Ferroquine, an Ingenious Antimalarial Drug –Thoughts on the Mechanism of Action

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Introduction

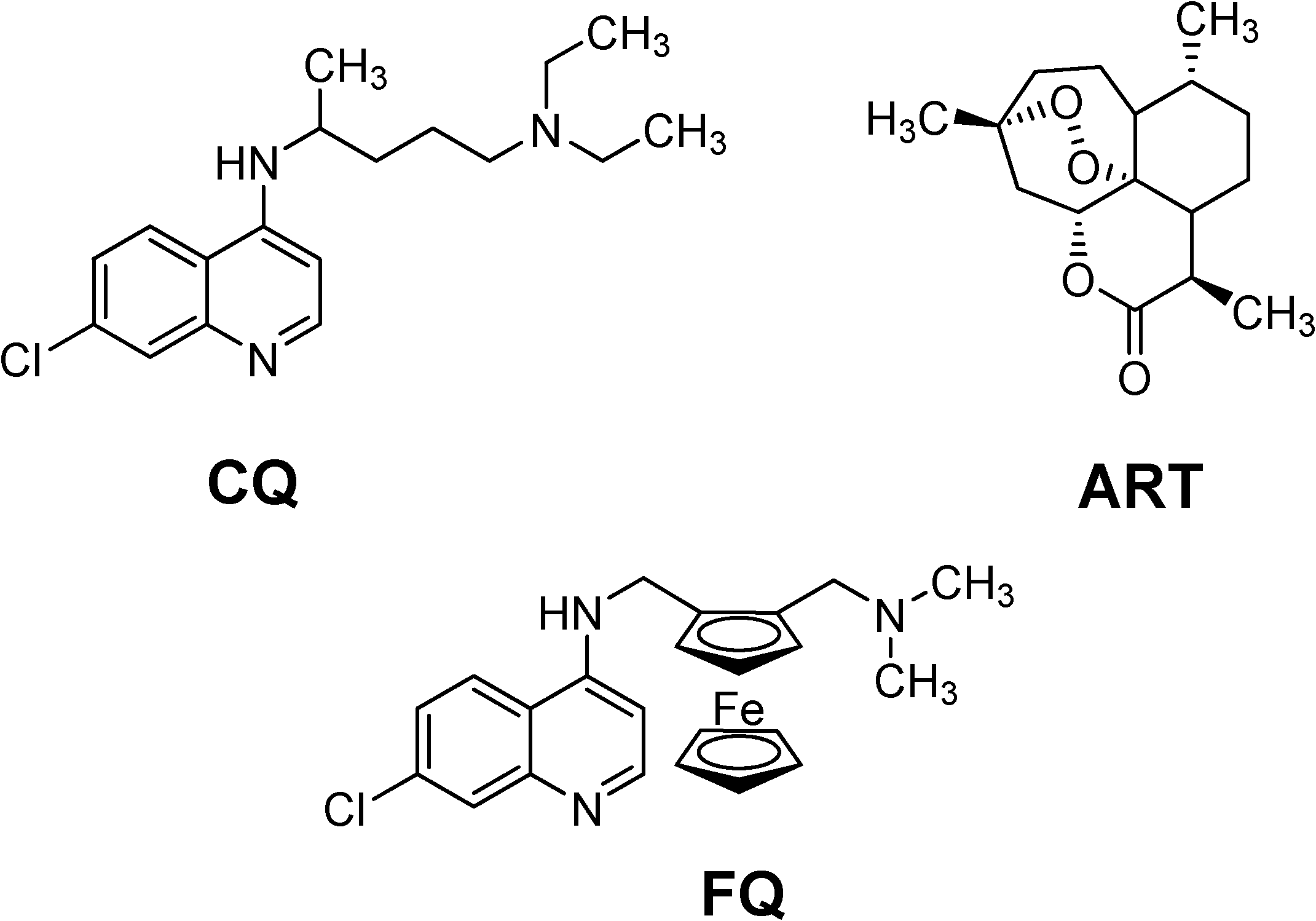
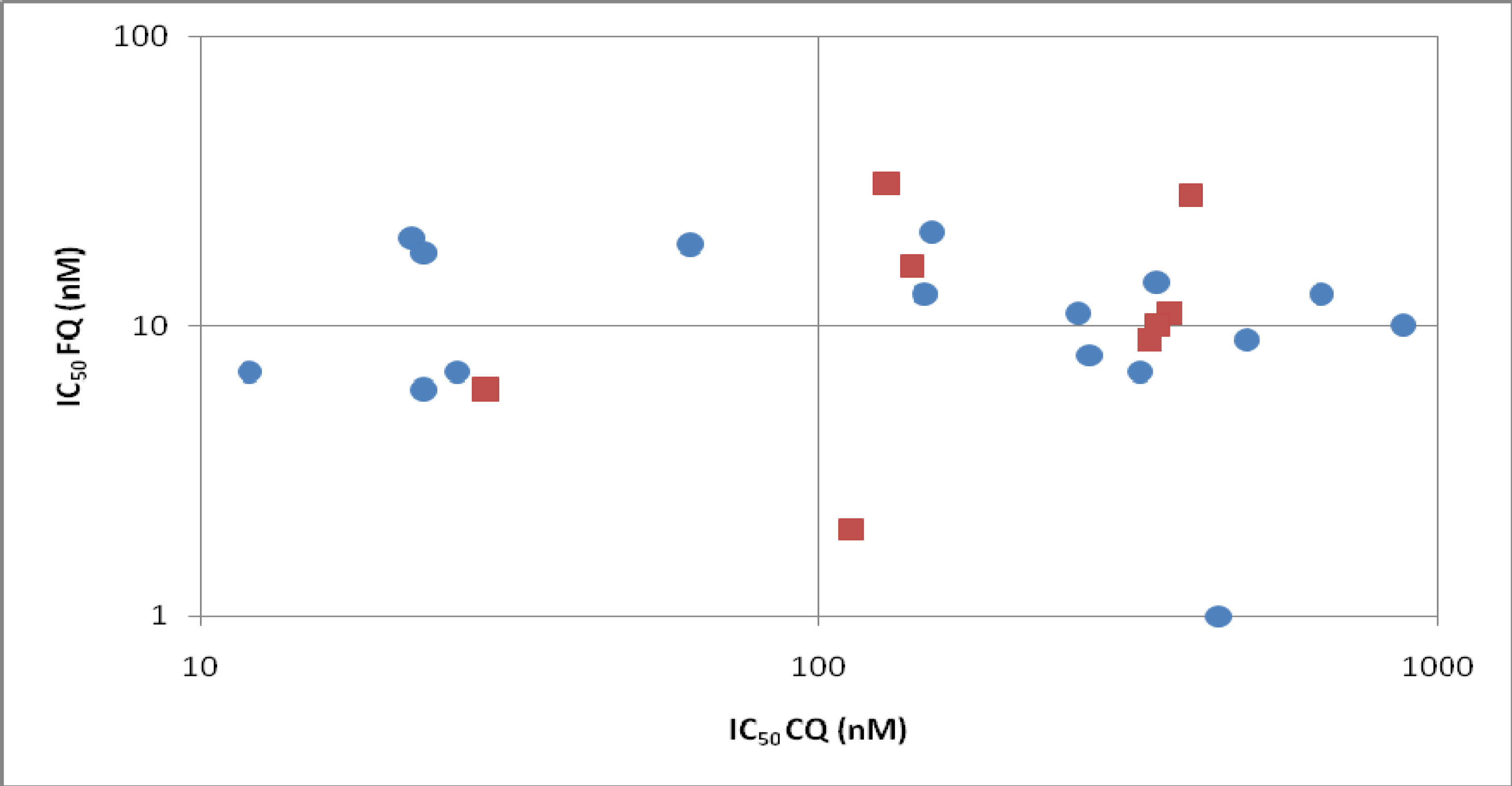
Specific pharmacology
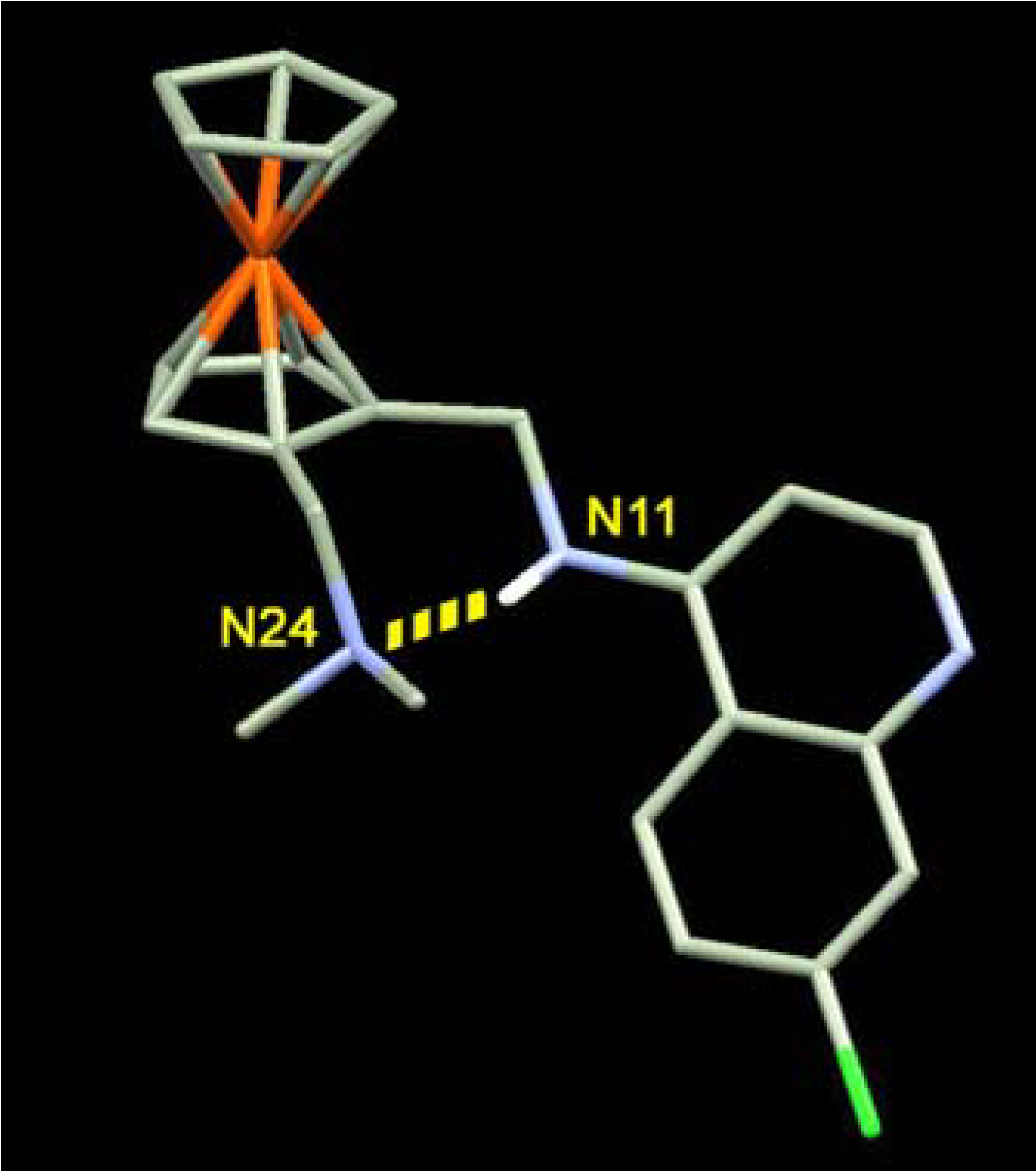
Mechanism of action

Conclusions
Acknowledgements
References
- World Health Organization. Available online: http://www.who.int.
- Le Bras, J.; Musset, L.; Clain, J. Antimalarial drug resistance. Med. Mal. Infect. 2006, 36, 401–405. [Google Scholar] [CrossRef]
- Tang, Y.; Dong, Y.; Vennerstrom, J.L. Synthetic peroxides as antimalarials. Med. Res. Rev. 2004, 24, 425–448. [Google Scholar] [CrossRef]
- Mutabingwa, T.K. Artemisinin-based combination therapies (ACTs): Best hope for malaria treatment but inaccessible to the needy. Acta. Trop. 2005, 95, 305–315. [Google Scholar]
- Yeung, S.; Van Damme, W.; Socheat, D.; White, N.J.; Mills, A. Cost of increasing access to artemisinin combination therapy: the Cambodian experience. Malar. J. 2008, 7, 84. [Google Scholar] [CrossRef]
- Dinglasan, R.R.; Jacobs-Lorena, M. Flipping the paradigm on malaria transmission-blocking vaccines. Trends Parasitol. 2008, 24, 364–370. [Google Scholar] [CrossRef]
- Top, S.; Vessières, A.; Leclercq, G.; Quivy, J.; Tang, J.; Vaissermann, J.; Huché, M.; Jaouen, G. Synthesis, biochemical properties and molecular modelling studies of organometallic specific estrogen receptor modulators (SERMs), the ferrocifens and hydroxyferrocifens: evidence for an antiproliferative effect of hydroxyferrocifens on both hormone-dependent and hormone-independent breast cancer cell lines. Chem. Eur. J. 2003, 9, 5223–5236. [Google Scholar] [CrossRef]
- Vessières, A.; Top, S.; Pigeon, P.; Hillard, E.A.; Boubeker, L.; Spera, D.; Jaouen, G. Modification of the estrogenic properties of diphenols by the incorporation of ferrocene. Generation of antiproliferative effects in vitro. J. Med. Chem. 2005, 48, 3937–3940. [Google Scholar] [CrossRef]
- Dive, D.; Biot, C. Ferrocene conjugates of chloroquine and other antimalarials: the development of ferroquine, a new antimalarial. ChemMedChem 2008, 3, 383–391. [Google Scholar] [CrossRef]
- Blackie, M.A.; Beagley, P.; Croft, S.L; Kendrick, H.; Moss, J.R.; Chibale, K. Metallocene-based antimalarials: an exploration into the influence of the ferrocenyl moiety on in vitro antimalarial activity in chloroquine-sensitive and chloroquine-resistant strains of Plasmodium falciparum. Bioorg. Med. Chem. 2007, 15, 6510–6516. [Google Scholar] [CrossRef]
- Blackie, M.A.; Chibale, K. Metallocene antimalarials: the continuing quest. Met. Based Drugs 2008, 495123. [Google Scholar]
- Available online: http://clinicaltrialsfeeds.org/clinical-trials/show/NCT00563914.
- Biot, C.; Daher, W.; Ndiaye, C.M.; Melnyk, P.; Pradines, B.; Chavain, N.; Pellet, A.; Fraisse, L.; Pelinski, L.; Jarry, C.; Brocard, J.; Khalife, J.; Forfar-Bares, I.; Dive, D. Probing the role of the covalent linkage of ferrocene into a chloroquine template. J. Med. Chem. 2006, 49, 4707–4714. [Google Scholar] [CrossRef]
- Beagley, P.; Blackie, M.A.L.; Chibale, K.; Clarkson, C.; Meijboom, R.; Moss, J.R.; Smith, P.J.; Su, H. Synthesis and antiplasmodial activity in vitro of new ferrocene chloroquine-analogues. Dalton Trans. 2003, 3046–3051. [Google Scholar]
- Biot, C.; Daher, W.; Chavain, N.; Fandeur, T.; Khalife, J.; Dive, D.; De Clercq, E. Design and synthesis of hydroxyferroquine derivatives with antimalarial and antiviral activities. J Med. Chem. 2006, 49, 2845–2849. [Google Scholar] [CrossRef] [Green Version]
- Delhaes, L.; Abessolo, H.; Biot, C.; Berry, L.; Delcourt, P.; Maciejewski, L.; Brocard, J.; Camus, D.; Dive, D. In vitro and in vivo antimalarial activity of ferrochloroquine, a ferrocenyl analogue of chloroquine against chloroquine-resistant malaria parasites. Parasitol. Res. 2001, 87, 239–244. [Google Scholar] [CrossRef]
- Biot, C.; Chavain, N.; Dubar, F.; Pradines, B.; Trivelli, X.; Brocard, J.; Forfar, I.; Dive, D. Structure-activity relationships of 4-N-substituted ferroquine analogues. Time to re-evaluate the mechanism of action of ferroquine. J. Organomet. Chem. 2008, in press. [Google Scholar]
- Atteke, C.; Ndong, J.M.; Aubouy, A.; Maciejewski, L.; Brocard, J.; Lebibi, J.; Deloron, P. In vitro susceptibility to a new antimalarial organometallic analogue, ferroquine, of Plasmodium falciparum isolates from the Haut-Ogooué region of Gabon. J. Antimicrob. Chemother. 2003, 51, 1021–1024. [Google Scholar] [CrossRef]
- Pradines, B.; Tall, A.; Rogier, C.; Spiegel, A.; Mosnier, J.; Marrama, L.; Fusai, T.; Millet, P.; Panconi, E.; Trape, J.F.; Parzy, D. In vitro activities of ferrochloroquine against 55 Senegalese isolates of Plasmodium falciparum in comparison with those of standard antimalarial drugs. Trop. Med. Int. Health 2002, 7, 265–270. [Google Scholar] [CrossRef]
- Pradines, B.; Fusai, T.; Daries, W.; Laloge, V.; Rogier, C.; Millet, P.; Panconi, E.; Kombila, M.; Parzy, D. Ferrocene-chloroquine analogues as antimalarial agents: in vitro activity of ferrochloroquine against 103 Gabonese isolates of Plasmodium falciparum. J. Antimicrob. Chemother. 2001, 48, 179–184. [Google Scholar] [CrossRef]
- Chim, P.; Lim, P.; Sem, R.; Nhem, S.; Maciejewski, L.; Fandeur, T. The in-vitro antimalarial activity of ferrochloroquine, measured against Cambodian isolates of Plasmodium falciparum. Ann. Trop. Med. Parasitol. 2004, 98, 419–424. [Google Scholar] [CrossRef]
- Kreidenweiss, A.; Kremsner, P.G.; Dietz, K.; Mordmüller, B. In vitro activity of ferroquine (SAR97193) is independent of chloroquine resistance in Plasmodium falciparum. Am. J. Trop. Med. Hyg. 2006, 75, 1178–1181. [Google Scholar]
- Barends, M.; Jaidee, A.; Khaohirun, N.; Singhasivanon, P.; Nosten, F. In vitro activity of ferroquine (SSR 97193) against Plasmodium falciparum isolates from the Thai-Burmese border. Malar. J. 2007, 6, 81. [Google Scholar] [CrossRef]
- Fraisse, L.; Ter-minassian, D. International Application Association between ferroquine and an artemisinine derivative for treating malaria. PCT/FR2006/000842, 2006. [Google Scholar]
- Long, T.T.; Nakazawa, S.; Onizuka, S.; Huaman, M.C.; Kanbara, H. Influence of CD4+CD25+ T cells on Plasmodium berghei NK65 infection in BALB/c mice. Int. J. Parasitol. 2003, 33, 175–183. [Google Scholar] [CrossRef]
- Pierrot, C.; Lafitte, S.; Dive, D.; Fraisse, L.; Brocard, J.; Khalife, J. Analysis of immune response patterns in naïve and Plasmodium berghei-infected young rats following a ferroquine treatment. Int. J. Parasitol. 2005, 35, 1601–1610. [Google Scholar] [CrossRef]
- Henry, M.; Briolant, S.; Fontaine, A.; Mosnier, J.; Baret, E.; Amalvict, R.; Fusaï, T.; Fraisse, L.; Rogier, C.; Pradines, B. In Vitro Activity of Ferroquine is Independent of Polymorphisms in Transport Proteins Genes Implicated in Quinoline Resistance in Plasmodium falciparum. Antimicrob. Agents Chemother. 2008, 52, 2755–2759. [Google Scholar] [CrossRef]
- Daher, W.; Biot, C.; Fandeur, T.; Jouin, H.; Pelinski, L.; Viscogliosi, E.; Fraisse, L.; Pradines, B.; Brocard, J.; Khalife, J.; Dive, D. Assessment of P. falciparum resistance to ferroquine in field isolates and in W2 strain under pressure. Malaria J. 2006, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Biot, C.; Taramelli, D.; Forfar-Bares, I.; Maciejewski, L.A.; Boyce, M.; Nowogrocki, G.; Brocard, J.S.; Basilico, N.; Olliaro, P.; Egan, T.J. Insights into the mechanism of action of ferroquine. Relationship between physicochemical properties and antiplasmodial activity. Mol Pharm. 2005, 2, 185–193. [Google Scholar] [CrossRef]
- De Villiers, K.A.; Marques, H.M.; Egan, T.J. The crystal structure of halofantrine-ferriprotoporphyrin IX and the mechanism of action of arylmethanol antimalarials. J. Inorg. Biochem. 2008, 102, 1660–1667. [Google Scholar] [CrossRef]
- Pisciotta, J.M.; Sullivan, D. Hemozoin: oil versus water. Parasitol. Int. 2008, 57, 89–96. [Google Scholar]
- Helms, V. Attraction within the membrane. Forces behind transmembrane protein folding and supramolecular complex assembly. EMBO Rep. 2002, 3, 1133–1138. [Google Scholar] [CrossRef]
- Chavain, N.; Vezin, H.; Dive, D.; Touati, N.; Paul, J.F.; Buisine, E.; Biot, C. Investigation of the Redox Behavior of Ferroquine, a New Antimalarial. Mol. Pharm. 2008, 5, 710–716. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dubar, F.; Khalife, J.; Brocard, J.; Dive, D.; Biot, C. Ferroquine, an Ingenious Antimalarial Drug –Thoughts on the Mechanism of Action. Molecules 2008, 13, 2900-2907. https://doi.org/10.3390/molecules13112900
Dubar F, Khalife J, Brocard J, Dive D, Biot C. Ferroquine, an Ingenious Antimalarial Drug –Thoughts on the Mechanism of Action. Molecules. 2008; 13(11):2900-2907. https://doi.org/10.3390/molecules13112900
Chicago/Turabian StyleDubar, Faustine, Jamal Khalife, Jacques Brocard, Daniel Dive, and Christophe Biot. 2008. "Ferroquine, an Ingenious Antimalarial Drug –Thoughts on the Mechanism of Action" Molecules 13, no. 11: 2900-2907. https://doi.org/10.3390/molecules13112900