcript amounts in each sample were normalized to the expression of reference gene GmCYP2. The expression levels are means ±SE. Asterisks indicate significant difference within a P level in t-tests. ** p < 0.01; *** p < 0.001. (C) qRT-PCR analysis of the expression of GmCEP6 in different nitrogen concentrations. (D) Quantification of nodule number at 28 DAI under different nitrate concentrations. Asterisks indicate significant difference within a P level in t-tests. * p < 0.05; ** p < 0.01; *** p < 0.001.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of GmCEP family members.
| Gene Name | Gene ID | Chromosomal Location | Number of Amino Acids | Theoretical pI | Molecular Weight (Average) | CEP Motif Number |
|---|---|---|---|---|---|---|
| GmCEP1 | Glyma.01G184800 | 1 | 99 | 8.71 | 11,292.71 | 1 |
| GmCEP2 | Glyma.01G184900 | 1 | 87 | 9.82 | 9211.79 | 1 |
| GmCEP3 | Glyma.01G185000 | 1 | 150 | 8.74 | 16,377.5 | 2 |
| GmCEP4 | Glyma.01G185100 | 1 | 94 | 7.09 | 10,099.56 | 1 |
| GmCEP5 | Glyma.05G083900 | 5 | 86 | 7.8 | 9800.21 | 1 |
| GmCEP6 | Glyma.05G084000 | 5 | 80 | 10.24 | 8740.03 | 1 |
| GmCEP7 | Glyma.05G084100 | 5 | 156 | 9.3 | 16,969.18 | 2 |
| GmCEP8 | Glyma.05G161100 | 5 | 96 | 10.21 | 10,600.58 | 1 |
| GmCEP9 | Glyma.08G118500 | 8 | 93 | 9.92 | 10,181.12 | 1 |
| GmCEP10 | Glyma.09G218000 | 9 | 85 | 9.1 | 9335.88 | 1 |
| GmCEP11 | Glyma.11G057100 | 11 | 87 | 8.03 | 9270.58 | 1 |
| GmCEP12 | Glyma.11G057200 | 11 | 148 | 9.34 | 15,859.04 | 2 |
| GmCEP13 | Glyma.11G057300 | 11 | 87 | 9.83 | 9277.83 | 1 |
| GmCEP14 | Glyma.13G226600 | 13 | 94 | 10.14 | 10,213.95 | 1 |
| GmCEP15 | Glyma.15G085800 | 15 | 88 | 9.78 | 9634.2 | 1 |
| GmCEP16 | Glyma.16G108400 | 16 | 82 | 9.3 | 9010.27 | 1 |
| GmCEP17 | Glyma.17G176500 | 17 | 86 | 9.14 | 9865.22 | 1 |
| GmCEP18 | Glyma.17G176800 | 17 | 152 | 8.89 | 16,346.37 | 2 |
| GmCEP19 | Glyma.17G176900 | 17 | 87 | 10.6 | 9658.01 | 1 |
| GmCEP20 | Glyma.17G177000 | 17 | 163 | 9.27 | 17,533.65 | 2 |
| GmCEP21 | Glyma.17G177300 | 17 | 158 | 8.63 | 17,110.26 | 2 |
| GmCEP22 | Glyma.17G244700 | 17 | 108 | 6.26 | 11,742.32 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wu, S.; Wang, X.; Qin, J.; Tian, W.; Wang, M.; Yue, A.; Wang, L.; Du, W.; Zhao, J. Soybean CEP6 Signaling Peptides Positively Regulate Nodulation. Agronomy 2024, 14, 988. https://doi.org/10.3390/agronomy14050988
AMA Style
Wu S, Wang X, Qin J, Tian W, Wang M, Yue A, Wang L, Du W, Zhao J. Soybean CEP6 Signaling Peptides Positively Regulate Nodulation. Agronomy. 2024; 14(5):988. https://doi.org/10.3390/agronomy14050988
Chicago/Turabian StyleWu, Shuai, Xiaoli Wang, Jie Qin, Wenqing Tian, Min Wang, Aiqin Yue, Lixiang Wang, Weijun Du, and Jinzhong Zhao. 2024. "Soybean CEP6 Signaling Peptides Positively Regulate Nodulation" Agronomy 14, no. 5: 988. https://doi.org/10.3390/agronomy14050988
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.