Functional Analysis of Polyphenol Oxidases by Antisense/Sense Technology


1
Suranaree University of Technology, 111 University Ave., Muang District, Nakhon Ratchasima 30000, Thailand
2
Department of Entomology, Louisiana State University, 402 Life Sciences Building, Louisiana State University, Baton Rouge, LA 70803, USA
*
Author to whom correspondence should be addressed.
Molecules 2007, 12(8), 1569-1595; https://doi.org/10.3390/12081569
Submission received: 30 May 2007
/
Revised: 19 July 2007
/
Accepted: 19 July 2007
/
Published: 27 July 2007
(This article belongs to the Special Issue Phenolics and Polyphenolics)
Abstract
:Polyphenol oxidases (PPOs) catalyze the oxidation of phenolics to quinones, the secondary reactions of which lead to oxidative browning and postharvest losses of many fruits and vegetables. PPOs are ubiquitous in angiosperms, are inducible by both biotic and abiotic stresses, and have been implicated in several physiological processes including plant defense against pathogens and insects, the Mehler reaction, photoreduction of molecular oxygen by PSI, regulation of plastidic oxygen levels, aurone biosynthesis and the phenylpropanoid pathway. Here we review experiments in which the roles of PPO in disease and insect resistance as well as in the Mehler reaction were investigated using transgenic tomato (Lycopersicon esculentum) plants with modified PPO expression levels (suppressed PPO and overexpressing PPO). These transgenic plants showed normal growth, development and reproduction under laboratory, growth chamber and greenhouse conditions. Antisense PPO expression dramatically increased susceptibility while PPO overexpression increased resistance of tomato plants to Pseudomonas syringae. Similarly, PPO-overexpressing transgenic plants showed an increase in resistance to various insects, including common cutworm (Spodoptera litura (F.)), cotton bollworm (Helicoverpa armigera (Hübner)) and beet army worm (Spodoptera exigua (Hübner)), whereas larvae feeding on plants with suppressed PPO activity had higher larval growth rates and consumed more foliage. Similar increases in weight gain, foliage consumption, and survival were also observed with Colorado potato beetles (Leptinotarsa decemlineata (Say)) feeding on antisense PPO transgenic tomatoes. The putative defensive mechanisms conferred by PPO and its interaction with other defense proteins are discussed. In addition, transgenic plants with suppressed PPO exhibited more favorable water relations and decreased photoinhibition compared to nontransformed controls and transgenic plants overexpressing PPO, suggesting that PPO may have a role in the development of plant water stress and potential for photoinhibition and photooxidative damage that may be unrelated to any effects on the Mehler reaction. These results substantiate the defensive role of PPO and suggest that manipulation of PPO activity in specific tissues has the potential to provide broad-spectrum resistance simultaneously to both disease and insect pests, however, effects of PPO on postharvest quality as well as water stress physiology should also be considered. In addition to the functional analysis of tomato PPO, the application of antisense/sense technology to decipher the functions of PPO in other plant species as well as for commercial uses are discussed.
Introduction
Plants are exposed to a great many abiotic and biotic stresses in their environments that can, together or separately, reduce plant fitness. Accordingly, plants possess a wide variety of defensive traits to reduce the fitness impacts of these stresses. Among these defense mechanisms, the secondary metabolites of plants (defined broadly here to include defense-related proteins and enzymes as well as low-molecular weight organic molecules) play a central role. Because the occurrence of biotic and abiotic stresses is highly variable in space and time, and because multiple stresses may impinge on plants simultaneously, it is evident that plant defenses and the expression of plant defenses must exhibit flexibility, to allow appropriate defensive phenotypes to be displayed by plants at all times. Two attributes of most secondary metabolites in particular enable such flexibility: first, most secondary metabolites have roles in defense against multiple stresses and, second, many secondary metabolites are inducible; that is, they are only expressed, or are expressed at higher levels, as a result of prior stress.
Polyphenol oxidases (PPOs) catalyze the O2-dependent oxidation of mono- and o-diphenols to o-diquinones, highly reactive intermediates, the secondary reactions of which are believed to be responsible for the oxidative browning that occurs as a consequence of plant senescence, wounding and pathogen infection [1]. PPOs have been known to biochemistry for a century and several hypotheses regarding the function of PPO have been proposed, including roles in the phenylpropanoid pathway [2], the Mehler reaction, electron cycling, oxygen regulation [3], flower petal coloration [4] and plant defense [5]. A defensive role for PPO has frequently been suggested due to the conspicuous appearance of PPO reaction products upon wounding, pathogen infection, or insect infestation, and due to the inducibility of PPO in response to various abiotic and biotic injuries or signaling molecules [6]. Here we review the functional analysis of PPOs particularly through the use of transgenic tomato (Lycopersicon esculentum) with PPO expression levels altered by antisense/sense technology. The focus is on our previous research on the role of PPO in Pseudomonas syringae resistance and during water stress as well as new data on the role of PPO in defense against Lepidopteran insects. Other aspects of PPO such as reaction mechanisms, structure, import and processing have been extensively reviewed [1a, 7].
Polyphenol oxidase (PPO) and PPO genes
PPOs are ubiquitous, nuclear-encoded, copper metalloproteins found in all angiosperms [7]. PPOs catalyze two distinct reactions using oxygen: the o-hydroxylation of monophenols to o-diphenols (cresolase, tyrosinase, or monophenol oxidase activity [EC 1.14.18.1]), and the dehydrogenation of o-dihydroxyphenols to o-diquinones (catecholase or diphenol oxygen oxidoreductase activity [EC 1.10.3.2]). Quinones are highly reactive electrophiles that can participate in two secondary, nonenzymatic reaction pathways: the covalent 1,4 addition of o-quinones to cellular nucleophiles, and the reversed disproportionation of quinones to semiquinone radicals. Semiquinone radicals produced as a result of the latter pathway can either add covalently to other molecules or carry out the single-electron reduction of molecular oxygen to the superoxide anion radical (O2-), which may then give rise to other reactive oxygen species (ROS [7]). Moreover, in some plant species, nonenzymatic cyclization to amine-chrome may occur following an intramolecular 1,4 addition (i.e. o-dopaquinone [8]). These reactions can lead to the formation of black or brown quinone adducts that impose losses in quality and nutritional value in many fresh and processed vegetable and fruit crops [1]. Since most PPOs are targeted to the thylakoid lumen, PPO activity is often latent until disruptive forces such as wounding, senescence, or attack by insects or pathogens release it from the thylakoid to interact with its mono and/or o-diphenolic substrates that were previously kept in the vacuole [7].
PPO genes and cDNAs have been isolated and characterized from a number of plant species (summarized in Table 1) including tomato, potato (Solanum tuberosum), tobacco (Nicotiana tabacum), hybrid poplar (Populus trichocarpa x Populus deltoides), trembling aspen (Populus tremuloides), sugarcane (Saccharum spp.), wheat (Triticum aestivum), pokeweed (Phytolacca americana), red clover (Trifolium pratense), pineapple (Ananas comosus), broad bean (Vicia faba), spinach (Spinacia oleracea), apple (Malus domesticus) and grape (Vitis vinifera [9]). In most plant species studied, multiple members of the PPO gene family have been found which often exhibit differential expression patterns during growth and development and in response to stresses [9g, 10, 6c, 9l, 9n]. In contrast, only a single PPO gene has been identified in grape, although other PPO gene members with low degrees of similarity to this identified gene may be present in the grape genome [9d]. PPOs from all plant species studied possess two conserved copper-binding domains, CuA and CuB, responsible for copper coordination and interaction with molecular oxygen and phenolic substrates, and most also contain a transit peptide targeted to thylakoid lumen [7]. Only the PPO from snapdragon (Antirrhinum majus) has a N-terminal sequence that does not share the features of such transit peptide, and is localized in the vacuole, where its substrates are kept [4,11].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant | Genomic/ cDNA | Tissue source | Number of genes | Transcript size (Kb) | Protein size (kDa) | Reference |
|---|---|---|---|---|---|---|
| Tomato | Genomic | NA 1 | 7 | 2.0 | 66-71 (57-62) 2 | [9c] |
| Potato | cDNA | Leaf/young tuber | 6 | 2.0 | 67-68 (57) | [9b, 9g] |
| Tobacco | cDNA | Stigma/style | 1 (≥2) 3 | 2.3 | 68 (58) | [9i] |
| Apple | Genomic/cDNA | Leaf/fruit peel/ young fruit | 2 (≥4) | 2.0-2.2 | 65-66 (55-57) | [9e, 65, 10b] |
| Japanese pear | Genomic | Leaf | 1 (≥2) | NA | 66 (56) | [16] |
| Broad bean | cDNA | Mature leaf | 3 | 2.2 | 68-70 (58-60) | [9a, 66] |
| Pineapple | cDNA | Blackheart flesh | 2 (≥4) | 2.1 | 69 (58) | [9l] |
| Sweet potato | Genomic | NA | 2 | 2.1 | 66 (56) | [67] |
| Grape | cDNA | Immature berry | 1 | 2.2 | 67 (40) | [68, 9d] |
| Apricot | cDNA | Immature fruit | 1 (≥2) | 2.2 | 67 (56) | [17c] |
| Plum (Nai) | cDNA | Leaf/pulp | 2 | 2.2 | 67-68 (57) | [69] |
| Hybrid poplar | cDNA | Leaf (systemically wounded) | 1 (2) | 2.1 | 64 (57) | [9j] |
| Trembling aspen | cDNA | Wounded leaf | 1 (2) | 2.1 | 65 (59) | [9k] |
| Pokeweed | cDNA | Suspension culture | 2 | 2.1-2.3 | 65 (54) | [9f] |
| Lettuce | cDNA | Young leaf | 1 | 2.1 | 68 (57) | [70] |
| Spinach | cDNA | Primary leaf/ etiolated cotyledon | 2 | 2.5 | 73 (62-64) | [71] |
| Banana | cDNA | Flesh and peel of small fruit | 4 (≥4) | 2.1 | 67 (56) | [12a, 70] |
| Sugarcane | cDNA | Immature stem | 1 (≥2) | 2.2 | 67 (59) | [9h] |
| Wheat | Genomic/ cDNA | Immature seed/ QTL/EST | 3 (≥6) | 2.0 | 57-68 (50-58) | [9m, 12b, 72] |
| Red clover | cDNA | Leaf | 3 (3-5) | 1.9 | 68-71 (57-60) | [9n] |
| Alfafa | Genomic | NA | 1 | NA | 69 (58) | [73] |
| Creosate bush | cDNA | Leaf | 1 | 1.8 | 66 (43) | [51] |
| Snapdragon | cDNA | Petal | 1 | 1.7 | 64 (39) | [4] |
| Tea | Genomic | NA | 3 | NA | 65-68 (54-57) | [74] |
- 1
- NA: not available.
- 2
- Predicted protein size (predicted mature protein size after N-terminal cleavage of a transit peptide and/or C-terminal cleavage).
- 3
- The number in parenthesis indicates expected number of genes in the genome estimated from Southern analysis.
The PPO gene family of tomato consists of seven members, PPO A, A’, B, C, D, E and F, clustered on chromosome 8. All seven genes lack introns, a feature that is common to most PPO genes, except for a banana PPO gene that appears to contain a 85-bp intron in its sequence and two wheat PPO genes that contain 2 introns [9c, 12]. Although these genes are highly conserved (70-100% identity in their coding regions), they exhibit spatially and temporally differential expression in vegetative and reproductive organs of tomato as well as differential inducibility in response to various biotic and abiotic agents [9c, 6c, 10a, 5d, 13]. In potato, multiple PPO cDNAs were isolated and characterized from leaves and tubers, indicating a gene family with specific temporal and spatial expression patterns of individual members, and encode polypeptides whose sequences are highly homologous to tomato PPO [9b, 9g]. A PPO cDNA isolated from tobacco, another Solanaceae species, also encodes a polypeptide with high similarity to both tomato and potato. This gene is a member of the multigene family and shows flower-specific pistil-predominant expression [9i]. Bucheli et al. [9h] and Stewart et al. [9l] showed that the PPO protein sequences of this Solanaceae family formed one group in the phylogenetic tree, while those of apple, bean, pineapple, sweet potato (Ipomoea batatas), grape, apricot (Prunus armeniaca), pokeweed and lettuce (Lactuca sativa) formed another distinct group. The spinach PPO appeared different from all the other C3 dicotyledonous sequences, and its expression was found to be light-regulated [14, 9h].
In the monocotyledonous C4 plant sugarcane, the sequence of PPO cDNA is significantly different from those of C3 dicotyledonous plants, although the size of the mature PPO protein (59 kDa) is within the range found in dicotyledonous species [9h]. Amino acid sequences of wheat PPO share more similarity to those of monocotyledonous sugarcane and pineapple [9m, 12b].
In fruit trees, PPOs of Rosaceae plants (apple, pear [Pyrus communis], Japanese pear [Pyrus pyrifolia], Chinese quince [Pseudocydonia sinensis], Japanese loquat [Eriobotrya japonica] and peach [Prunus persica]) are highly homologous in their core amino acid sequences (85.3-97.5%), and their immunological and enzymatical criteria [15]. The distribution of PPOs in apple and Japanese pear fruits appeared to be spatially different: mainly localized around the core for apple and throughout the fruit for Japanese pear [16]. The deduced amino acid sequence of trembling aspen PPO shows particularly significant identity with those from the woody plants (hybrid poplar, apple, apricot and grape).
Red clover PPO is most closely related with those of the legumes V. faba and alfafa (Medicago sativa; 68-72% similarity). The three red clover PPO genes differ in their spatial and temporal expression patterns of leaves, petioles, stems and flowers [9n].
Highest PPO expression levels are usually associated with young tissues (leaves, flowers, fruits, tubers) and in meristematic regions, which are particularly vulnerable to diseases and insect pests, and gene expression generally declines during development and maturation of plant tissues [9a, 17, 9b, 9d, 9e, 9g, 10a, 12a, 9n]. In contrast, the expression of pokeweed PPO specifically localized in ripened fruits that had turned red from betalain accumulation [9f]. In some plant species, PPOs are very stable throughout growth and development and often exist in a latent form in mature tissues when PPO transcripts are no longer found [9d, 9h, 12a].
Wound inducibility of PPO has been observed in many tissues of various plants. Potato and tomato PPOs were systemically and/or locally induced in response to wounding or pathogen infection, especially in young leaf tissues [17b, 6c]. Stewart et al. [9l] reported PPO induction in both leaves and fruits of pineapple following wounding. Chilling and blackheart symptoms also induced the increases in pineapple PPO transcript levels. In trembling aspen and hybrid poplar, leaf-age-dependent local and systemic induction of PPO expression was detected [9j, 9k]. An apple PPO cDNA is upregulated in wounded fruit and foliar tissues, and also in peel tissue showing the symptoms of superficial scald, a postharvest disorder [9e, 10b]. However, in banana peel and flesh, no significant increase in PPO activity was observed after a variety of wounding regimes; sliced, bashed, perforated and rolled [12a]. Other species such as apricot exhibited lack of wound-induced accumulation of foliar PPO transcripts [17c].
Antisense/sense transgenic tomato
Biochemical and physiological studies have provided few conclusive answers regarding the function of PPO. In addition, no PPO-null mutants have been identified; therefore, the use of genetically modified plants with altered PPO expression is an essential platform for testing various functional hypotheses in an otherwise identical genetic background [7]. Two types of transgenic tomato were generated by our group for this purpose: antisense PPO plants with suppressed PPO activity (SP) and sense PPO plants overexpressing PPO activity (OP). To generate these transgenic SP and OP tomatoes, transformation constructs were made by ligating a 2 kb potato PPO cDNA [9b], exhibiting 68-91% DNA sequence similarity with the seven tomato PPO genes [9c], into transformation vector pBI 121 (Clontech) in the antisense orientation relative to the CaMV 35S promoter, or ligating the same gene in the sense orientation relative to the CaMV 35S promoter within cloning vector pART 7 [18]. These SP and OP transgenic plants have been described in detail previously [5c, 5d]. Two representative SP transgenic lines, A14-6 and A19-3, were further used for the analysis. A14-6 possesses two linked T-DNA copies [19]. A19-3 also showed a single-locus inheritance of the antisense transgene. For OP plants, two transgenic lines, S-18 and S-28, both of them carrying at least two T-DNA copies [5c], were chosen for further experimentation. A14-6 and A19-3 have ca. 2-40 fold reduced PPO activity, and lack or have low levels of immunologically detectable PPO in leaf homogenates. Neither endogenous nor antisense PPO mRNA is detectable in leaves or flowers of A14-6, while S-18 and S-28 contain 2-10 fold increased PPO activity and immunodetectable PPO, and up to 30 fold increase in PPO transcripts [5c, 5d, 13, unpublished data].
The alteration of PPO activity had no effect on plant growth and development. Germination percentages, growth rates, total leaf area, shoot and root dry weights and numbers of seeds per fruit were not statistically different among SP, OP transgenic plants and nontransformed (NT) control plants. In addition, the transgenic plants exhibited similar morphology and plant vigor to NT plants and flowered and set seeds normally, implying that under the growth conditions of greenhouse, growth chamber and laboratory, PPO does not play a critical role in plant metabolism, growth and development [7, 5c, 5d, 13, unpublished data]. The PPO activity levels (measured in the presence of catalase to eliminate the contribution of peroxidase [PO] activity to phenolic oxidation) of these transgenic tomatoes as well as NT controls appeared to vary from experiment to experiment according to plant developmental stage and growth conditions. However, in all experiments, the relative ranking remained the same with S-18 and S-28 having the highest PPO activity, NT having moderate PPO activity and A14-6 and A19-3 having the lowest PPO activity [5c, 5d, 13, Mahanil et al., unpublished manuscript, Bhonwong et al., unpublished manuscript]. However, no changes in PO activity, total protein contents and p-coumaric acid and caffeic acid pools was observed in the foliage of transgenic tomatoes with modified PPO activity levels compared to NT control [5c, 5d, unpublished data].
Role of PPO in disease resistance
PPO has been frequently implicated in resistance to diverse pathogens since quinones, its primary products, are highly reactive electrophiles that can undergo complex secondary reaction pathways, in particular the 1,4 addition of o-quinones to cellular nucleophiles and the reversed disproportionation of quinones to semiquinone radicals that may lead to generation of reactive oxygen species (ROS).
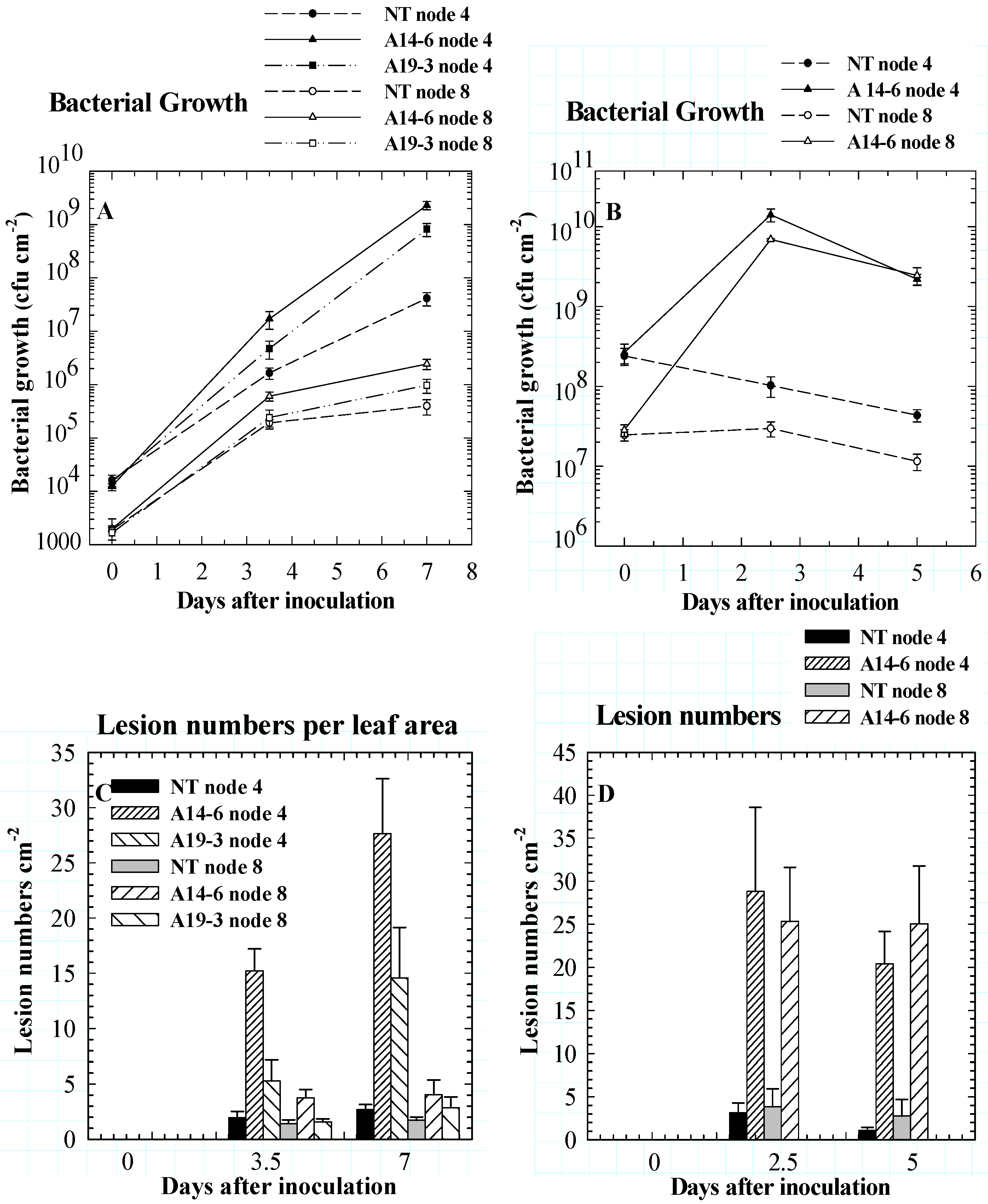
Figure 1.
In both compatible interactions (A, C) and incompatible interactions (B, D) between tomato and Pseudomonas syringae, PPO suppression results in enhanced susceptibility to infection. Antisense PPO transgenic plants have higher bacterial growth (A, B) and lesion numbers (C, D) than nontransformed controls. Results are presented as means ± SE. Leaf node 1 is closest to the apex [5d].
Figure 1.
In both compatible interactions (A, C) and incompatible interactions (B, D) between tomato and Pseudomonas syringae, PPO suppression results in enhanced susceptibility to infection. Antisense PPO transgenic plants have higher bacterial growth (A, B) and lesion numbers (C, D) than nontransformed controls. Results are presented as means ± SE. Leaf node 1 is closest to the apex [5d].

We have demonstrated a role for tomato PPO in resistance against P. syringae pv. tomato in both compatible and incompatible plant-pathogen interactions. Transgenic tomato plants with 40- to 180-fold reductions in PPO activity levels and nondetectable PPO mRNA levels (A14-6) showed markedly enhanced susceptibility to the pathogen compared to NT controls in both compatible and incompatible plant-pathogen interactions. In the compatible interaction, bacterial growth in A14-6 foliage was up to 55 times greater (p<0.01) than in NT controls (Figure 1A). In the incompatible interaction, where a resistant Pto gene was transferred to A14-6 and NT plants and subsequently tested against an avirulent strain of P. syrinagae, the differences in P. syrinagae populations between foliage of antisense PPO transgenic tomato plants and NT controls were more pronounced (up to 250 fold higher; Figure 1B). The number of lesions formed by P. syrinagae was similarly affected by the absence of PPO. In both compatible and incompatible interactions, antisense suppression of PPO resulted in up to 10-fold increases in lesion numbers (Figure 1C, D [5d]).
PPO-generated quinones and ROS can play an array of defense-related functions simultaneously. In both compatible and incompatible plant-pathogen interactions, PPO may act after cell decompartmentalization as a component of cell death responses, with quinone production as one of the terminal events in the progression of such responses. Direct toxicity to plant and pathogen cellular macromolecules via quinone-mediated covalent modification or direct anti-microbial toxicity of H2O2 in the vicinity of pathogen attack may restrict disease progression. Moreover, protein bioavailability to pathogens may be reduced due to the alkylation of plant proteins. At the same time, a physical pathogen barrier could be generated as a result of cross-linking of oxidized phenolics as well as of oxidative cross-linking of cell wall proteins and enhanced lignin formation.
In addition to the direct defense response, PPO-generated H2O2 could also be a component of signaling processes by acting as a diffusible inducer of cellular protectant genes, phytoalexin biosynthesis, and salicylic acid and ethylene production, as well as a trigger of cell death resulting in restricted lesions delimited from surrounding healthy tissue. In addition, other plant defense genes and ultimately plant immunity may be elicited by ROS-mediated systemic signaling network [20, 5d].
The role of PPO in P. syringae resistance was further substantiated when transgenic tomato plants with 5-10 fold increase in PPO activity levels showed over 15-fold fewer lesions and over 100 fold reduction in P. syrinagae growth compared to NT control at a high inoculum concentration [5c]. Although the mechanism by which PPO contributes to disease limitation is still unclear, it is evident that PPO may participate in plant defense as a component of both the response and signaling process that ultimately limits disease progression.
The efficiency of this defense mechanism, however, appears to be dependent on the type and virulence of the target pathogen. Preliminary observations of the NT and transgenic SP and OP tomato genotypes in our laboratory suggested a positive correlation between PPO activity and levels of foliar resistance to Alternaria solani, the causal pathogen of early blight (unpublished data), similar to the correlation found for P. syringae. In contrast, when A14-6 and NT control seedlings were evaluated for resistance against Ralstonia solanacearum, the causal agent of bacterial wilt in tomato, no significant differences in susceptibility were observed between the 2 genotypes (J. Chunwongse, personal communication). While R. solanacearum mainly attacks and moves through vascular system of tomato, where accumulation of PPO is limited to phloem [10a], P. syringae infects epidermal, mesophyll and cortical cells of tomato foliage and stems in which PPO has been shown to accumulate abundantly, particularly in young tissues [10a]. Moreover, PPO F was found to be induced locally surrounding P. syringae lesions and systemically in foliage and stems [6c, 5d].
PPOs have also been proposed to confer resistance to various diseases in plant species in addition to tomato. Most of the studies involving other species associated levels of PPO activity with disease resistance or found induction of PPO activity in response to pathogen infection. In bean, a positive correlation was observed among activities of PPO, PO, and levels of phenolics and resistance to anthracnose, caused by Colletotricum lindemuthianum. This pathogen also elicited increases in PPO and PO activities in bean cultivars with higher resistance [21]. Similar positive correlations between PPO activity and disease resistance, and PPO induction upon infection were found in the pearl millet-Sclerospora graminicola and wheat-Alternaria triticina interactions [22]. In banana, leaf area without leaf streaks caused by Mycosphaerella fijiensis was positively correlated with the activities of PPO, phenylalanine ammonia-lyase (PAL), ascorbic acid oxidase and catalase [23].
Biochemical analysis of isogenic Indian mustard lines varying in resistance levels to Albugo candida revealed an association of white rust resistance with PPO and PO activities. In resistant lines, PO and PPO activities also increased upon infection [24]. After potato tubers were infected with an incompatible strain of Phytophthora infestans, PO and PPO activities were found to be induced [25]. Similarly, infection of pear fruits and wheat heads by Erwinia amylovora and Fusarium graminearum, respectively, increased PPO activity levels in both compatible and incompatible interactions [26]. PPO activity induced by abiotic factors may help confer resistance to subsequent pathogen infection. Stahmann et al. [27] found that ethylene-induced resistance to infection by Ceratocystis fimbriata coincided with the increase in PPO and PO activities in sweet potato root tissue. Root application of Si also enhanced activities of PPO, PO and chitinase in foliage of Podosphaera xanthii-inoculated cucumber and enhanced suppression of subsequent infection by the same pathogen [28].
The role that PPO plays in plant-microbe interactions appears to be complex. Even symbiotic microorganisms such as mycorrhizae could induce PPO activity in some plant species including Ziziphus xylopyrus and Moringa concanensis; such induction could presumably be beneficial to plant hosts by protecting them from attacking pathogens [29]. Application to tomato of a biocontrol agent, P. fluorescens, also induced PPO, PO and PAL activities and resulted in significantly reduced infestation of root-knot nematode [30]. Similarly, Trichoderma-treated bean also exhibited induced P. syringae resistance concomitant with PPO and PO induction [31].
Several transgenic approaches have been utilized to produce plants with increased resistance to various pathogens. Modification of plants using genes other than PPO may also affect PPO activities, with consequences for plant defense. For example, transgenic potatoes expressing pectate lyase from Erwinia carotovora provided enhanced resistance to soft rot-causing Erwinia bacteria. The spreading of the bacteria was prevented presumably because of the enhanced PPO activity [32]. Transgenic cotton, tobacco and potato expressing elevated levels of H2O2 , an inducer of PPO and other defense genes, showed reduced development of many fungal diseases including Rhizoctonia, Verticilium, Phytophthora and Alternaria spp. [33].
Role of PPO in insect resistance
A variety of evidence suggests that PPO plays a role in plant resistance to arthropod herbivores. Much of this evidence is analogous to the evidence gathered for the role of PPO in disease resistance. PPO activity is correlated with resistance to arthropod herbivores in some plants (e.g., [5a, 34]), and inhibition of PPO activity by antioxidants and other chemical inhibitors improves the growth rates of insects feeding on plant-derived diets high in PPO activity [5a]. PPO can also reduce the growth of Lepidopteran larvae when incorporated in artificial diet together with an appropriate phenolic substrate [5a, 35]. Moreover, PPO activity is inducible in many plants by wounding and arthropod herbivory and plants with induced PPO activity often show increased resistance against a broad spectrum of arthropod herbivores. In tomato, for example, treatment with jasmonic acid (JA) or wounding by chewing insects, which are both known to induce PPO activity as well as a number of other responses, results in a broad-spectrum increase in resistance that extends to other chewing insects, mites, aphids, thrips and even a bacterial pathogen [5b, 36]. Recently, mutants and transgenic plants with modified expression of defense genes including PPO have been shown to possess altered levels of insect resistance. Transgenic tobacco overexpressing TobpreproHypSys-A, encoding the hydroxyproline-rich glycopeptide systemin precursor protein, had enhanced resistance against Helicoverpa armigera larvae that was suggested to stem from induced accumulation of proteinase inhibitors (PIs) and PPO [37]. In contrast, a tomato mutant, jasmonic acid-insensitive 1 (jai1), which is defective in JA signaling, and unable to express PPO and PIs in response to wounding or JA, exhibited severely compromised resistance against the two-spotted spider mite [38].
As is the case with PPO-mediated resistance to plant pathogens, PPO is thought to exert its negative effects on insects via a variety of mechanisms, although experimental evidence for many of these putative mechanisms is lacking. Quinones and ROS generated as a result of PPO activity may be directly toxic to arthropods as well as pathogens, and H2O2 and other ROS may influence resistance to arthopods indirectly by participating in plant signaling networks related to arthropod resistance. The cross-linking of cell walls and increased lignification of plant tissues that occur as a result of enhanced PPO activity may interfere with nutrient acquisition by herbivores. The mechanism that has received the most attention with respect to herbivore resistance, however, is the ability of quinones produced by PPO activity to alklylate (arylate) dietary protein, thereby rendering plant protein unavailable to herbivores and reducing the nutritional value of plant tissues. Felton et al. [5a] found that oxidation of chlorogenic acid by PPO in tomato foliage reduced levels of free NH2 groups and Biorad-detectable protein in foliage and that these effects on dietary protein were associated with a reduction in the growth rates of beet armyworm (Spodoptera exigua) larvae on foliage. Felton et al. [39] further showed that many of the amino acids most susceptible to derivatization by PPO-generated quinones (i.e., amino acids with nucleophilic centers such as cysteine, methionine, lysine, and histidine) are limiting for the growth of Lepidopteran larvae.
The use of transgenic plants with altered PPO activity represents a powerful approach for investigating the role of PPO in plant resistance to arthropods. The use of such plants allows activities of PPO to be manipulated independently of other plant constituents that may affect plant resistance. This is particularly important when attempting to determine the role of PPO in induced resistance, because the contribution of induced PPO activity to induced resistance is obscured by the concurrent induction of many other allelochemicals and defense-related proteins such as PIs. Expressing PPO constitutively thus removes some of the ambiguity involved in discerning causal relationships in plant resistance (although it may create additional ambiguity because, given the putative multifunctionality of PPO in plants, altering PPO levels may have unintended effects on physiological process and on primary and/or secondary metabolism). Despite the potential utility of the transgenic approach, published examples have thus far been limited to a single plant species, poplar (Populus sp.). Wang and Constabel [5e] showed that overexpression of PPO in Populus plants resulted in higher mortality and reduced weight gain of larvae of the forest tent caterpillar (Malacosoma disstria) when larvae from older egg masses were used. In a second study, Barbehenn et al. [5f] found no reductions, or only slight reductions (6.5% to 16%) in relative consumption rates and relative growth rates of two species of caterpillar (Lymantria dispar and Orgyia leucostigma) feeding on PPO-overexpressing poplar. Clearly, additional studies in systems other than poplar are needed to gain a clear picture of the role of PPO in plant defense.
We have recently conducted a series of studies on the effects of PPO overexpression and underexpression in transgenic tomato plants on the growth and development of three Lepidopteran pests: common cutworm (Spodoptera litura (F.)), beet armyworm (S. exigua (Hűbner)), and cotton bollworm (Helicoverpa armigera (Hűbner)) (Mahanil et al., unpublished manuscript; Bhonwong et al., unpublished manuscript). In addition, the effects of these transgenic tomato plants on the Colorado potato beetle (Leptinotarsa decemlineata (Say)), were investigated in an earlier set of experiments (M. Hunt, J. Steffens, unpublished data). All four insects are widely-distributed pests of a large number of crop plants (including tomato) and all four cause millions of dollars in losses annually. Control measures for all four pests are expensive and environmentally damaging, and insecticide resistance is an additional concern for all four pests. Therefore, development of alternative control methods against these pests is needed.
The most extensive research on the effects of transgenic tomato plants with altered PPO activities on insect growth was conducted using S. litura (Mahanil et al., unpublished manuscript). Insect growth assays were conducted using both young leaves and old leaves of the NT, OP and SP tomato plants described above. Under the conditions of the experiments conducted using S. litura, SP plants exhibited up to 7.3-fold reductions in PPO activity, whereas OP plants showed increases of up to 5.7 fold compared to NT controls. In addition, old leaves (leaves from the eighth node of tomato plants, with the youngest leaf node counted as node 1) had lower PPO activities than young tomato leaves (leaves from node 4). In an initial experiment, S. litura larvae were placed as neonates on excised tomato leaves and allowed to develop until the fourth instar. In this experiment, simple growth rates (weight gains per day) of larvae were up to 60% lower on leaves of OP plants than on leaves of NT controls, whereas simple growth rates of larvae placed on SP plants were up to 37% higher than on leaves of NT controls. Larvae also consumed less plant material when placed on leaves of OP plants than when placed on NT controls, and consumed more leaf material when placed on leaves from SP plants. Although mortality in this experiment was low, there was a trend toward higher mortality on OP leaves than on controls. In a subsequent experiment in which larvae were allowed to develop until pupation, similar effects of PPO over- and underexpression on insect growth, feeding, and mortality were observed. In addition, larval development time was generally longer when feeding on leaves of OP plants. Pupal weights did not differ significantly among tomato genotypes. Finally, a third experiment provided support for the hypothesis that PPO reduces the nutritional value of foliage for Lepidopteran larvae. Relative growth rates (weight gains per gram of body weight per day) of third instars feeding on young leaves of OP plants decreased, and relative consumption rates increased, with increasing PPO activities. The latter result demonstrates that greater leaf consumption by larvae
![Molecules 12 01569 g002]() on SP plants in previous experiments was simply a consequence of larger larval size on these plants, and furthermore demonstrates that the effects of PPO on larvae were not the result of an antifeedant effect. Moreover, efficiencies of conversion of ingested and digested food also differed among larvae placed on different tomato genotypes; conversion efficiencies were higher on both young and old leaves of SP plants than on leaves of NT plants, and conversion efficiencies were lower on both young and old leaves of OP plants than on corresponding leaves of NT plants. Taken together, these results are consistent with the hypothesis that higher PPO activities interfere with the bioavailability of nutrients by complexing with nucleophilic amino acids in protein (although they do not exclude other mechanisms).
on SP plants in previous experiments was simply a consequence of larger larval size on these plants, and furthermore demonstrates that the effects of PPO on larvae were not the result of an antifeedant effect. Moreover, efficiencies of conversion of ingested and digested food also differed among larvae placed on different tomato genotypes; conversion efficiencies were higher on both young and old leaves of SP plants than on leaves of NT plants, and conversion efficiencies were lower on both young and old leaves of OP plants than on corresponding leaves of NT plants. Taken together, these results are consistent with the hypothesis that higher PPO activities interfere with the bioavailability of nutrients by complexing with nucleophilic amino acids in protein (although they do not exclude other mechanisms).
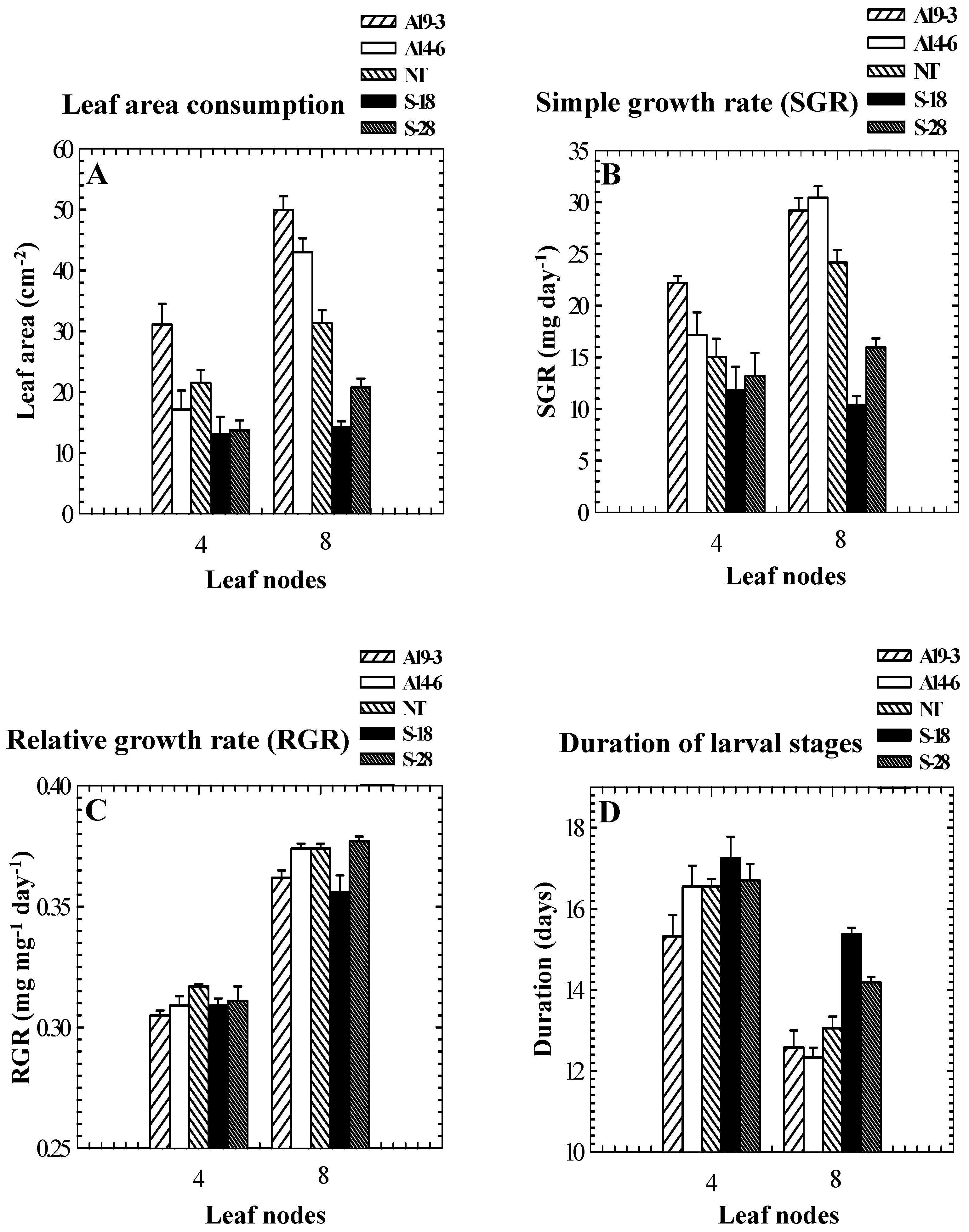
Figure 2.
Leaf area consumption (A) and simple growth rates (B) of beet armyworms (Spodoptera exigua) feeding on nodes 4 and 8 leaves of tomato vary in different genotypes differing in PPO activity levels. Relative growth rates (C) are not related to PPO activities. Durations of larval stages (D) of beet armyworms are similar among genotypes when fed on node 4 leaves, but are extended on OP plants and attenuated on SP plants compared to controls when fed on node 8 leaves. Results are presented as means ± SE. Leaf node 1 is closest to the apex.
Figure 2.
Leaf area consumption (A) and simple growth rates (B) of beet armyworms (Spodoptera exigua) feeding on nodes 4 and 8 leaves of tomato vary in different genotypes differing in PPO activity levels. Relative growth rates (C) are not related to PPO activities. Durations of larval stages (D) of beet armyworms are similar among genotypes when fed on node 4 leaves, but are extended on OP plants and attenuated on SP plants compared to controls when fed on node 8 leaves. Results are presented as means ± SE. Leaf node 1 is closest to the apex.
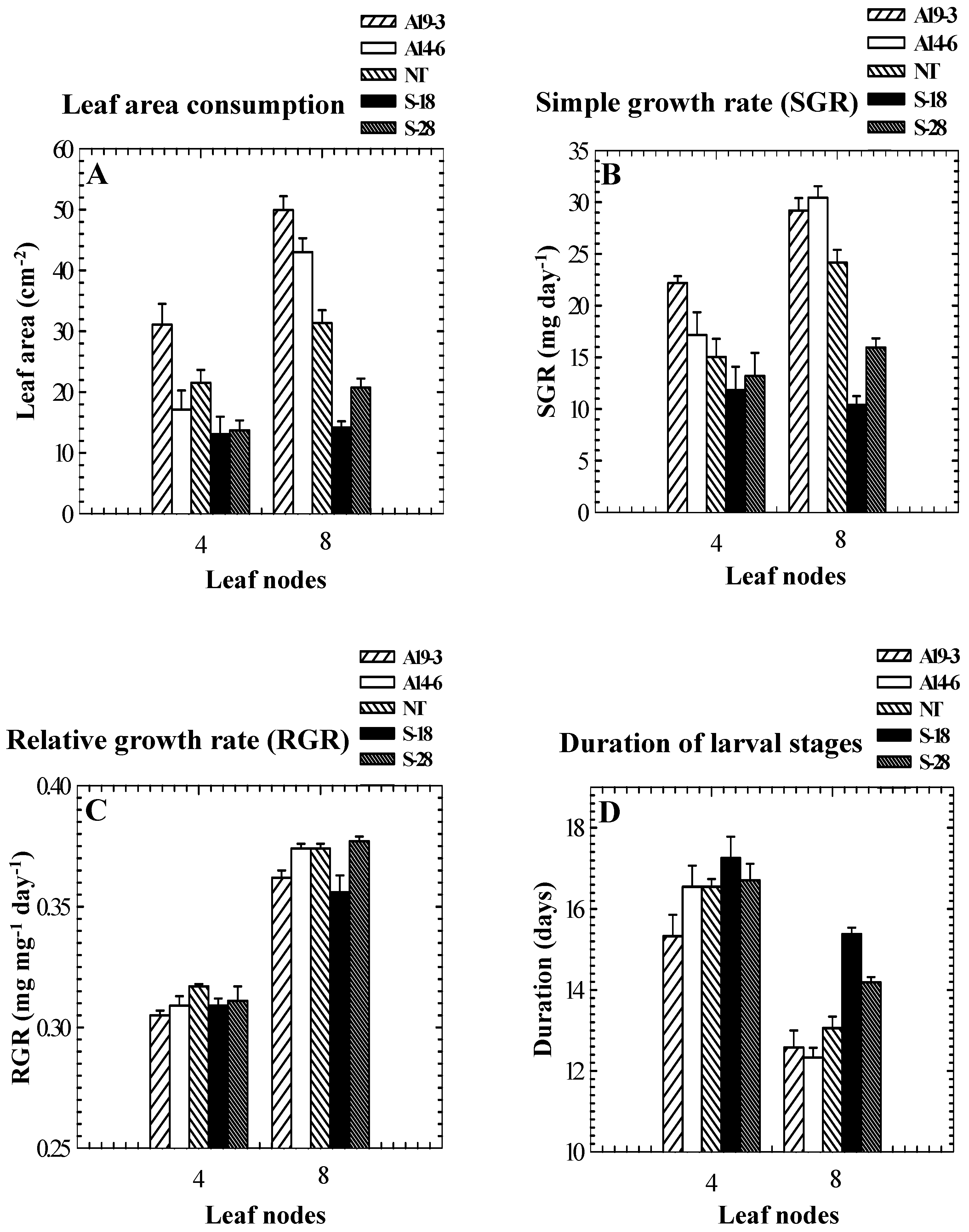
Results obtained with the other two Lepidopteran species, S. exigua and H. armigera, generally mirrored the results obtained with S. litura. For S. exigua feeding on old (node 8) leaves, leaf areas consumed and simple growth rates were higher on leaves from SP plants and lower on leaves from OP plants compared to consumption and growth on leaves from NT control plants (Figure 2A, B). Pupal weights and mortality did not differ among treatments, but development time was extended on OP plants and attenuated on SP plants compared to controls (Figure 2D, Bhonwong et al., unpublished manuscript). However, no relationship between relative growth rates and PPO activities was found (Figure 2C). For S. exigua feeding on young (node 4) leaves, effects on larvae feeding on leaves from SP plants were similar but effects on larvae feeding on OP plants were not as pronounced (Figure 2A, B). There were no significant effects of plant genotype on pupal weights and development time for larvae feeding on node 4 leaves (Figure 2D, Bhonwong et al., unpublished manuscript). The fact that the effects of PPO over- and underexpression on larvae differed for larvae feeding on young and old leaves is consistent with previous studies showing that the effects of PPO on nutritional quality are dependent on quality and quantity of dietary protein (e.g., Felton et al. [39]), because young and old leaves used in this study likely differed in both quality and quantity of protein.
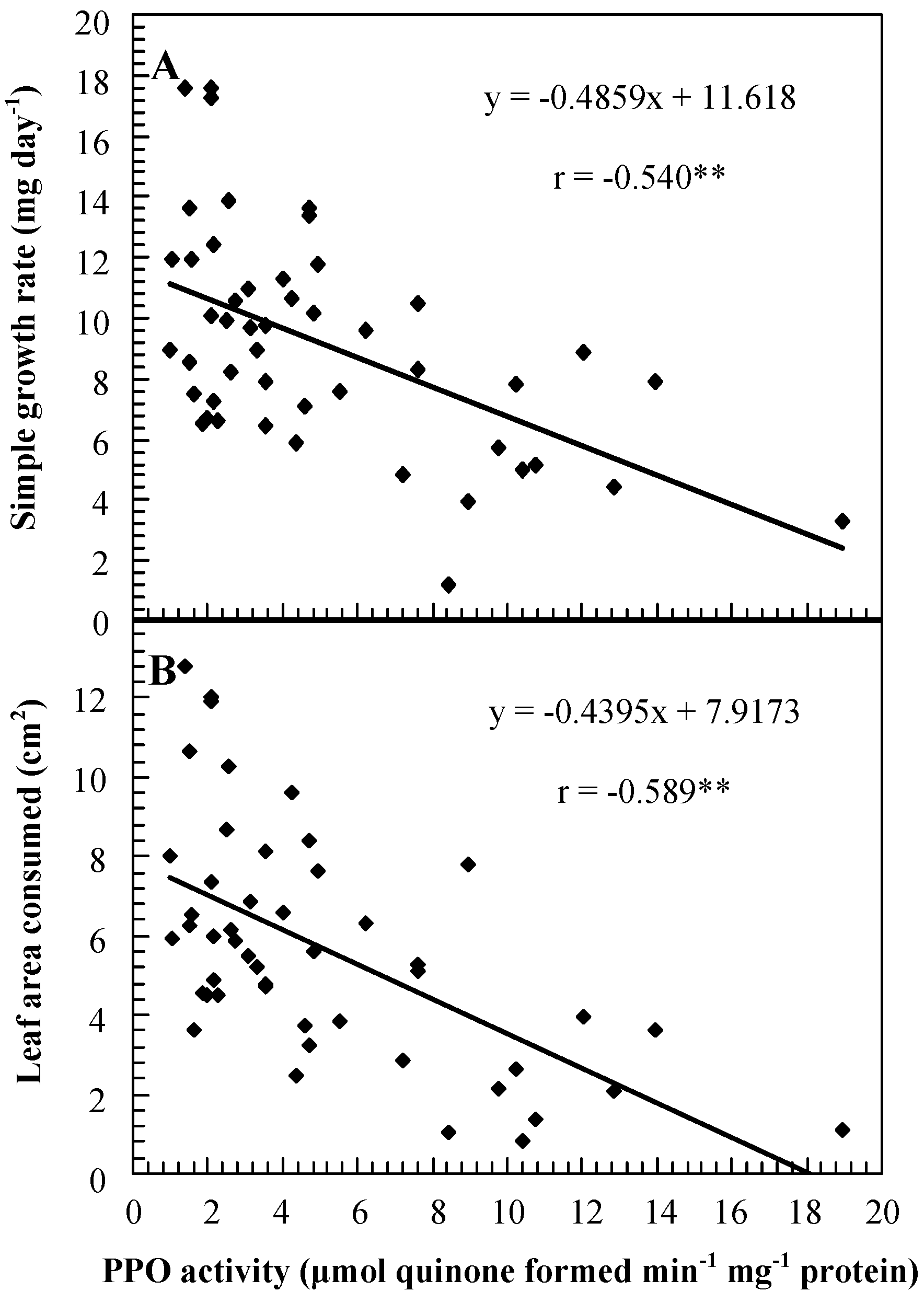
Results obtained with H. armigera were similar to results obtained with the other two species: leaf areas consumed, survivorship, simple growth rates, and relative growth rates were generally higher on SP plants than on controls and were generally lower on OP plants than on controls. Growth rates of, and leaf areas consumed by, neonate larvae placed on young excised foliage of SP and OP genotypes were negatively correlated with PPO activities (Figure 3).
In two feeding trials conducted with the Colorado potato beetle (Table 2), antisense downregulation of PPO activity resulted in significant reductions in larval mortality and significant increases in weight gained and foliage consumed compared with controls. These significant effects were observed in a feeding trial conducted in summer but not in a trial conducted in winter, when PPO activities in the control plants were much lower.
Thus, data obtained using transgenic tomato plants that exhibit both higher PPO activities than wild-type plants and lower PPO activities than wild-type plants are consistent with the hypothesis that PPO is involved in the resistance of tomato plants to these Lepidopteran and Coleopteran species and that the increases in resistance of tomato plants to these insects following JA treatment or wounding are at least partially attributable to the increases in PPO activities also observed following JA treatment and wounding. Studies are ongoing with other types of herbivores, and results from past research indicate that PPO activities will also be correlated with resistance to these pests as well.
Figure 3.
Correlation between PPO activities and simple growth rates of cotton bollworms (Helicoverpa armigera) feeding on leaflets at node 4 of transgenic plants with suppressed and overexpressed PPO activity segregating for PPO activity levels (A), and between PPO activities and leaf areas consumed (B).
Figure 3.
Correlation between PPO activities and simple growth rates of cotton bollworms (Helicoverpa armigera) feeding on leaflets at node 4 of transgenic plants with suppressed and overexpressed PPO activity segregating for PPO activity levels (A), and between PPO activities and leaf areas consumed (B).
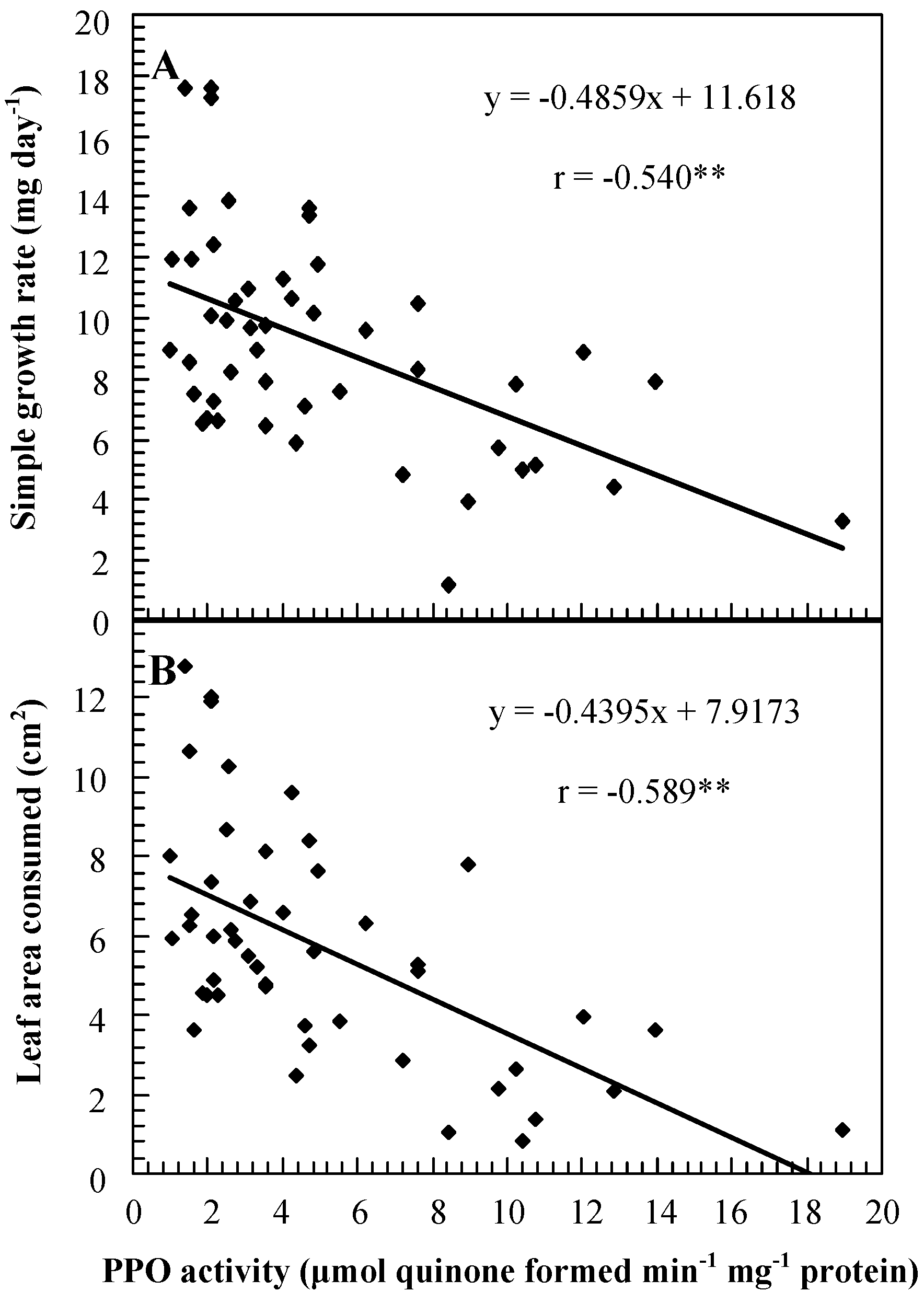
Table 2.
Percent mortality, weight gain and foliage consumption of Colorado potato beetle feeding on transgenic tomatoes with suppressed PPO activity and nontransformed controls.
| Genotype | PPO activity (μmol quinone formed min-1 mg-1 protein) 1 | Percent mortality | Weight gain (mg)/larva | Foliage consumption (mg)/larva | Date of feeding assay |
|---|---|---|---|---|---|
| A14-6 2 | ND 3 | 45 a | 43 ± 1.9 a 4 | 123 ± 6.4 a | July 1993 |
| NT | 16.0 | 68 b | 32 ± 4.1 b | 76 ± 5.0 b | |
| A14-6 | ND | 3 | 50 ± 2.6 | 131 ± 12.3 | Dec. 1993 |
| NT | 7.1 | 10 | 44 ± 3.7 | 115 ± 13.7 |
- 1
- Values are from leaf nodes 1-2 and do not reflect the PPO activity of foliage fed to larvae (nodes 3-6).
- 2
- A14-6, A19-3, transgenic lines with suppressed PPO activity; NT, nontransformed control; S-18, S-28, transgenic lines overexpressing PPO activity.
- 3
- ND: not detectable.
- 4
- Data are presented as means ± SE. Data not followed by the same letter in a column are significantly different (p<0.05).
At the same time, the lack of effect of altered PPO levels on some measures of insect performance and the failure of altered PPO levels to affect some insects in some plants species and in some environmental contexts is a reminder that plant resistance to arthropods is a complex phenomenon, and that no single biochemical trait is likely to confer high levels of resistance to all arthropods in all environments. This caveat aside, transgenic tomato plants that overexpress PPO may have a place in insect pest management programs in tomato. This possibility is further supported by a study showing that PPO synergizes the activity of Bt toxin by akylating the crystal protein and thereby increasing its toxicity [40]. However, enhanced PPO activity may not be compatible with all management strategies, as phenolase activity in cotton and tomato has been shown to reduce the efficacy of baculoviruses toward Lepidopterans [41].
Role of PPO during water stress
Due to the association of PPO with thylakoid membranes, the high Km of PPO for O2, and the association between induction of PPO activity and O2 evolution in isolated chloroplasts, PPO in the mesophyll chloroplast has been proposed to have a role in the Mehler reaction, photoreduction of molecular oxygen by PSI, and regulation of plastidic oxygen levels [42,3]. The Mehler reaction is a potentially important nondestructive sink for excess photosynthetic electrons under conditions of water stress when carbon assimilation decreases because of stomatal closure, and hence may help prevent over-reduction of components of linear electron transport [43]. When excess excitation energy is not dissipated by protective mechanisms, it is used to form cytotoxic ROS [44]. Photoinhibition and ultimately photooxidative damage will occur if the combined capacity of ROS scavenging systems is exceeded. Photoinhibition is commonly detected as a decrease in the ratio of variable fluorescence (Fv) to maximal fluorescence (Fm) of PSII reaction centers measured on dark-adapted leaves [45]. If PPO contributes to the Mehler reaction, plants with elevated PPO are expected to have improved stress tolerance.
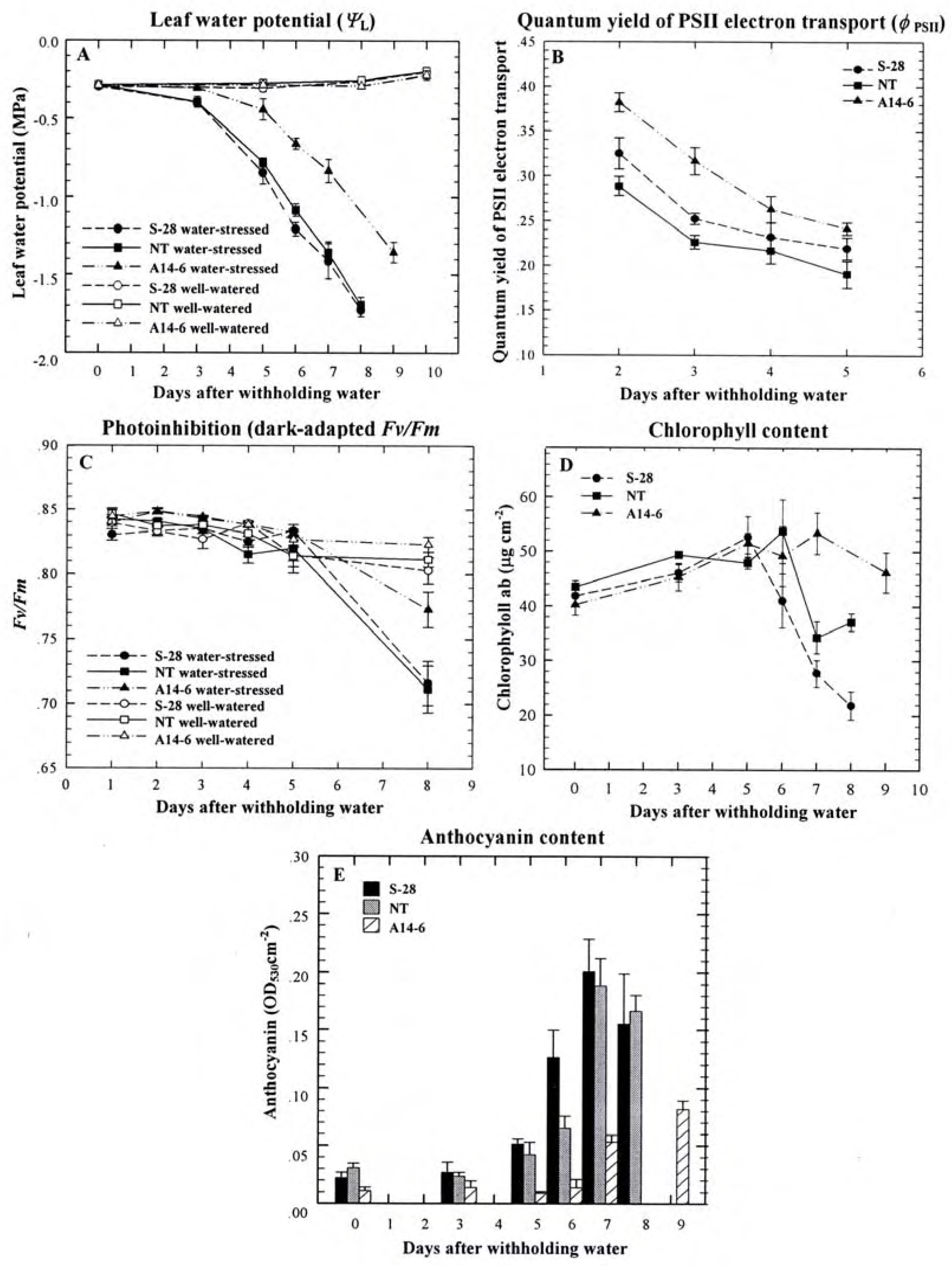
In a previous report, we evaluated the response of transgenic A14-6 plants with 20 to 40-fold decreased PPO activity and transgenic S-28 plants with 2 to 6-fold increased PPO activity to drought stress [13]. Briefly, it was found that A14-6 plants began exhibiting stress symptoms --wilting, leaf curling and yellowing of old leaves-- later than S-28 and NT plants under similar drought stress conditions. At most periods after initiation of drought stress, water stress, measured by ΨL, was significantly less negative in A14-6 plants (Fig.4A), and stomatal conductance (gs) of stressed plants declined more slowly in A14-6 plants compared to S-28 and NT plants [13]. These A14-6 plants delayed photoinhibition by generally maintaining higher light adapted (φPSII) and dark adapted (Fv/Fm) quantum yields of PSII photochemistry compared to NT and S-28 plants when drought stress was imposed on (Figure 4B, C). Moreover, they delayed photooxidative damage and suffered chlorophyll (a+b) losses in older leaves to a lesser extent than did S-28 and NT plants as drought stress progressed (Figure 4D [13]). The higher levels of φPSII, Fv/Fm and total chlorophyll content later in the stress that was observed in A14-6 plants, compared to NT and S-28, suggested that lower photooxidative stress and rate of ROS production, which was reflected by the lower content of anthocyanin, a putative antioxidant, occurred in these plants (Figure 4E). Since A14-6 rather than S-28 plants exhibited more favorable water relations and decreased photoinhibition, PPO was proposed to have a role in the development of plant water stress and potential for photoinhibition and photooxidative damage that may be unrelated to any effects on the Mehler reaction.
Figure 4.
Leaf water potential (ΨL) during water stress declines more slowly in transgenic plants with suppressed PPO levels (A). During water stress these plants exhibits higher quantum yields of PSII electron transport (φPSII; B) but lower photoinhibition (C) compared to nontransformed controls. Suppression of PPO reduces chlorophyll loss (D) and anthocyanin accumulation (E) in leaflets at node 7. Results are presented as means ± SE [13].
Figure 4.
Leaf water potential (ΨL) during water stress declines more slowly in transgenic plants with suppressed PPO levels (A). During water stress these plants exhibits higher quantum yields of PSII electron transport (φPSII; B) but lower photoinhibition (C) compared to nontransformed controls. Suppression of PPO reduces chlorophyll loss (D) and anthocyanin accumulation (E) in leaflets at node 7. Results are presented as means ± SE [13].
Expression analysis showed that PPO was induced in response to water stress with the highest magnitude of induction in older leaves and corresponding abscission zones, which might facilitate cell death preferentially in these tissues. The abscission of older leaves might represent an adaptive strategy under water stress since it could reduce further water loss and allow limited nutrients to be partitioned to younger tissues [13]. Induction of PPO activities under water stress has been previously demonstrated in tomatoes [46]. In other plant species such as coconut, increased PPO was also observed under water stress condition [47]. However, further investigation is needed to understand the physiological role of PPO during water stress more clearly.
Not only water stress but UV radiation and salinity could also impose oxidative stresses on plants. Niknam et al. [48] found increase in PPO activity of Trigonella foenum-graecum calli cultured in media containing NaCl. However, in UV-B treated tomato, PPO activity was reduced to 42%, presumably to minimize oxidative stress injury while the contents of phenols and anthocyanins, the antioxidants, in the cells increased [49]. It appears that the well-defined responsiveness of PPO to various abiotic and biotic stresses, depending on plant genotype and environmental as well as ecological context, may be an adaptive strategy of plants to cope with a multitude of stresses that often occur simultaneously.
Other possible roles
Sherman et al. [50] proposed that PPO may have developed simultaneously with the adaptation to an oxygenated atmosphere where its function is vital in the photosynthetic apparatus of terrestrial plants. Since then PPO genes may have evolved to perform diverse functions in different plant species. A role for PPO in pigment formation was suggested in snapdragon flowers for biosynthesis of aurones (yellow pigment), and in Caryophyllales order for biosynthesis of betalains (yellow or red pigments [4,8]). Heterologous expression of aureusidin synthase (PPO) and chalcone 4’-o-glucosyltransferase allowed the accumulation of aureusidin 6-o glucoside and, hence, yellow pigments in transgenic flowers of torenia (Torenia hybrida) normally lacking this endogenous biosynthetic mechanism [11]. In creosote bush (Larrea tridentate), Cho et al. [51] have shown a role for PPO in biosynthesis of 8 to 8’ linked lignans, metabolites that have potent antiviral, anticancer and antioxidant properties.
Due to the ability of PPOs to convert monophenols to o-diphenols, PPOs have also been assumed to have a function in hydroxylation of p-coumaric acid to caffeic acid in the phenylpropanoid pathway (i.e., coumarate 3-hydroxylase [42b, 2]). However, no conspicuous differences in leaf p-coumaric acid and caffeic acid pools between A14-6 and NT were found, suggesting that PPO does not contribute to coumarate 3-hydroxylation in leaves [5d]. Moreover, disruption of plastid PPO uptake and processing by tentoxin treatment indicated that PPO is not involved in coumarate hydroxylation [52]. Recent identification of a cytochrome p450 p-coumarate 3-hydroxylase [53] and the localization of PPO on the luminal side of thylakoid membrane [50,54] further negated this hypothesis.
Since we did not observe phenotypic effects in any SP and OP transgenic lines that were consistent with involvement of PPO in photosynthesis, we conclude that role of PPO in this area is minimal. By BLAST searching the Arabidopsis genome, Sullivan et al. [9n] also found no apparent genes encoding PPO, challenging an indispensable role for PPO in chloroplast function. In forage crops such as red clover and alfafa, PPO inhibits postharvest proteolysis at the early stage of ensiling, possibly by protease inactivation through reaction of quinones with nucleophilic sites. Proteolysis of transgenic red clover plants whose PPO expression was silenced by expressing double-stranded PPO RNA (RNAi) was dramatically increased whilst browning was delayed, compared to nontransformed controls [55]. Whereas heterologous expression of a red clover PPO gene in alfafa significantly reduced proteolysis and caused more rapid browning in alfafa leaf extracts in the presence of o-diphenol, compared to controls [9n].
Other roles such as producing phenolic signal molecules in specific developmental flower processes, regulating flowering via IAA catabolism, maintaining the turnover rate of phenols during oxidative stresses, and catalyzing the polymerization of phenols in hydropotes for the trapping of cadmium crystals in some plant species have also been suggested, however, no conclusive evidence has yet been provided for any of these roles [49, 9i, 56].
Application of transgenic plants with modified PPO activity
Because of the conspicuous brown or black pigments formed and/or degradation of lycopene as a result of PPO-catalyzed reactions during the harvesting, postharvest storage and processing of many commercially important vegetable fruit, and cereal crops [1b, 57], many attempts have been made to downregulate PPO in various plant species. In potato, downregulation of PPO using antisense/sense technology with potato or tomato PPO genes has been a successful approach to reduce tuber browning (black spot formation) from internal damage during mechanical harvesting and storage that may deliver significant savings to the fresh, frozen and processed potato industry [58]. In order to develop Sultana grape cultivars which yields premium golden colored raisins when dried instead of the PPO-imparted brown colored ones, transgenic grape with suppressed PPO activity in fruits was generated using antisense technology [59]. Field trials of this transgenic grape, as well as the antisense PPO transgenic pineapple generated to reduce discoloration of fruit (blackheart) following harvesting and low temperature storage, have been conducted in Australia [59b, 60]. Apple, which is notorious for browning, is another target plant for PPO downregulation. The antisense PPO gene or gene encoding PPO RNAi was transformed into apple to produce transgenic apples with lower browning potential in shoots and fruits when sliced, bruised, scuffed or bitten [61]. In sugarcane, overexpression of PPO resulted in darker juice and raw sugar, suggesting that suppression of PPO activity may result in lighter colored sugar and minimize the production cost. Though antisense transgenic sugarcane failed to suppress PPO activity and reduce the color intensity of juice and raw sugar, further attempts to lower PPO activity may still be warranted [62]. It was shown that sapburn (browning, blackening and necrosis of the skin) in mango fruits was induced by the sap and catalyzed by PPO in the skin [63]. This problem might be alleviated by producing transgenic mango with reduced PPO activity in this tissue. In addition, PPO-catalyzed browning is not welcome in many other crops: it causes discolored white wine, and browning in lettuce, banana and beans; the latter usually require optimal blanching. Moreover, discoloration of cabinet-making timbers, the need for bleaching in paper-making as well as off-coloring and browning of cherry stems may be reduced if PPO activity is suppressed. Attempts to develop commercialized suppressed PPO transformants to minimize browning are currently underway for some of these plant species [59a, 64].
In some cases, overexpression of PPO may be more beneficial than downregulation. For example, heterologous expression of red clover PPO gene in alfafa reduced postharvest proteolysis during the early stages of ensiling, thereby reducing the loss of nutritional value [9n]. Moreover, as discussed above, enhanced resistance may be obtained in some plant-pest interactions. For example, transgenic tomato overexpressing PPO had enhanced bacterial disease resistance [5c], and transgenic poplar overexpressing PPO has been shown to provide enhanced resistance to forest tent caterpillar [5e].
Conclusion and future prospects
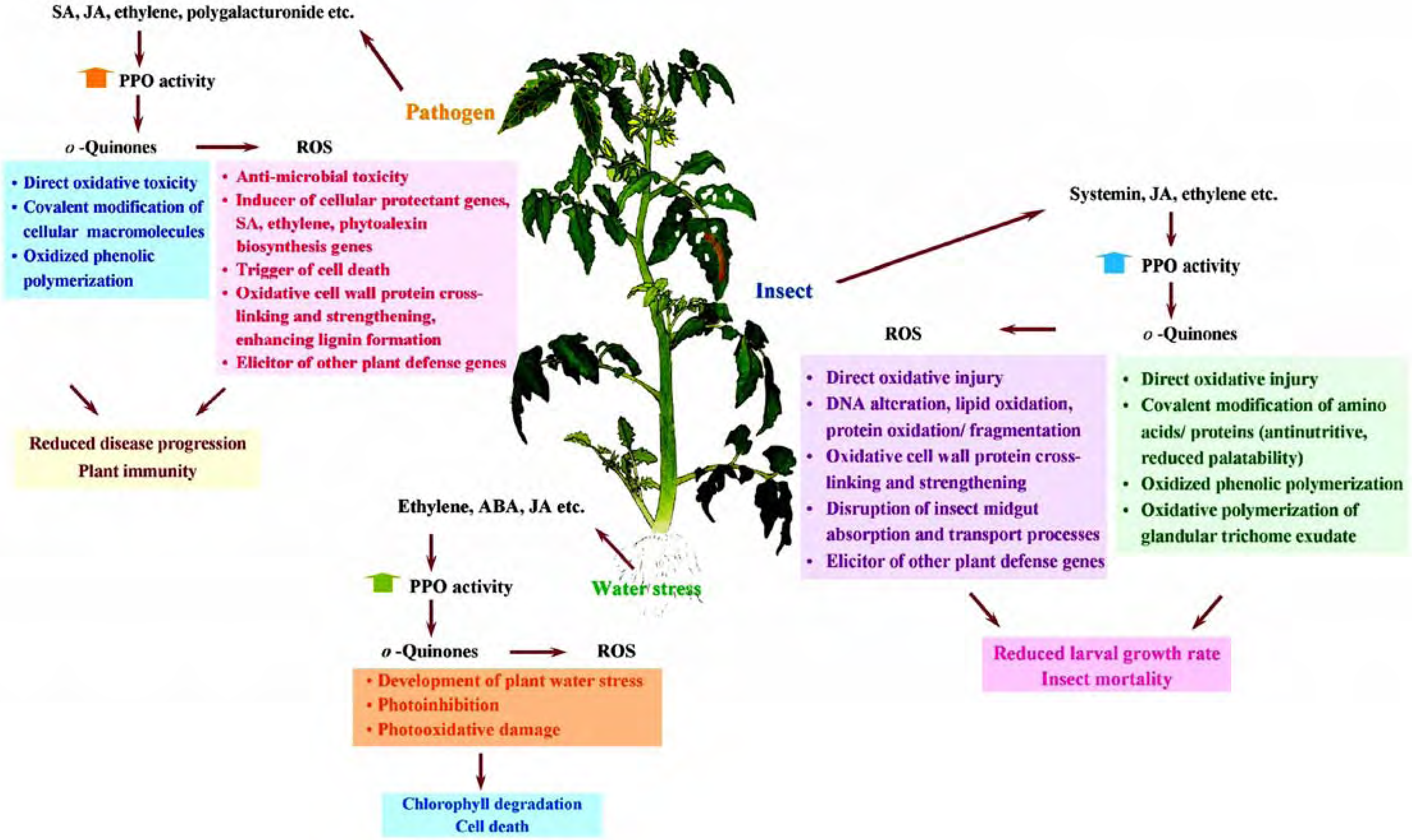
We have utilized antisense/sense technology as a powerful tool for establishing conclusive roles of tomato PPO in defense against pathogens and various insect herbivores, as well as evaluating a putative role of PPO during water stress. Figure 5 summarizes possible mechanisms of action of PPO in response to these biotic and abiotic stresses. In nature, it is very likely that PPO may play different roles simultaneously and plant responses to one stress may synergize or antagonize the responses to other stresses. Antisense/sense technology has also been used for functional analysis of PPO in other plant species, and roles for PPO in pigment biosynthesis, inhibition of postharvest proteolysis, and insect resistance have been investigated [9n, 5e, 11, 55]. Since PPOs appear to have different functions in different plant species, using this technology specifically in other plant species will shed more light on the physiological functions of PPO. Functional analysis of individual PPO gene members within the same species whose sequences are highly similar is a more challenging task and might require additional technology such as RNAi for inhibiting specific genes.
Figure 5.
Hypothetical mechanisms of action of tomato PPO in response to pathogen infection, insect infestation and water stress.
Figure 5.
Hypothetical mechanisms of action of tomato PPO in response to pathogen infection, insect infestation and water stress.
In addition to functional analysis, antisense/sense technology also has commercial applications, in particular the downregulation of PPO expression to enhance quality attributes of several crop products. However, unexpected phenotypes could arise if critical but overlooked roles of PPO exist [7]. In this respect, it is crucial to restrict the reduction in PPO activity to only the organs and tissues for which it is desirable to inhibit enzymatic browning because PPO may be involved with disease and insect resistance. On the other hand, the development of plant varieties overexpressing PPO that could confer high resistance to diverse diseases and insect pests is a potentially sustainable, environmentally sound, and cost-effective strategy; however, effects on postharvest quality as well as water stress physiology must also be considered. In order to minimize any undesirable effects, suitable regulatory sequences should be evaluated and used for controlling antisense/sense PPO gene expression. The possibility of selectively downregulate PPO expression in specific organs and tissues whereas overexpressing it in others would allow maximal benefits of reduced browning and enhanced pest resistance simultaneously.
As more and more PPO genes are cloned and characterized, wider applications of this technology are expected. The fact that expression of heterologous PPO genes in either antisense/sense orientation has been successfully used to reduce or enhance PPO activity in closely related plant species suggests that PPO genes from one species might be effectively used in other species without the need to isolate every PPO genes from all plant species [58b, 5c, 5d]. This fact should allow faster and wider applications of this technology in the future.
Acknowledgments
We are grateful to Dr. M.D. Hunt and Ms. A. Bhonwong for conducting experiments on Colorado potato beetle, beet army worm and cotton bollworm, respectively. This work was supported by grants from the National Science and Technology Development Agency and Thailand Research Fund, Thailand to P.T.
References and Notes
- Mayer, A. M.; Harel, E. Phenoloxidases and Their Significance in Fruit and Vegetables. In Food Enzymology; Fox P., F., Ed.; Elsevier: New York, 1991; pp. 373–398. [Google Scholar] Friedman, M. Chemistry, Biochemistry, and Dietary Role of Potato Polyphenols. J. Agric. Food Chem. 1997, 45, 1523–1540. [Google Scholar]
- Kojima, M.; Takeuchi, W. Detection and Characterization of p-Coumaric Acid Hydroxylase in Mungbean, Vigna mungo, Seedlings. J. Biochem. 1989, 105, 265–270. [Google Scholar]
- Vaughn, K. C.; Lax, A. R.; Duke, S. O. Polyphenol Oxidase: The Chloroplast Oxidase With No Established Function. Physiol. Plant. 1988, 72, 659–665. [Google Scholar] Trebst, A.; Depka, B. Polyphenol Oxidase and Photosynthesis Research. Photosynth. Res. 1995, 46, 41–44. [Google Scholar]
- Nakayama, T.; Yonekura-Sakakibara, K.; Sato, T.; Kikuchi, S.; Fukui, Y.; Fukuchi-Mizutani, M.; Ueda, T.; Nakao, M.; Tanaka, Y.; Kusumi, T.; Nishino, T. Aureusidin Synthase: A Polyphenol Oxidase Homolog Responsible for Flower Coloration. Science 2000, 290, 1163–1166. [Google Scholar] [CrossRef]
- Felton, G. W.; Donato, K. K.; Del Vecchio, R. J.; Duffey, S. S. Activation of Plant Foliar Oxidases by Insect Feeding Reduces Nutritive Quality of Foliage for Noctuid Herbivores. J. Chem. Ecol. 1989, 15, 2667–2694. [Google Scholar] Stout, M. J.; Workman, K. V.; Bostock, R. M.; Duffey, S. S. Stimulation and Attenuation of Induced Resistance by Elicitors and Inhibitors of Chemical Induction in Tomato (Lycopersicon esculentum) Foliage. Entomol. Exper. Appli. 1998, 86, 267–279. [Google Scholar] Li, L.; Steffens, J. C. Overexpression of Polyphenol Oxidase in Transgenic Tomato Plants Results in Enhanced Bacterial Disease Resistance. Planta 2002, 215, 239–247. [Google Scholar] Thipyapong, P.; Hunt, M. D.; Steffens, J. C. Antisense Downregulation of Polyphenol Oxidase Results in Enhanced Disease Susceptibility. Planta 2004, 220, 105–117. [Google Scholar] Wang, J.; Constabel, C. P. Polyphenol Oxidase Overexpression in Transgenic Populus Enhances Resistance to Herbivory by Forest Tent Caterpillar (Malacosoma disstria). Planta 2004, 220, 87–96. [Google Scholar] Barbehenn, R. V.; Jones, C. P.; Yip, L.; Tran, L.; Constabel, C. P. Does the Induction of Polyphenol Oxidase Defend Trees against Caterpillars? Assessing Plant Defenses One at a Time with Transgenic Poplar. Oecologia 2007, 220. in press. [Google Scholar]
- Mayer, A. M.; Harel, E. Polyphenol Oxidases in Plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] Constabel, C. P.; Bergey, D. R.; Ryan, C. A. Systemin Activates Synthesis of Wound-inducible Tomato Leaf Polyphenol Oxidase via the Octadecanoid Defense Signaling Pathway. Proc. Natl. Acad. Sci. USA 1995, 92, 407–411. [Google Scholar] Thipyapong, P.; Steffens, J. C. Tomato Polyphenol Oxidase: Differential Response of the Polyphenol Oxidase F Promoter to Injuries and Wound Signals. Plant Physiol. 1997, 115, 409–418. [Google Scholar] Maki, H.; Morohashi, Y. Development of Polyphenol Oxidase Activity in the Micropylar Endosperm of Tomato Seeds. J. Plant Physiol. 2006, 163, 1–10. [Google Scholar]
- Steffens, J. C.; Harel, E.; Hunt, M. D. (1994). Polyphenol Oxidase. In Genetic Engineering of Plant Secondary Metabolism; Ellis, B. E., et al., Eds.; Plenum Press: New York, 1994; pp. 275–312. [Google Scholar]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Characterization of the Monophenolase Activity of Tyrosinase on Betaxanthins: The Tyramine-betaxanthin/dopamine-betaxanthin Pair. Planta 2005, 222, 307–318. [Google Scholar] [CrossRef]
- Cary, J. W.; Lax, A. R.; Flurkey, W. H. Cloning and Characterization of cDNAs Coding for Vicia faba Polyphenol Oxidase. Plant Mol. Biol. 1992, 20, 245–253. [Google Scholar] Hunt, M. D.; Eannetta, N. T.; Yu, H.; Newman, S. M.; Steffens, J. C. cDNA Cloning and Expression of Potato Polyphenol Oxidase. Plant Mol. Biol. 1993, 21, 59–68. [Google Scholar] Newman, S. M.; Eannetta, N. T.; Yu, H.; Prince, J. P.; Carmen de Vicente, M.; Tanksley, S. D.; Steffens, J. C. Organisation of the Tomato Polyphenol Oxidase Gene Family. Plant Mol. Biol. 1993, 21, 1035–1051. [Google Scholar] Dry, I. B.; Robinson, S. P. Molecular Cloning and Characterization of Grape Berry Polyphenol Oxidase. Plant Mol. Biol. 1994, 26, 495–502. [Google Scholar] Boss, P. K.; Gardner, R. C.; Janssen, B.-J.; Ross, G. S. An Apple Polyphenol Oxidase cDNA is Up-regulated in Wounded Tissues. Plant Mol. Biol. 1995, 27, 429–433. [Google Scholar] Joy IV, R. W.; Sugiyama, M.; Fukuda, H.; Komamine, A. Cloning and Characterization of Polyphenol Oxidase cDNAs of Phytolacca americana. Plant Physiol 1995, 107, 1083–1089. [Google Scholar] Thygesen, P. W.; Dry, I. B.; Robinson, S. P. Polyphenol Oxidase in Potato: A Multigene Family that Exhibits Differential Expression Patterns. Plant Physiol 1995, 109, 525–531. [Google Scholar] Bucheli, C. S.; Dry, I. B.; Robinson, S. P. Isolation of a Full-length cDNA Encoding Polyphenol Oxidase from Sugarcane, a C4 Grass. Plant Mol. Biol. 1996, 31, 1233–1238. [Google Scholar] Goldman, M. H. S.; Seurinck, J.; Marins, M.; Goldman, G. H.; Mariani, C. A Tobacco Flower-specific Gene Encodes a Polyphenol Oxidase. Plant Mol. Biol. 1998, 36, 479–485. [Google Scholar] Constabel, C. P.; Yip, L.; Patton, J. J.; Christopher, M. E. Polyphenol Oxidase from Hybrid Poplar: Cloning and Expression in Response to Wounding and Herbivory. Plant Physiol. 2000, 124, 285–295. [Google Scholar] Haruta, M.; Pedersen, J. A.; Constabel, C. P. Polyphenol Oxidase and Herbivore Defense in Trembling Aspen (Populus tremuloides): cDNA Cloning, Expression, and Potential Substrates. Plant Physiol. 2001, 112, 552–558. [Google Scholar] Stewart, R. J.; Sawyer, B. J. B.; Bucheli, C. S.; Robinson, S. P. Polyphenol Oxidase is Induced by Chilling and Wounding in Pineapple. Aust. J. Plant Physiol. 2001, 28, 181–191. [Google Scholar] Demeke, T.; Morris, C. F. Molecular Characterization of Wheat Polyphenol Oxidase (PPO). Theor. Appl. Genet. 2002, 104, 813–818. [Google Scholar] Sullivan, M. L.; Hatfield, R. D.; Thoma, S. L.; Samac, D. A. Cloning and Characterization of Red Clover Polyphenol Oxidase cDNAs and Expression of Active Protein in Escherichia coli and Transgenic Alfalfa. Plant Physiol. 2004, 136, 3234–3244. [Google Scholar]
- Thipyapong, P.; Joel, D. M.; Steffens, J. C. Differential Expression and Turnover of the Tomato Polyphenol Oxidase Gene Family during Vegetative and Reproductive Development. Plant Physiol. 1997, 113, 707–718. [Google Scholar] Kim, J. Y.; Seo, Y. S.; Kim, J. E.; Sung, S.-K.; Song, K. J.; An, G.; Kim, W. T. Two Polyphenol Oxidases are Differentially Expressed during Vegetative and Reproductive Development and in Response to Wounding in the Fuji Apple. Plant Sci. 2001, 161, 1145–1152. [Google Scholar]
- Ono, E.; Hatayama, M.; Isono, Y.; Sato, T.; Watanabe, R.; Yonekura-Sakakibara, K.; Fukuchi-Mizutani, M.; Tanaka, Y.; Kusumi, T.; Nishino, T.; Nakayama, T. Localization of a Flavonoid Biosynthetic Polyphenol Oxidase in Vacuoles. Plant J. 2006, 45, 133–143. [Google Scholar] [CrossRef]
- Gooding, P. S.; Bird, C.; Robinson, S. P. Molecular Cloning and Characterisation of Banana Fruit Polyphenol Oxidase. Planta 2001, 213, 748–757. [Google Scholar] Anderson, J. V.; Morris, C. F.; Robinson, S. P. Characterization and Expression of Polyphenol Oxidase in Developing Grain of Wheat (Triticum aestivum L.). In Plant Biology 2003; Honolulu, Hawaii, USA, 2003. [Google Scholar]
- Thipyapong, P.; Melkonian, J.; Wolfe, D. W.; Steffens, J. C. Suppression of Polyphenol Oxidases Increases Stress Tolerance in Tomato. Plant Sci. 2004, 167, 693–703. [Google Scholar] [CrossRef]
- Hind, G.; Marshak, D. R.; Coughlan, S. J. Spinach Thylakoid Polyphenol Oxidase: Cloning, Characterization, and Relation to a Putative Protein Kinas. Biochemistry 1995, 34, 8157–8164. [Google Scholar] [CrossRef]
- Haruta, M.; Murata, M.; Kadokura, H.; Homma, S. Immunological and Molecular Comparison of Polyphenol Oxidase in Rosaceae Fruit Trees. Phytochemistry 1999, 50, 1021–1025. [Google Scholar] [CrossRef]
- Nishimura, M.; Fukuda, C.; Murata, M.; Homma, S. Cloning and Some Properties of Japanese Pear (Pyrus pyrifolia) Polyphenol Oxidase, and Changes in Browning Potential during Fruit Maturation. J. Sci. Food Agric. 2003, 83, 1156–1162. [Google Scholar] [CrossRef]
- Shahar, T.; Hennig, N.; Gutfinger, T.; Hareven, D.; Lifschitz, E. The Tomato 66.3-kD Polyphenoloxidase Gene: Molecular Identification and Developmental Expression. Plant Cell 1992, 4, 135–147. [Google Scholar] Thipyapong, P.; Hunt, M. D.; Steffens, J. C. Systemic Wound Induction of Potato (Solanum tuberosum) Polyphenol Oxidase. Plant Cell 1995, 40, 673–676. [Google Scholar] Chevalier, T.; de Rigal, D.; Mbéguié-A-Mbéguié, D.; Gauillard, F.; Richard-Forget, F.; Fils-Lycaon, B. R. Molecular Cloning and Characterization of Apricot Fruit Polyphenol Oxidase. Plant Physiol. 1999, 119, 1261–1269. [Google Scholar]
- Gleave, A. P. A Versatile Binary Vector System With a T-DNA Organisational Structure Conducive to Efficient Integration of Cloned DNA into the Plant Genome. Plant Mol. Biol. 1992, 20, 1203–1207. [Google Scholar] [CrossRef]
- Thipyapong, P. Polyphenol Oxidase Gene Family: Differential Expression during Vegetative and Reproductive Development, and in Response to Injuries, and Defensive Functional Analysis. Ph.D. Thesis, Cornell University, Ithaca, New York, 1997. [Google Scholar]
- Levine, A.; Pennell, R. I.; Alvarez, M. E.; Palmer, R.; Lamb, C. Calcium-mediated Apoptosis in a Plant Hypersensitive Disease Resistance Response. Current Biol. 1996, 6, 427–437. [Google Scholar] Chamnongpol, S.; Willekens, H.; Moeder, W.; Langebartels, C.; Sandermann, H., Jr.; Montagu, M. V.; Inzé, D.; Camp, W. V. Defense Activation and Enhanced Pathogen Tolerance Induced by H2O2 in Transgenic Tobacco. Proc. Natl. Acad. Sci. USA 1998, 95, 5818–5823. [Google Scholar] Grant, J. J.; Loake, G. J. Role of Reactive Oxygen Intermediates and Cognate Redox Signaling in Disease Resistance. Plant Physiol. 2000, 124, 21–29. [Google Scholar] Garcia-Olmedo, F.; Rodriguez-Palenzuela, P.; Molina, A.; Alamillo, J. M.; Lopez-Solanilla, E.; Berrocal-Lobo, M.; Poza-Carrion, C. Antibiotic Activities of Peptides, Hydrogen Peroxide and Peroxynitrite in Plant Defence. FEBS Letters 2001, 498, 219–222. [Google Scholar]
- Campos, A. D.; Ferreira, A. G.; Vozarí Hampe, M. M.; Antunes, I. F.; Brancão, N.; da Silveira, E. P.; Osório, V. A.; Augustin, E. Peroxidase and Polyphenol Oxidase Activity in Bean Anthracnose Resistance. Pesq. agropec. bras., Brasilia. 2004, 39, 637–643. [Google Scholar]
- Tyagi, M.; Kayastha, A. M.; Sinha, B. The Role of Peroxidase and Polyphenol Oxidase Isozymes in Wheat Resistance to Alternaria triticina. Biol. Plant. 2000, 43, 559–562. [Google Scholar] Raj, S. N.; Sarosh, B. R.; Shetty, H. S. Induction and Accumulation of Polyphenol Oxidase Activities as Implicated in Development of Resistance Against Pearl Millet Downy Mildew Disease. Funct. Plant Biol. 2006, 33, 563–571. [Google Scholar]
- Krishnamoorthy, V.; Kumar, N.; Angappan, K.; Soorianathasundaram, K. Evaluation of New Banana Hybrids Against Black Leaf Streak Disease. Infomusa 2004, 13, 25–27. [Google Scholar]
- Banga, S. S.; Kaur, K.; Ahuja, K. L.; Banga, S. K. Introgression and Biochemical Manifestation of the Gene(s) for White Rust Resistance in Indian Mustard (Brassicajuncea (L.) Coss.) [online]. 2004. available from: http://www.cropscience.org.au/icsc2004/poster/3/7/3/296_banga.htm.
- Tomiyama, K.; Stahmann, M. A. Alteration of Oxidative Enzymes in Potato Tuber Tissue by Infection with Phytophthora infestans. Plant Physiol. 1964, 39, 483–490. [Google Scholar] [CrossRef]
- Mohammadi, M.; Kazemi, H. Changes in Peroxidase and Polyphenol Oxidase Activities in Susceptible and Resistant Wheat Heads Inoculated with Fusarium graminearum and Induced Resistance. Plant Sci. 2002, 162, 491–498. [Google Scholar] Honty, K.; Hevesi, M.; Tóth, M.; Stefanovíts-Bányai, É. Some Biochemical Changes in Pear Fruit Tissue Induced by Erwinia aymlovora. Acta Biol. Szeged 2005, 49, 127–129. [Google Scholar]
- Stahmann, M. A.; Clare, B. G.; Woodbury, W. Increase Disease Resistance and Enzyme Activity Induced by Ethylene and Ethylene Production by Black Rot Infected Sweet Potato Tissue. Plant Physiol. 1966, 41, 1505–1512. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y. C.; Sun, W. C.; Si, J.; Römheld, V. Effects of Foliar- and Root-applied Silicon on the Enhancement of Induced Resistance to Powdery Mildew in Cucumis sativus. Plant Pathol. 2005, 54, 678–685. [Google Scholar] [CrossRef]
- Mathur, N.; Vyas, A. Biochemical Changes in Ziziphus xylopyrus by VA Mycorrhizae. Bot. Bull. Acad. Sin. 1996, 37, 209–212. [Google Scholar] Panwar, J.; Vyas, A. AM Fungi: A Biological Approach Towards Conservation of Endangered Plants in Thar Desert, India. Curr. Sci. 2002, 82, 576–578. [Google Scholar]
- Kavitha, P. G.; Varadarajan, R.; Jonathan, E. I. Induced Systemic Resistance of Pseudomonas fluorescens Against Root-knot Nematode, Meloidogyne incognita in Tomato. Resistant Pest Management Newsletter n.d., [ http://whalonlab.msu.edu/rpmnews/vol.15_no.2/globe/Kavitha_etal.htm], 15.
- Gailite, A.; Samsone, I.; Ievinsh, G. Ethylene is Involved in Trichoderma-induced Resistance of Bean Plants Against Pseudomonas syringae. Acta Universitatis Latviensis, Biology 2005, 691, 59–70. [Google Scholar]
- Wegener, C. B.; Olsen, O. Heterologous Pectate Lyase Isoenzymes are Not Different in Their Effects on Soft Rot Resistance in Transgenic Potatoes. Physiol. Mol. Plant Pathol. 2004, 65, 59–66. [Google Scholar] [CrossRef]
- Punja, Z. K. Genetic Engineering of Plants to Enhance Resistance to Fungal Pathogens-A Review of Progress and Future Prospects. Can. J. Plant Pathol. 2001, 23, 216–235. [Google Scholar]
- Castañera, P.; Steffens, J. C.; Tingey, W. M. Biological Performance of Colorado Potato Beetle Larvae on Potato Genotypes with Differing Levels of Polyphenol Oxidase. J. Chem. Ecol. 1996, 22, 91–101. [Google Scholar] Ramiro, D. A.; Guerreiro-Filho, O.; Mazzafera, P. Phenol Contents, Oxidase Activities, and the Resistance of Coffee to the Leaf Miner Leucoptera coffeella. J. Chem. Ecol. 2006, 32, 1997–1988. [Google Scholar]
- Duffey, S. S.; Stout, M. J. Antinutritive and Toxic Components of Plant Defense Against Insects. Arch. Insect Biochem. Physiol. 1996, 32, 3–37. [Google Scholar] [CrossRef]
- Thaler, J. S.; Karban, R.; Ullman, D. E.; Boege, K.; Bostock, R. M. Cross-talk Between Jasmonate and Salicylate Plant Defense Pathways: Effects on Several Plant Parasites. Oecologia 2002, 131, 227–235. [Google Scholar] Cooper, W. C.; Jia, L.; Goggin, F. L. Acquired and R-gene-mediated Resistance Against the Potato Aphid in Tomato. J. Chem. Ecol. 2004, 30, 2527–2542. [Google Scholar]
- Ren, F.; Lu, Y.-T. Overexpression of Tobacco Hydroxyproline-rich Glycopeptide Systemin Precursor A Gene in Transgenic Tobacco Enhances Resistance Against Helicoverpa armigera Larvae. Plant Sci. 2006, 171, 286–292. [Google Scholar] [CrossRef]
- Li, L.; Zhao, Y.; McCaig, B. C.; Wingerd, B. A.; Wang, J.; Whalon, M. E.; Pichersky, E.; Howe, G. A. The Tomato Homolog of Coronatine-insensitive1 is Required for the Maternal Control of Seed Maturation, Jasmonate-signaled Defense Responses, and Glandular Trichome Developmen. Plant Cell 2004, 16, 126–143. [Google Scholar] [CrossRef]
- Felton, G. W.; Donato, K. K.; Broadway, R. M.; Duffey, S. S. Impact of Oxidized Plant Phenolics on the Nutritional Quality of Dietary Protein to a Noctuid Herbivore, Spodoptera exigua. J. Insect Physiol. 1992, 38, 277–285. [Google Scholar] [CrossRef]
- Ananthakrishnan, T. N. Applied Chemical Ecology: Induced and Transgenic Defences in Insect-plant Interactions. Current Sci. 2003, 84, 492–493. [Google Scholar]
- Hoover, K.; Stout, M. J.; Alaniz, S. A.; Hammock, B. D.; Duffey, S. S. Influence of Induced Plant Defenses in Cotton and Tomato on the Efficacy of Baculoviruses on Noctuid Larvae. J. Chem. Ecol. 1998, 24, 253–271. [Google Scholar]
- Tolbert, N. E. Activation of Polyphenol Oxidase of Chloroplasts. Plant Physiol. 1973, 51, 234–244. [Google Scholar] Vaughn, K. C.; Duke, S. O. Function of Polyphenol Oxidase in Higher Plants. Physiol. Plant. 1984, 60, 106–112. [Google Scholar]
- Badger, M. R.; von Caemmerer, S.; Ruuska, S.; Nakano, H. Electron Flow to Oxygen in Higher Plants and Algae: Rates and Control of Direct Photoreduction (Mehler Reaction) and Rubisco Oxygenase. Phil. Trans. R. Soc. Lond. B. 2000, 355, 1433–1446. [Google Scholar] Haupt-Herting, S.; Fock, H. P. Oxygen Exchange in Relation to Carbon Assimilation in Water-stressed Leaves during Photosynthesis. Annals Bot. 2002, 89, 851–859. [Google Scholar]
- Asada, K.; Takahashi, M. Production and Scavenging of Active Oxygen in Chloroplasts. In Photoinhibition; Kyle, D. J., Osmond, C. B., Arntzen, C. J., Eds.; Elsevier: Amsterdam, 1987; pp. 227–287. [Google Scholar]
- Demmig, B.; Bjorkman, O. Comparison of the Effect of Excessive Light on Chlorophyll Fluorescence (77 K) and Photon Yield of O2 Evolution in Leaves of Higher Plants. Planta 1987, 171, 171–184. [Google Scholar] [CrossRef]
- English-Loeb, G.; Stout, M. J.; Duffey, S. S. Drought Stress in Tomatoes: Changes in Plant Chemistry and Potential Nonlinear Consequences for Insect Herbivores. OIKOS 1997, 79, 456–468. [Google Scholar] [CrossRef]
- Shivishankar, S. Polyphenol Oxidase Isozymes in Coconut Genotypes under Water Stress. Plant Physiol. Biochem. 1988, 15, 87–91. [Google Scholar]
- Niknam, V.; Razavi, N.; Ebrahimzadeh, H.; Sharifizadeh, B. Effect of NaCl on Biomass, Protein and Proline Contents, and Antioxidant Enzymes in Seedlings and Calli of Two Trigonella Species. Biol. Plant. 2006, 50, 591–596. [Google Scholar] [CrossRef]
- Balakumar, T.; Gayathri, B.; Anbudurai, P. R. Oxidative Stress Injury in Tomato Plants Induced by Supplemental UV-B Radiation. Biol. Plant. 1997, 39, 215–221. [Google Scholar] [CrossRef]
- Sherman, T. D.; Vaughn, K. C.; Duke, S. O. A Limited Survey of the Phylogenetic Distribution of Polyphenol Oxidase. Phytochemistry 1991, 30, 2499–2506. [Google Scholar] [CrossRef]
- Cho, M.-H.; Moinuddin, S. G. A.; Helms, G. L.; Hishiyama, S.; Eichinger, D.; Davin, L. B.; Lewis, N. G. (+)-Larreatricin Hydroxylase, an Enantio-specific Polyphenol Oxidase from the Creosote Bush (Larrea tridentata). Proc. Natl. Acad. Sci. USA 2003, 100, 10641–10646. [Google Scholar] [CrossRef]
- Vaughn, K. C.; Duke, S. O. Tissue Localization of Polyphenol Oxidase in Sorghum. Protoplasma 1981, 108, 319–327. [Google Scholar] Vaughn, K. C.; Duke, S. O. Tentoxin Effects on Sorghum: The Role of Polyphenol Oxidase. Protoplasma 1982, 110, 48–53. [Google Scholar]
- Franke, R.; Humphreys, J. M.; Hemm, M. R.; Denault, J. W.; Ruegger, M. O.; Cusumano, J. C.; Chapple, C. The Arabidopsis REF8 Gene Encodes the 3-Hydroxylase of Phenylpropanoid Metabolism. Plant J. 2002, 30, 33–45. [Google Scholar] [CrossRef]
- Sommer, A.; Néeman, E.; Steffens, J. C.; Mayer, A. M.; Harel, E. Import, Targeting and Processing of a Plant Polyphenol Oxidase. Plant Physiol. 1994, 105, 1301–1311. [Google Scholar]
- Sullivan, M. L.; Hatfield, R. D. Polyphenol Oxidase and o-Diphenols Inhibit Postharvest Proteolysis in Red Clover and Alfalfa. Crop Sci. 2006, 46, 662–670. [Google Scholar] [CrossRef]
- Ebrahimzadeh, H.; Abrishamchi, P. Changes in IAA, Phenolic Compounds, Peroxidase, IAA Oxidase, and Polyphenol Oxidase in Relation to Flower Formation in Crocus sativus. Russian J. Plant Physiol. 2001, 48, 190–195. [Google Scholar] Lavid, N.; Schwartz, A.; Lewinsohn, E.; Tel-Or, E. Phenols and Phenol Oxidases are Involved in Cadmium Accumulation in the Water Plants Nymphoides peltata (Menyanthaceae) and Nymphaeae (Nymphaeaceae). Planta 2001, 214, 189–195. [Google Scholar]
- Spagna, G.; Barbagallo, R. N.; Chisari, M.; Branca, F. Characterization of a Tomato Polyphenol Oxidase and Its Role in Browning and Lycopene Content. J. Agric. Food Chem. 2005, 53, 2032–2038. [Google Scholar] [CrossRef]
- Bachem, C. W. B.; Speckmann, G. J.; van der Linde, P. C. G.; Verheggen, F. T. M.; Hunt, M. D.; Steffens, J. C.; Zabeau, M. Antisense Expression of Polyphenol Oxidase Genes Inhibits Enzymatic Browning in Potato Tubers. Biotechnology 1994, 12, 1101–1105. [Google Scholar] Coetzer, C.; Corsini, D.; Love, S.; Pavek, J.; Tumer, N. Control of Enzymatic Browning in Potato (Solanum tuberosum L.) by Sense and Antisense RNA from Tomato Polyphenol Oxidase. J. Agri. Food Chem. 2001, 49, 652–657. [Google Scholar]
- O’Neill, G. New Greens Beat the Browning Blues. ECOS. 1994. [ http://www.publish.csiro.au/?act=view_file&file_id=EC82p12.pdf], 82. (b)CSIRO. Field Trial of GM Grapevines - Evaluation of Berry Colour, Sugar Composition, Flower and Fruit Development and Pollen Flow Study [online]. 2003. available from: http://www.ogtr.gov.au/rtf/ir/dir031finalrarmp.rtf. [Google Scholar]
- Queensland Department of Primary Industries. Field Trial of Pineapple Plants Modified for Blackheart Reduction and to Delay Flowering [online]. 2003. available from: http://www.ogtr.gov.au/rtf/ir/dir028finalrarmp2.rtf. [Google Scholar]
- Murata, M.; Haruta, M.; Murai, N.; Tanikawa, N.; Nishimura, M.; Homma, S.; Itoh, Y. Transgenic Apple (Malus × domestica) Shoot Showing Low Browning Potential. J. Agric. Food Chem. 2000, 48, 5243–5248. [Google Scholar] Cao, Y. H.; Zhang, Z.; Yao, Q. H.; Peng, R. H.; Xiong, A. S.; Li, X. Suppression of Apple Polyphenol Oxidase by Double-stranded RNA (RNAi). Acta Biol. Exper. Sinica 2004, 37, 487–493. (in Chinese). [Google Scholar] Okanagan Biotechnology Inc. Products & Services: Apple Varieties/ The Non-browning Apple [online]. 2005. available from: http://www.okanaganbiotechnology.com/apple-non-browning.php.
- Vickers, J. E.; Grof, C. P. L.; Bonnett, G. D.; Jackson, P. A.; Knight, D. P.; Roberts, S. E.; Robinson, S. P. Overexpression of Polyphenol Oxidase in Transgenic Sugarcane Results in Darker Juice and Raw Sugar. Crop Sci. 2005, 45, 354–362. [Google Scholar] [CrossRef]
- Robinson, S. P.; Loveys, B. R.; Chacko, E. K. Polyphenol Oxidase Enzymes in the Sap and Skin of Mango Fruit. Aust. J. Plant Physiol. 1993, 20, 99–107. [Google Scholar]
- Okanagan Biotechnology Inc. Products & Services: Cherry Varieties/ PPO Inhibited Cherry [online]. 2005. available from: http://www.okanaganbiotechnology.com/cherry-ppo-inhibited.php.
- Haruta, M.; Murata, M.; Hiraide, A.; Kadokura, H.; Yamasaki, M.; Sakuta, M.; Shimizu, S.; Homma, S. Cloning Genomic DNA Encoding Apple Polyphenol Oxidase and Comparison of the Gene Product in Escherichia coli and in Apple. Biosci. Biotechnol. Biochem. 1998, 62, 358–362. [Google Scholar] [CrossRef]
- Robinson, S. P.; Dry, I. B. Broad Bean Leaf Polyphenol Oxidase is a 60-kilodalton Protein Susceptible to Proteolytic Cleavage. Plant Physiol. 1992, 99, 317–323. [Google Scholar] [CrossRef]
- Greving, J.; Gerdemann, C.; Spener, F.; Krebs, B. Sequencing and Cloning of Genomic DNA Encoding Ipomoea batatas Catechol Oxidase, NCBI#AJ309175 [online]. 2001. [01/04/09], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nucleotide&val=29691901, http://www.ebi.uniprot.org/uniprot-srv/results/gridView.do;jsessionid=C3912C67B...
- Rathjen, A. H.; Robinson, S. P. Aberrant Processing of Polyphenol Oxidase in a Variegated Grapevine Mutant. Plant Physiol. 1992, 99, 1619–1625. [Google Scholar] [CrossRef]
- (a)Pan, D.; Chen, G.; Lai, Z.; Lv, L. Molecular Cloning of Full Length cDNA Encoding Polyphenol Oxidase in Pulp of Nai (Prunus salicina Lindl. var. cordata Y. He & J. Y. Zhang), NCBI#AAW58109, AAW59443, AY865623, AY870335 [online]. 2005; [05/08/03], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=71738563, http:// www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nucleotide&val=71738562. [Google Scholar] (b)Pan, D.; Chen, G.; Lai, Z.; Lv, L.; Peng, S.; Lai, Z.; Guo, Z. Isolation of Full Length Nai (Prunus salicina Lindl.) Leaf Polyphenol Oxidase, NCBI AAW65103 [online]. 2005. available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=58047496. [Google Scholar]
- Robinson, S. P. Polyphenol Oxidase Genes from Banana, Lettuce, Tobacco and Pineapple. US Pat. 2005. available from: http://www.freshpatents.com/Polyphenol-oxidase-genes-from-banana,-lettuce,-tobacco-and-pineapple-dt20050901ptan20050191739.php.
- Hind, G.; Marshak, D. R.; Coughlan, S. J. Spinach Thylakoid Polyphenol Oxidase: Cloning, Characterization, and Relation to a Putative Protein Kinase. Biochemistry 1995, 34, 8157–8164. [Google Scholar] Sokolenko, A.; Fulgosi, H.; Gal, A.; Altschmied, L.; Ohad, I.; Herrmann, R. G. The 64 kDa Polypeptide of Spinach may Not be the LHCII Kinase, But a Lumen-located Polyphenol Oxidase. FEBS Letters 1995, 371, 176–180. [Google Scholar]
- Jukanti, A. K.; Bruckner, P. L.; Fischer, A. M. Evaluation of Wheat Polyphenol Oxidase Genes. Cereal Chem. 2004, 81, 481–485. [Google Scholar] Raman, R.; Raman, H.; Johnstone, K.; Lisle, C.; Smith, A.; Matin, P.; Allen, H. Genetic and in Silico Comparative Mapping of the Polyphenol Oxidase Gene in Bread Wheat (Triticum aestivum L.). Funct. Integr. Genomics 2005, 5, 185–200. [Google Scholar]
- Sullivan, M. L.; Evans, M.; Thoma, S. L.; Samac, D. A.; Hatfield, R. D. Alfalfa Polyphenol Oxidase is Expressed in Flowers and Seed Pods, NCBI#AAP33165 [online]. 2003. [03/05/15], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=30790423. [Google Scholar]
- (a)Raizada, J.; Kumar, S.; Ahuja, P. S. Cloning and Characterization of Polyphenol Oxidase Gene in Camellia sinensis (L.) O. kuntze, NCBI#AAT75166 [online]. 2004. [04/08/20], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=50346969. [Google Scholar] (b)Li, B.; Chen, Z. Z.; Li, Q. H. Cloning Polyphenol Oxidase in Tea Variety Camellia ptilophylla, NCBI#ABF19601 [online]. 2006. [06/05/06], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=94450151. [Google Scholar] (c)Li, B.; Chen, Z. Z.; Li, Q. H. Cloning Polyphenol Oxidase in Tea Variety Camellia sinensis var. assamica cv. Yinghong 9, NCBI#ABF19602 [online]. 2006. [06/05/06], available from: http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=protein&val=94450153. [Google Scholar]
- Sample Availability: Available from the authors.
© 2007 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
MDPI and ACS Style
Thipyapong, P.; Stout, M.J.; Attajarusit, J. Functional Analysis of Polyphenol Oxidases by Antisense/Sense Technology. Molecules 2007, 12, 1569-1595. https://doi.org/10.3390/12081569
AMA Style
Thipyapong P, Stout MJ, Attajarusit J. Functional Analysis of Polyphenol Oxidases by Antisense/Sense Technology. Molecules. 2007; 12(8):1569-1595. https://doi.org/10.3390/12081569
Chicago/Turabian StyleThipyapong, Piyada, Michael J. Stout, and Jutharat Attajarusit. 2007. "Functional Analysis of Polyphenol Oxidases by Antisense/Sense Technology" Molecules 12, no. 8: 1569-1595. https://doi.org/10.3390/12081569