Application of Metabolic Engineering to the Production of Scopolamine

Abstract
:Introduction
- Increasing the number of producing cells
- Increasing the carbon flux through a biosynthetic pathway by overexpression of genes codifying for rate-limiting enzymes or blocking the mechanism of feedback inhibition and competitive pathways
- Decreasing catabolism

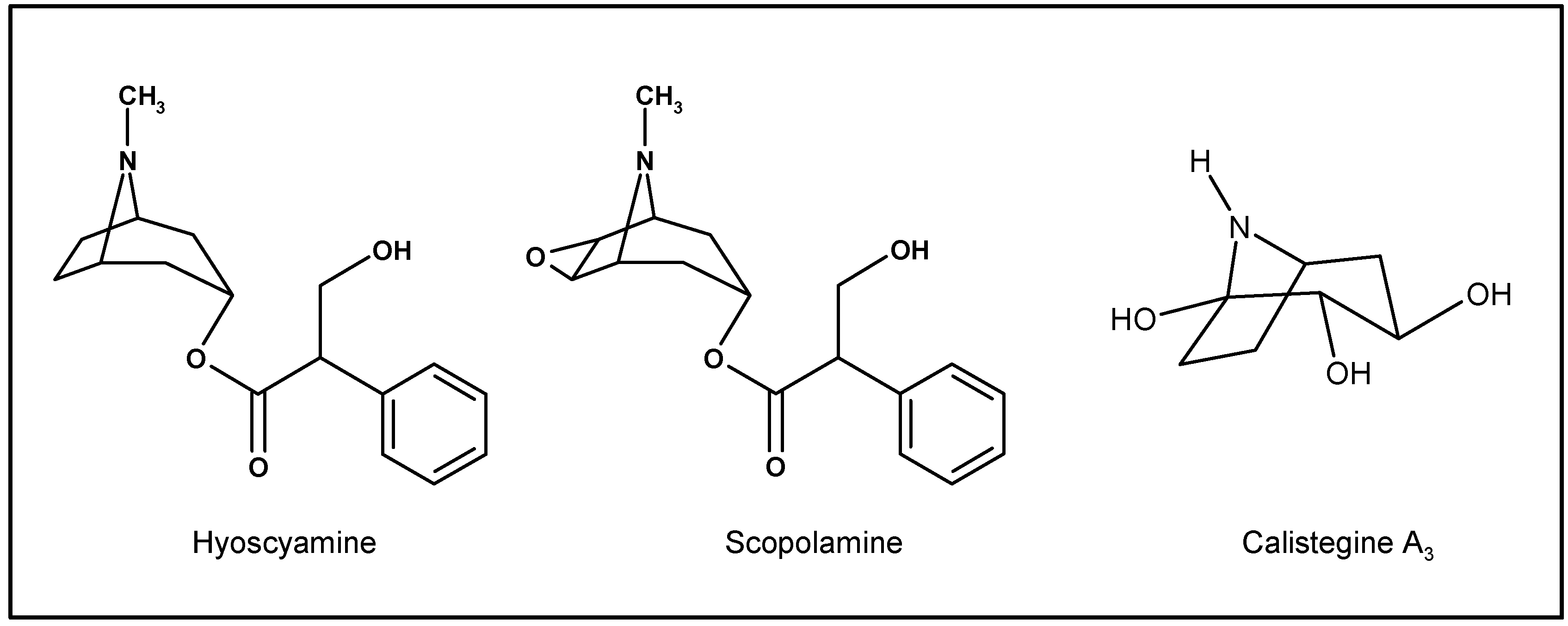
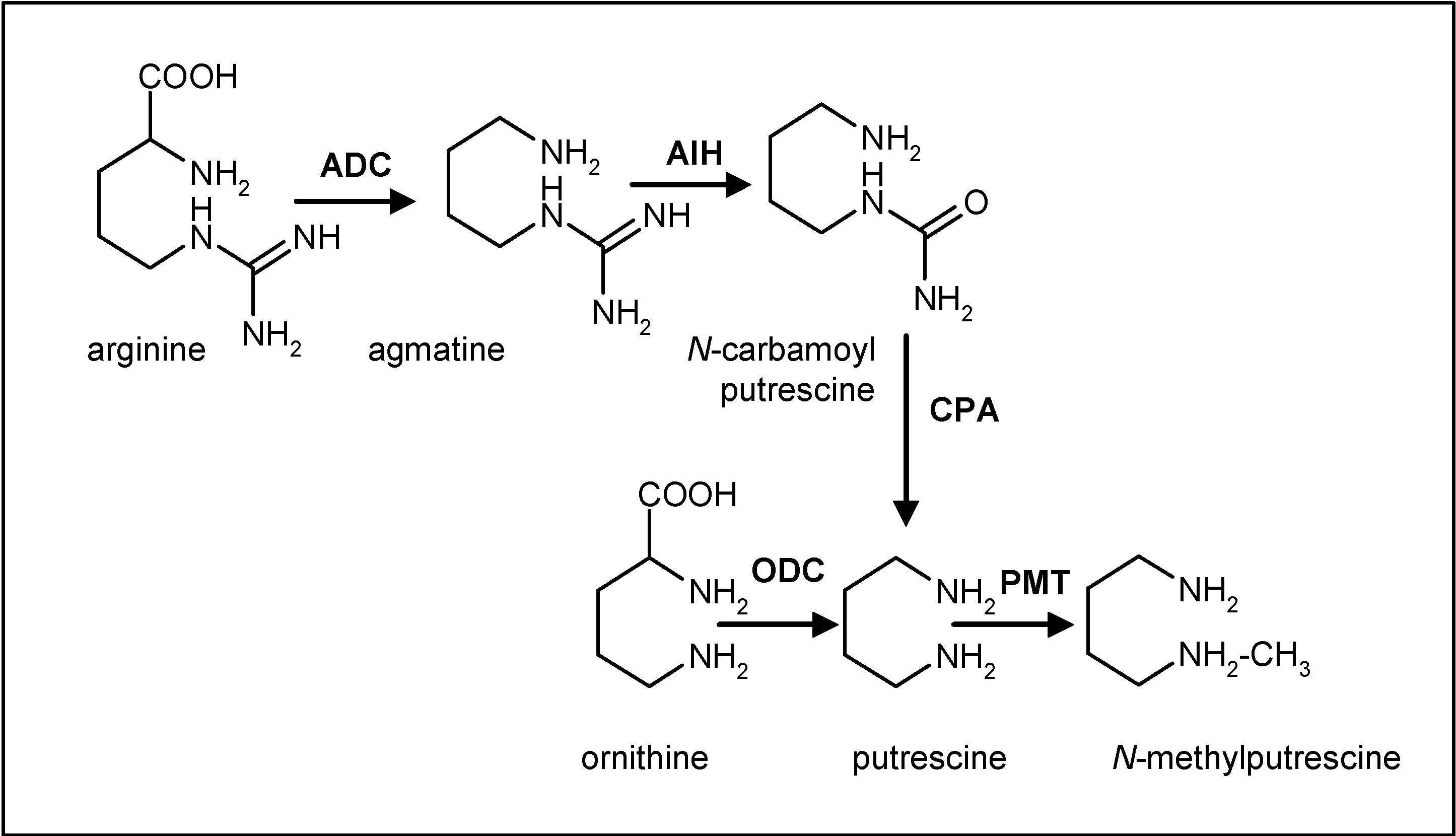
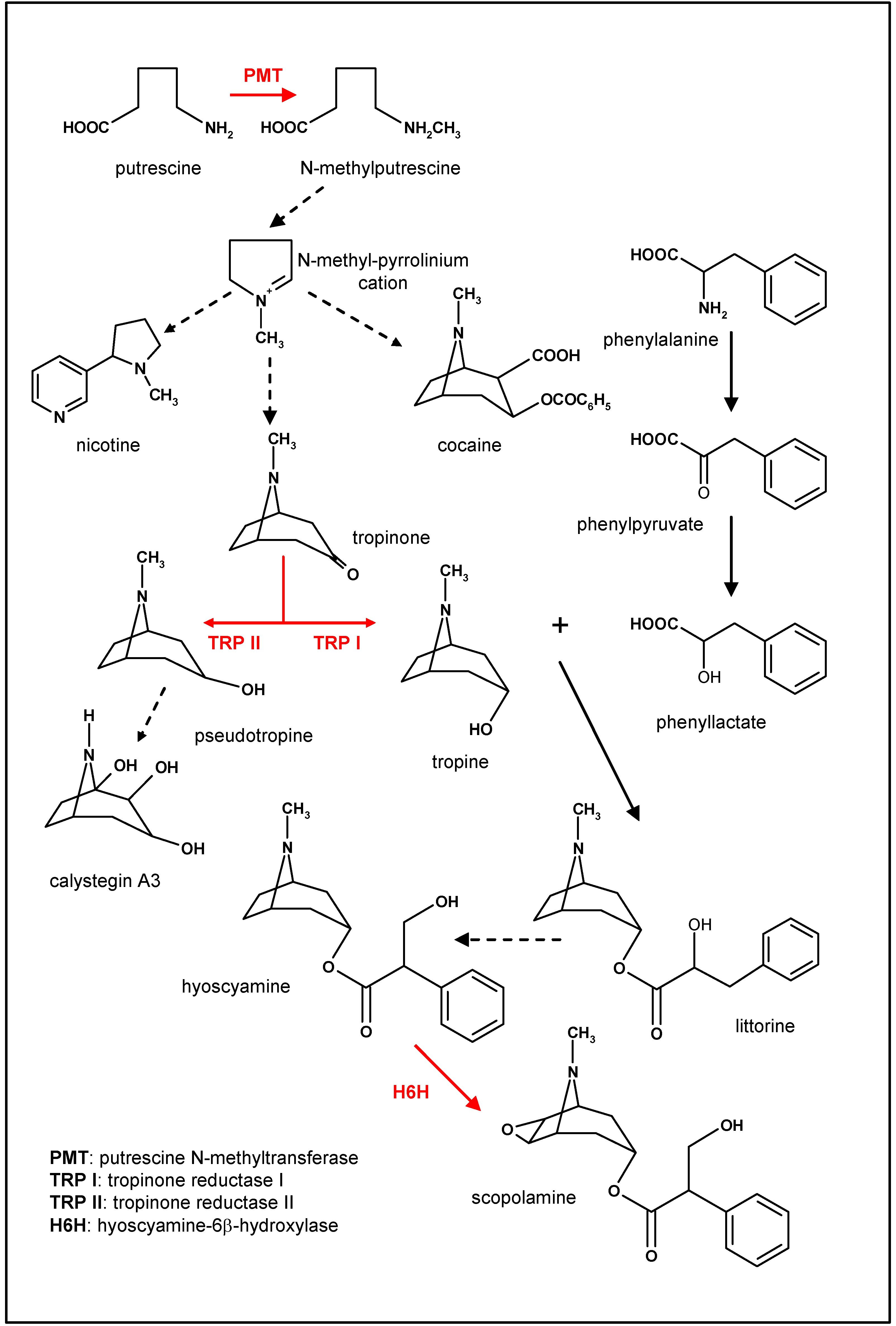
Biosynthesis of scopolamine

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solanoidae | Daturae | Datura | D. stramonium, D. ferox, D. quercifolia, D. pruinosa, D. leichahhardtii, D. inoxia, D. discolor, D. metel, D. wrightii |
| Brugmansia | B. aurea, B. sanguinea, B. arborea, B. xcandida, B. xdolichocarpa, B. xinsignis, B. versicolor, B. vulcanicola | ||
| Solandrae | Solandra | S. longifolia, S. grandifolia, S. guttata, S. hartvegii, S. hirsute, S. macranthe | |
| Solaneae | Atropa | A. belladonna | |
| Latua | L. pubiflora | ||
| Acristus | A. arborea | ||
| Mandragora | M. autumnale, M. vernalis | ||
| Salpichroa | S. organiflora | ||
| Hyoscyameae | Scopolia | S. carniolica, S. parviflora | |
| Hyoscyamus | H. muticus, H. niger, H. albus, H. aureus | ||
| Physochlaina | P. physaloides, P. orientalis | ||
| Przewalskia | P. tangutica | ||
| Cestroideae | Anthocercidae | Duboisia | D. hopwoodii, D. leichhardtii, D. myoporoides, D. arenitensis, D. hybrid |
| Anthotroche | A. myoporoides, A. pannosa, A. walcottii | ||
| Anthocercis | A. littorea, A. viscose, A. fasciculote, A. ilicitolia, A. genistoides | ||
| Cyphanthera | C. anthocercidea, C. albicans | ||
| Symonanthus | S. aromaticus | ||
| Grammosolen | G. dixonii |


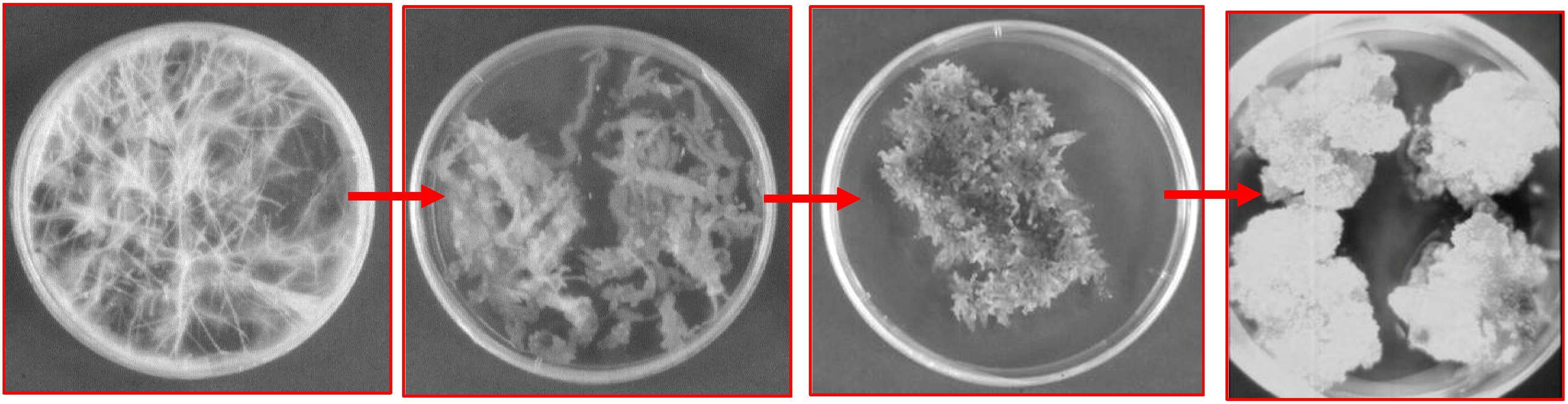
Hairy root cultures as a source of scopolamine

Overexpression of the pmt gene to improve scopolamine production

Overexpression of tropinone reductases
Overexpression of the h6h gene to increase the epoxidation of hyoscyamine into scopolamine
Biotransformation of hyoscyamine into scopolamine in transgenic tobacco hairy roots

Future Challenges
Acknowledgements
References
- Canter, P. H.; Thomas, H.; Ernst, E. Brinding medicinal plants into cultivation: opportunities and challenges for biotechnology. Trends Biotechnol. 2005, 23, 180–185. [Google Scholar] [CrossRef]
- Lessard, P. Metabolic engineering, the concept coalesces. Nat. Biotechnol. 1996, 14, 1654–1655. [Google Scholar] [CrossRef]
- Kinney, A. J. Manipulating flux through plant metabolic pathways. Curr. Opin. Plant Biol. 1998, 1, 173–178. [Google Scholar] [CrossRef]
- Whitmer, S.; Van der Heijden, R.; Verpoorte, R. Plant Biotechnology and Transgenic Plants; Oksman-Caldentey, K. -M., Barz, W.H., Eds.; Marcel & Dekker: New York-Basel, 2002; pp. 373–405. [Google Scholar]
- Ramachandra, S. R.; Ravishankar, G. A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 1001–153. [Google Scholar]
- Griffing, W. J.; Lin, G. D. Chemotaxonomy and geographical distribution of tropane alkaloids. Phytochemistry 2000, 53, 623–637. [Google Scholar] [CrossRef]
- Jouhikainen, K.; Lindgren, L.; Jokelainen, T.; Hiltunen, R.; Teeri, T.H.; Oksman-Caldentey, K. M. Enhancement of scopolamine production in Hyoscyamus muticus L. hairy root cultures by genetic engineering. Planta 1999, 208, 545–551. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yamada, Y. Alkaloid biogenesis: molecular aspects. Annu. Rev. Plant Phys. 1994, 45, 257–285. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K. M.; Strauss, A. Somaclonal variation of scopolamine content in protoplast-derived cell culture clones of Hyoscyamus muticus. Planta Med. 1986, 52, 6–12. [Google Scholar] [CrossRef]
- Palazón, J.; Altabella, T.; Cusidó, R. M.; Ribó, M.; Piñol, M. T. Growth and tropane alkaloid production in Agrobacterium transformed roots and derived callus of Datura. Biol. Plantar. 1995, 37, 161–168. [Google Scholar] [CrossRef]
- Arroo, R.; Woolley, J.; Oksman-Caldentey, K. M. Biotechnology in Agriculture and Forestry; Pua, E. C., Davey, M. R., Eds.; Springer: Berlin-Heidelberg-New York, 2007; Vol. 61, Transgenic Crops VI,. Pua, E.C.; Davey, M.R., Eds.; Springer: Berlin Heidelberg New York, 2007; pp. 189–204. [Google Scholar]
- Walton, N. J.; Robins, R. J.; Peerless, A. C. J. Enzymes of N-methylputrescine biosynthesis in relation to hyoscyamine formation in transformed root cultures of Datura stramonium and Atropa belladonna. Planta 1990, 182, 136–141. [Google Scholar]
- Yamada, Y.; Hashimoto, T.; Endo, T.; Yukimune, Y.; Cono, J.; Hamaguchi, N.; Dräger, B. Secondary Products from Plant Tissue Culture; Charlwood, R. V., Rhodes, M. J. C., Eds.; Clarendon Press: Oxford, 1990; pp. 227–242. [Google Scholar]
- Robins, R. J.; Parr, A. J.; Bent, E. G. Studies on the biosynthesis of tropane alkaloids in Datura stramonium L. transformed root cultures. Planta 1991, 183, 385–390. [Google Scholar]
- Ansarin, M.; Woolly, J. G. The biosynthesis of tropic acid. Part 6. Enantioselective, intact incorporation of (R)-(+)-3-phenyllactic acid into the tropic acid ester alkaloids of Datura. J. Chem. Soc. Perkin Trans. I 1995, 487–490. [Google Scholar] [CrossRef]
- Humphrey, A. J.; O’Hagan, D. Tropane alkaloid biosynthesis. A century old problem unresolved. Nat. Prod. Rep. 2001, 18, 494–502. [Google Scholar] [CrossRef]
- Richter, U.; Rothe, G.; Fabian, A. K.; Rahfeld, B.; Dräger, B. Overexpression of tropinone reductases alters alkaloid composition in Atropa belladonna root cultures. J. Exp. Bot. 2005, 56, 645–652. [Google Scholar] [CrossRef]
- Rothe, G.; Garske, U.; Draeger, B. Calystegines in root cultures of Atropa belladonna respond to sucrose, not to elicitation. Plant Sci. 2001, 160, 1043–1053. [Google Scholar] [CrossRef]
- Leete, E.; Kowano, N.; Newmark, R. A. Use of Carbon-13 Nuclear Magnetic Resonance to establish that the biosynthesis of tropic acid involves an intramolecular rearrangement of phenylalanine. J. Am. Chem. Soc. 1975, 97, 6826–6830. [Google Scholar] [CrossRef]
- Ansarin, M.; Woolly, J. G. The rearrangement of phenyllactate in the biosynthesis of tropic acid. Phytochemistry 1994, 35, 935–939. [Google Scholar] [CrossRef]
- Chesters, N. C. J. E.; O’Hagan, D.; Robins, R. J. The biosynthesis of tropic acid: The (R)-D-Phenyllactic moiety is processed by the mutase involved in hyoscyamine biosynthesis in Datura stramonium. J. Chem. Soc. Chem. Comm. 1995, 2, 127–129. [Google Scholar]
- Robins, R. J.; Chesters, N. C. G. E.; O’Hagan, D.; Parr, A.; Walton, N. J.; Woolley, J. G. The biosynthesis of hyoscyamine: the process by which littorine rearranges to hyoscyamine. J. Chem. Soc. Perkin Trans. I 1995, 481–485. [Google Scholar]
- Patterson, S.; O’Hagan, D. Biosynthetic studies on the tropane alkaloid hyoscyamine in Datura stramonium; hyoscyamine is stable to in vivo oxidation and is not derived from littorine via a vicinal interchange process. Phytochemistry 2002, 61, 323–329. [Google Scholar] [CrossRef]
- Li, R.; Reed, D. W.; Liu, E.; Nowak, J.; Pelcher, L. E.; Page, J. E.; Covello, P. S. Functional genomic analysis of alkaloid biosynthesis in Hyoscyamus niger reveals a cytochrome P450 involved in littorine rearrangement. Chem. Biol. 2006, 13, 513–520. [Google Scholar] [CrossRef]
- Toivonen, L. Utilization of hairy root cultures for production of secondary metabolites. Biotechnol. Prog. 1993, 9, 12–20. [Google Scholar] [CrossRef]
- Palazón, J.; Piñol, M. T.; Cusido, R. M.; Morales, C.; Bonfill, M. Application of transformed root technology to the production of bioactive metabolites. Recent Res. Dev. Plant Phys. 1997, 1, 125–143. [Google Scholar]
- Palazón, J.; Moyano, E.; Bonfill, M.; Cusidó, R. M.; Piñol, M. T. Floriculture, Ornamental and Plant Biotechnology: Advances and Topical Issues; Teixeira da Silva, J. A., Ed.; Global Science Books, Ltd: London, UK, 2006; Vol. II, pp. 209–221. [Google Scholar]
- Suzuki, K.; Yamada, Y.; Hashimoto, T. Expression of Atropa belladonna putrescine N-methyltransferase gene in root pericycle. Plant Cell Physiol. 1999, 40, 289–297. [Google Scholar] [CrossRef]
- Hashimoto, T.; Hayashi, A.; Amano, Y.; Cono, J.; Iwanari, H.; Usuda, S.; Yamada, Y. Hyoscyamine 6β-hydroxilase, an enzyme envolved in tropane alkaloid biosynthesis, is localized at the pericycle of the roots. J. Biol. Chem. 1992, 266, 4648–4653. [Google Scholar]
- Kanegae, T.; Kajiya, H.; Amano, Y.; Hashimoto, T.; Yamada, Y. Species-dependent expression of the hyoscyamine 6β-hydroxilase gene in the pericycle. Plant Physiol. 1994, 105, 483–490. [Google Scholar]
- Nakajima, K.; Hashimoto, T. Two tropinone reductases, that catalyze opposite stereospecific reductions in tropane alkaloid biosynthesis, are localized in plants root with different cell-specific patterns. Plant Cell Physiol. 1999, 40, 1099–1107. [Google Scholar]
- Rothe, G.; Hachiya, A.; Yamada, Y.; Hashimoto, T.; Dräger, B. Alkaloids in plants and roots cultures of Atropa belladona overexpressing putrescine N-methyltransferase. J. Exp. Bot. 2003, 54, 2065–2070. [Google Scholar] [CrossRef]
- Sevón, N.; Oksman-Caldentey, K. M. Agrobacterium rhizogenes-mediates transformation: Root cultures as a source of alkaloids. Planta Med. 2002, 68, 859–868. [Google Scholar] [CrossRef] [Green Version]
- White, F. F.; Nester, E. W. Hairy root: plasmid encodes virulence in Agrobacterium rhizogenes. J. Bacteriol. 1980, 141, 1134–1141. [Google Scholar]
- Jouanin, L. Restriction map of an agropine-type Ri-plasmid and its homologies with Ti-plasmids. Plasmid 1984, 12, 91–102. [Google Scholar] [CrossRef]
- Binns, A. N.; Tomashow, J. V. Cell biology of Agrobacterium infection and transformation of plants. Annu. Rev. Microbiol. 1988, 42, 575–606. [Google Scholar] [CrossRef]
- Morris, R. O. Genes specifying auxin and cytokinin biosynthesis in phytopathogens. Annu. Rev. Plant Physiol. 1986, 37, 509–538. [Google Scholar] [CrossRef]
- Chriqui, D.; Guivarch, A.; Dewitte, W.; Prinsen, E.; Van Onkelen, H. Rol genes and root initiation and development. Plant Soil 1996, 187, 47–55. [Google Scholar] [CrossRef]
- Palazón, J.; Cusidó, R. M.; Roig, C.; Piñol, M. T. Effect of rol genes from Agrobacterium rhizogenes TL-DNA on nicotine production in tobacco root cultures. Plant Physiol. Biochem. 1997, 35, 155–162. [Google Scholar]
- Nilsson, O.; Olsson, O. Getting to the root: the role of the Agrobacterium rhizogenes rol genes in the formation of hairy roots. Physiol. Plant. 1997, 100, 463–473. [Google Scholar] [CrossRef]
- Piñol, M. T.; Palazón, J.; Cusidó, R.; Serrano, M. Effects of Ri T-DNA from Agrobacterium rhizogenes on growth and hyoscyamine production in Datura stramonium root cultures. Bot. Acta 1996, 109, 133–138. [Google Scholar] [CrossRef]
- Bonhomme, V.; Laurain-Mattar, D.; Fliniaux, M. A. Effects of the rol C gene on hairy root: Induction development and tropane alkaloid production by Atropa belladonna. J. Nat. Prod. 2000, 63, 1249–1252. [Google Scholar] [CrossRef]
- Bonhomme, V.; Laurain-Mattar, D.; Lacoux, J.; Fliniaux, M. A.; Jacquin-Dubreil, A. Tropane alkaloid production by hairy roots of Atropa belladonna obtained after transformation with Agrobacterium rhizogenes 15834 and Agrobacterium tumefaciens containing rol A, B, C genes only. J. Biotechnol. 2000, 81, 151–158. [Google Scholar] [CrossRef]
- Palazón, J.; Cusidó, R. M.; Gonzalo, J.; Bonill, M.; Morales, C.; Piñol, M. T. Relation between the amount of rol C gene product and indole alkaloid accumulation in Catharanthus roseus transformed root cultures. J. Plant Physiol. 1998, 153, 712–718. [Google Scholar]
- Bulgakov, V. P.; Khodakovskaya, M. V.; Labetskaya, N. V.; Chernoded, G. K.; Zhuravlev, Y. N. The impact of plant rol C oncogene on ginsenoside production by ginseng hairy root cultures. Phytochemistry 1998, 49, 1929–1934. [Google Scholar] [CrossRef]
- Moyano, E.; Fornalé, S.; Palazón, J.; Cusidó, R. M.; Bonfill, M.; Morales, C.; Piñol, M. T. Effect of Agrobacterium rhizogenes T-DNA on alkaloid production in Solanaceae plants. Phytochemistry 1999, 52, 1287–1292. [Google Scholar] [CrossRef]
- Mallol, A.; Cusidó, R. M.; Palazón, J.; Bonfill, M.; Morales, C.; Piñol, M. T. Ginsenoside production in different phenotypes of Panax ginseng transformed roots. Phytochemistry 2001, 57, 365–371. [Google Scholar] [CrossRef]
- Furmanova, M.; Syklowska-Baranek, K. Hairy root cultures of Taxus x media var. Hicksii Rehd. as a new source of paclitaxel and 10-deacetylbaccatin III. Biotechnol. Lett. 2000, 22, 606–616. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O. L.; Miller, R. A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar] [CrossRef]
- Schenk, R. V.; Hildebrandt, A. C. Medium techniques for induction and growth of monocotyledonous and dycotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Sevón, N.; Oksman-Caldentey, K. M. Agrobacterium rhizogenes-mediated transformation: Root cultures as a source of alkaloids. Planta Med. 2002, 68, 859–868. [Google Scholar] [CrossRef]
- Cusidó, R.M.; Palazón, J.; Piñol, M. T.; Bonfill, M.; Morales, C. Datura metel: In vitro production of tropane alkaloids. Planta Med. 1999, 65, 144–148. [Google Scholar] [CrossRef]
- Eibl, R.; Eibl, D. Plant Biotechnology and Transgenic Plant; Oksman-Caldentey, K.M., Barz, W. H., Eds.; Marcel & Dekker: New York, 2002; pp. 163–200. [Google Scholar]
- Srivastava, S.; Srivastava, A. K. Hairy root culture for mass-production of high-value secondary metabolites. Crit. Rev. Biotechnol. 2007, 27, 29–43. [Google Scholar] [CrossRef]
- Wilson, P. D. G. Hairy roots: Culture and applications; Doran, P. M., Ed.; Harwood Academic Publishers: Amsterdam, 1997; pp. 179–190. [Google Scholar]
- Carvalho, E. B.; Curtis, W. R. Characterization of fluid-flow resistance in root cultures with a connective flow tubular bioreactor. Biotechnol. Bioeng. 1998, 60, 375–384. [Google Scholar] [CrossRef]
- Williams, G. R.; Doran, P. M. Hairy root culture in a liquid-dispersed bioreactor: characterization of spatial heterogeneity. Biotechnol. Progr. 2000, 16, 391–401. [Google Scholar] [CrossRef]
- Min, J. Y.; Jung, H. Y.; Kang, S. M.; Kim, Y. D.; Kang, Y. M.; Park, D. J.; Prasad, D. T.; Choi, M. S. Production of tropane alkaloids by small-scale bubble column bioreactor cultures of Scopolia parviflora adventitious roots. Bioresour. Technol. 2007, 98, 1748–1753. [Google Scholar] [CrossRef]
- Eibl, R.; Eibl, D. Plant Tissue Culture Engineering. Focus on Biotechnology; vol 6, Gupta, S. D., Ibaraki, Y., Eds.; Springer: Berlin-Heidelberg-New York, 2006; pp. 203–227. [Google Scholar]
- Dräger, B. Chemistry and biology of calystegines. Nat. Prod. Rep. 2004, 21, 211–223. [Google Scholar] [CrossRef]
- Feth, F.; Arfmann, H. A.; Wray, V.; Wagner, K. G. Determination of putrescine N-methyltransferase by high performance liquid chromatography. Phytochemistry 1985, 24, 921–923. [Google Scholar] [CrossRef]
- Walton, N. J.; Peerless, A. C. J.; Robins, R. J.; Rhodes, M. J. C.; Boswell, H. D.; Robins, D. J. Purification and properties of putrescine N-methyltransferase from transformed roots of Datura stramonium L. Planta 1994, 193, 9–15. [Google Scholar]
- Hashimoto, T.; Yukimune, Y.; Yamada, Y. Putrescine and putrescine N-methyltransferase in the biosynthesis of tropane alkaloids in cultured roots of Hyoscyamus albus. Planta 1989, 178, 123–130. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yukimune, Y.; Yamada, Y. Putrescine and putrescine N-methyltransferase in the biosynthesis of tropane alkaloids in cultured roots of Hyoscyamus albus. Planta 1989, 178, 131–137. [Google Scholar] [CrossRef]
- Hibi, N.; Fujita, T.; Hatano, M.; Hashimoto, T.; Yamada, Y. Putrescine N-methyltransferase in cultured roots of Hyoscyamus albus: n-butylamine as a potent inhibitor of the transferase both in vitro and in vivo. Plant Physiol. 1992, 100, 820–825. [Google Scholar] [CrossRef]
- Shoji, T.; Yamada, Y.; Hashimoto, T. Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol. 2000, 41, 831–839. [Google Scholar] [CrossRef]
- Baldwin, I. T.; Zhang, Z.; Diab, N.; Ohnmeiss, T. E.; McCloud, E. S.; Lynds, G. Y.; Schmelz, E. A. Quantification, correlations and manipulations of wound-induced changes in jasmonic acid and nicotine in Nicotiana sylvestris. Planta 1997, 201, 397–404. [Google Scholar] [CrossRef]
- Shoji, T.; Nakajima, K.; Hashimoto, T. Ethylene suppresses jasmonate-induced gene expression in nicotine biosynthesis. Plant Cell Physiol. 2000, 41, 1072–1076. [Google Scholar] [CrossRef]
- Imanishi, S.; Katsuhito, K.; Nakakita, M.; Kojima, H.; Matsubayashi, Y.; Hashimoto, T.; Sakagami, Y.; Yamada, Y.; Nakamura, K. Differential induction by methyl jasmonate of genes encoding ornithine decarboxylase and other enzymes involved in nicotine biosynthesis in tobacco cell cultures. Plant Mol. Biol. 1998, 38, 1101–1111. [Google Scholar] [CrossRef]
- Zhang, L.; Kai, G. Y.; Lu, B. B.; Zhang, H. M.; Tang, K. X.; Jiang, J. H.; Chen, W. S. Metabolic engineering of tropane alkaloid biosynthesis in plants. J. Integr. Plant Biol. 2005, 47, 136–143. [Google Scholar] [CrossRef]
- Sato, F.; Hashimoto, T.; Hachiya, A.; Tamura, K.; Choi, K.; Morishige, T.; Fujimoto, H.; Yamada, Y. Metabolic engineering of plant alkaloid biosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 367–372. [Google Scholar] [CrossRef]
- Moyano, E.; Fornalé, S.; Palazón, J.; Cusidó, R. M.; Bagni, N.; Piñol, M. T. Alkaloid production in Duboisia hybrid hairy root cultures overexpressing the pmt gene. Phytochemistry 2002, 59, 697–702. [Google Scholar] [CrossRef]
- Moyano, E.; Jouhikainen, K; Tammela, P.; Palazón, J; Cusido, R. M.; Piñol, M. T.; Teeri, T. H.; Oksman-Caldentey, K. M. Effect of pmt gene overexpression on tropane alkaloid production in transformed root cultures of Datura metel and Hyoscyamus muticus. J. Exp. Bot. 2003, 54, 203–211. [Google Scholar] [CrossRef]
- Biondi, S.; Scaramagli, S.; Oksman-Caldentey, K. M. Secondary metabolism in roots and callus cultures of Hyoscyamus muticus L.: The relationship between morphological organization and the response to methyl jasmonate. Plant Sci. 2002, 163, 563–569. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, B.; Lu, B.; Kai, G.; Wang, Z.; Xia, Y.; Ding, R.; Zhang, H.; Sun, X.; Chen, W.; Tang, K. Tropane alkaloids production in transgenic Hyoscyamus niger hairy root cultures over-expressing putrescine N-methyltransferase is methyl jasmonate-dependent. Planta 2007, 225, 887–896. [Google Scholar] [CrossRef]
- Nakajima, K; Hashimoto, T.; Yamada, Y. Two tropinone reductases with different stereospecificities are short-chain dehydrogenases evolved from a common ancestor. P. Nat. Acad. Sci. USA 1993, 90, 9591–9595. [Google Scholar] [CrossRef]
- Nakajima, K.; Yamashita, A.; Akama, H.; Nakatsu, T.; Kato, H.; Hashimoto, T.; Oda, J.; Yamada, Y. Crystal structures of two tropinone reductases: different reactions stereospecificities in the same protein fold. Proc. Natl. Acad. Sci. USA 1998, 95, 4876–4881. [Google Scholar] [CrossRef]
- Portsteffen, A.; Draeger, B.; Nahrstedt, A. The reduction of tropinone in Datura stramonium root cultures by two specific reductases. Phytochemistry 1994, 37, 391–400. [Google Scholar] [CrossRef]
- Draeger, B.; Hashimoto, T.; Yamada, Y. Purification and characterization of pseudotropine forming tropinone reductase from Hyoscyamus niger root cultures. Agric. Biol. Chem. 1988, 52, 2663–2667. [Google Scholar] [CrossRef]
- Draeger, B.; Schaal, A. Tropinone reduction in Atropa belladonna root cultures. Phytochemistry 1994, 35, 1441–1447. [Google Scholar] [CrossRef]
- Nakajima, K.; Hashimoto, T.; Yamada, Y. cDNA encoding tropinone reductase-II from Hyoscyamus niger. Plant Physiol. 1993, 103, 1465–1466. [Google Scholar]
- Nakajima, K.; Hashimoto, T. Two tropinone reductases that catalyze opposite stereospecific reductions in tropane alkaloid biosynthesis are localized in plant root with different cell-specific patterns. Plant Cell Physiol. 1999, 40, 1099–1107. [Google Scholar] [CrossRef]
- Dräger, B. Tropinone reductases, enzymes at the branch point of tropane alkaloid metabolism. Phytochemistry 2006, 67, 327–337. [Google Scholar] [CrossRef]
- Yun, D. J.; Hashimoto, T.; Yamada, Y. Metabolic engineering of medicinal plants: Transgenic Atropa belladonna with an improved alkaloid composition. Proc. Natl. Acad. Sci. USA 1992, 89, 11799–11803. [Google Scholar] [CrossRef]
- Hashimoto, T.; Yun, D. J.; Yamada, Y. Production of tropane alkaloids in genetically engineered root cultures. Phytochemistry 1993, 32, 713–718. [Google Scholar] [CrossRef]
- Palazón, J.; Moyano, E.; Cusido, R.M.; Bonfill, M.; Oksman-Caldentey, K. M.; Piñol, M. T. Alkaloid production in Duboisia hybrid hairy roots and plants overexpresing the h6h gene. Plant Sci. 2003, 165, 1289–1295. [Google Scholar] [CrossRef]
- Zarate, R.; Jaber-Vazdekis, N.; Medina, B.; Ravelo, A. G. Tailoring tropane alkaloid accumulation in transgenic hairy roots of Atropa baetica by over-expressing the gene encoding hyoscyamine 6β-hydroxylase. Biotechnol. Lett. 2006, 28, 1271–1277. [Google Scholar] [CrossRef]
- Zhang, L.; Ding, R.; Chai, Y.; Bonfill, M.; Moyano, E.; Oksman-Caldentey, K. M.; Xu, T.; Pi, Y.; Wang, Z.; Zhang, H.; Kai, G.; Liao, Z.; Sun, X.; Tang, K. Engineering tropane biosynthetic pathway in Hyoscyamus niger hairy root cultures. Proc. Natl. Acad. Sci. USA 2004, 101, 6786–6791. [Google Scholar] [CrossRef]
- Rocha, P.; Stenzel, O.; Parr, A.; Walton, N.; Christou, P.; Dräger, B.; Leech, M. J. Functional expression of tropinone reductase I (trI) and hyoscyamine-6b-hydroxylase (h6h) from Hyoscyamus niger in Nicotiana tabacum. Plant Sci. 2002, 162, 905–913. [Google Scholar] [CrossRef]
- Häkkinen, S. T.; Moyano, E.; Cusidó, R. M.; Palazón, J.; Piñol, M. T.; Oksman-Caldentey, K. M. Enhanced secretion of tropane alkaloids in Nicotiana tabacum hairy roots expressing heterologous hyoscyamine-6ß-hydroxylase. J. Exp. Bot. 2005, 56, 2611–2618. [Google Scholar] [CrossRef]
- Moyano, E; Palazón, J.; Bonfill, M.; Osuna, L.; Cusido, R. M.; Oksman-Caldentey, K. M.; Piñol, M. T. Biotransformation of hyoscyamine into scopolamine in transgenic tobacco cell cultures. J. Plant Physiol. 2007, 164, 521–524. [Google Scholar] [CrossRef]
- Oksman-Caldentey, K. M.; Inzé, D. Plant cell factories in the post genomic era: new ways to produce designer secondary metabolites. Trends Plant Sci. 2004, 9, 433–440. [Google Scholar] [CrossRef]
- Goossens, A.; Häkkinen, T.; Laakso, I.; Seppänen-Laakso, T.; Biondi, S.; De Sutter, V.; Lammertyn, F.; Nuutila, M.; Söderlund, H.; Zabeau, M.; Inzé, D.; Oksman-Caldentey, K. M. A functional genomics approach toward the understanding of secondary metabolism in plant cells. Proc. Natl. Acad. Sci. USA 2003, 100, 8595–8600. [Google Scholar] [CrossRef]
- Rischer, H.; Orešič, M.; Seppänen-Laakso, T.; Katajamaa, M.; Lammertyn, F.; Ardiles-Diaz, W.; Van Montagu, M. C. E.; Inzé, D.; Oksman-Caldentey, K. M.; Goossens, A. Gene-to-metabolite networks for terpenoid indole alkaloid biosynthesis in Catharanthus roseus cells. Proc. Natl. Acad. Sci. USA 2006, 103, 5614–5619. [Google Scholar] [CrossRef] [Green Version]
© 2008 by the authors. Licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Palazón, J.; Navarro-Ocaña, A.; Hernandez-Vazquez, L.; Mirjalili, M.H. Application of Metabolic Engineering to the Production of Scopolamine. Molecules 2008, 13, 1722-1742. https://doi.org/10.3390/molecules13081722
Palazón J, Navarro-Ocaña A, Hernandez-Vazquez L, Mirjalili MH. Application of Metabolic Engineering to the Production of Scopolamine. Molecules. 2008; 13(8):1722-1742. https://doi.org/10.3390/molecules13081722
Chicago/Turabian StylePalazón, Javier, Arturo Navarro-Ocaña, Liliana Hernandez-Vazquez, and Mohammad Hossein Mirjalili. 2008. "Application of Metabolic Engineering to the Production of Scopolamine" Molecules 13, no. 8: 1722-1742. https://doi.org/10.3390/molecules13081722