MALDI-TOF MS Profiling of Annonaceous Acetogenins in Annona muricata Products for Human Consumption

Abstract
:Abbreviations
| α-CHCA | α-cyano-4-hydroxycinnamate |
| ACG | Annonaceous acetogenin |
| amu | atomic mass unit |
| APCI | atmospheric pressure chemical ionization |
| DAD | diode array detector |
| DHB | 2,5-hydroxybenzoyc acid |
| ESI | electrospray ionization |
| I.C. | internal calibrant |
| MALDI-TOF | Matrix-Assisted Laser Desorption/Ionization-Time-of-Flight |
| MS | mass spectrometry |
| NMR | nuclear magnetic resonance |
| RP-HPLC | reversed-phase high performance liquid chromatography |
| THAP | 2,4,6-trihydroxy-acetophenone |
| THF | tetrahydrofuran |
| THP | tetrahydropyran |
Introduction
Results and Discussion
Choice of MALDI-TOF parameters and mass calibration

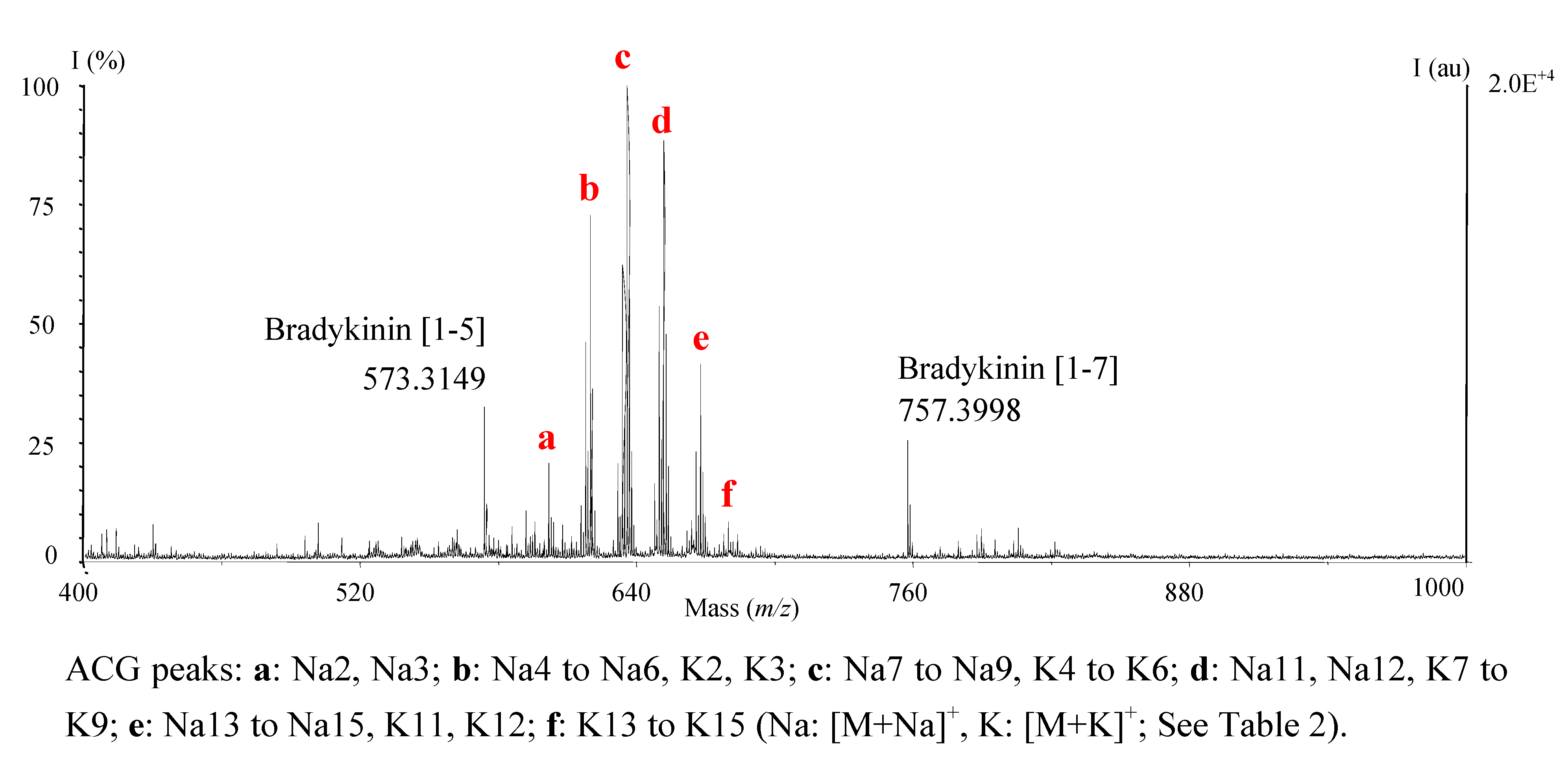{kind=link}
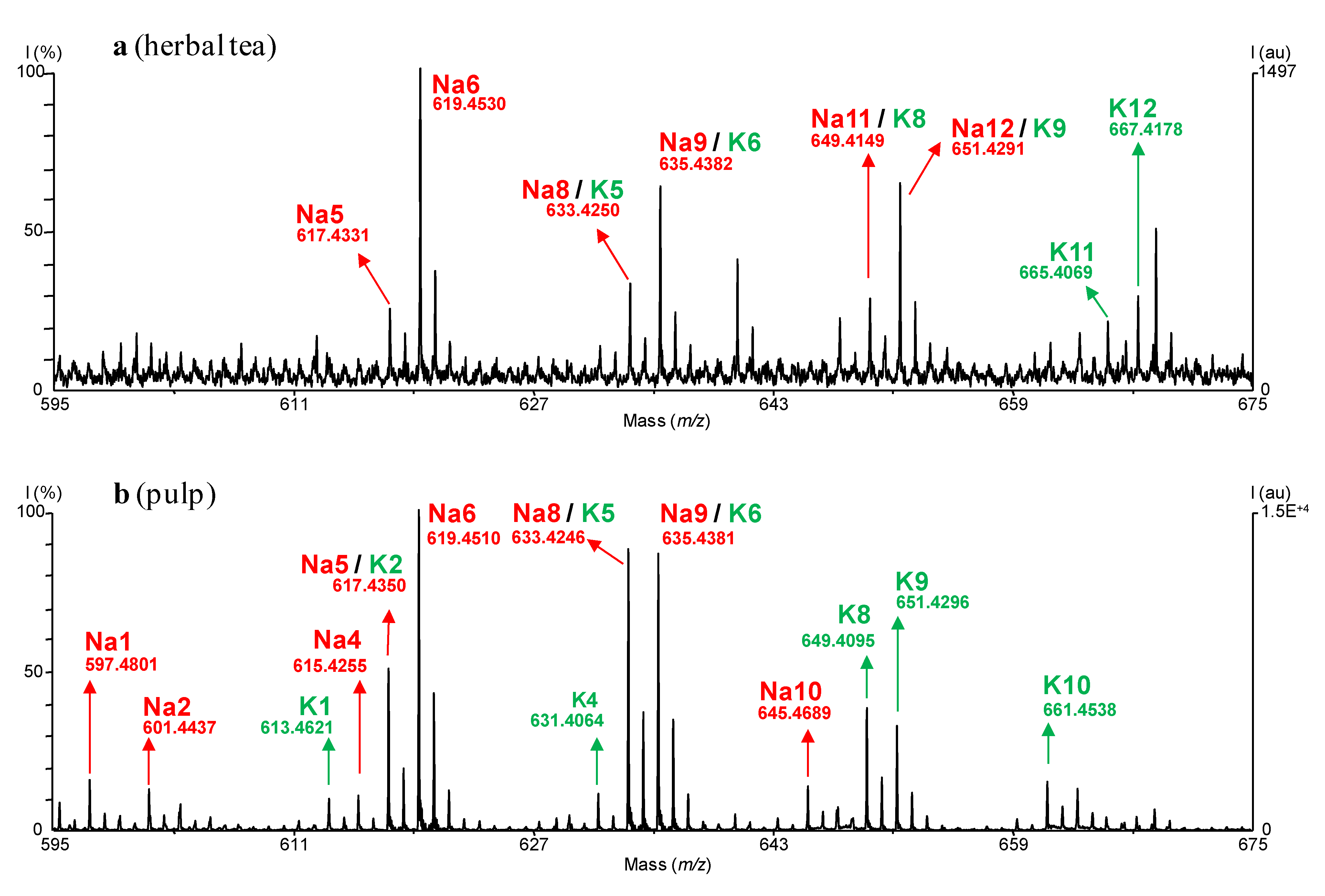{kind=link}
{kind=link}
| Calibration | [M+Na]+ | [M+K]+ | ||||
|---|---|---|---|---|---|---|
| m/z calcd | m/z meas. | error (ppm) | m/z calcd | m/z meas. | error (ppm) | |
| a: Pepmix 5 | 619.4550 | 619.4531 | 3.0 | 635.4289 | 635.4382 | 14.6 |
| a: PEG 400 | 619.4547 | 0.4 | 635.4417 | 20.1 | ||
| b: Pepmix 5 | 619.4532 | 3.1 | 635.4337 | 7.5 | ||

| [M+Na]+ | [M+K]+ | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Formula | Mass | Source | m/z meas. | m/z calcd | error | m/z meas. | m/z calcd | error | ||
| C37H66O4 | 574.4961 | p | Na1 | 597.4801 | 597.4859 | 9.6 | K1 | 613.4621 | 613.4598 | 3.7 |
| n | 597.4792 | 11.2 | 613.4510 | 14.4 | ||||||
| C35H62O6 | 578.4546 | * | Na2 | 601.4406 | 601.4444 | 6.4 | K2, Na5 | 617.4331 | 617.4183 | 23.9 |
| p | 601.4437 | 1.2 | 617.4350 | 27.0 | ||||||
| C35H64O6 | 580.4703 | * | Na3 | 603.4512 | 603.4601 | 14.6 | K3, Na6 | 619.4530 | 619.4340 | 30.6 |
| C35H60O7 | 592.4340 | * | Na4 | 615.4194 | 615.4237 | 6.9 | K4, Na7 | 631.4152 | 631.3976 | 27.9 |
| p | 615.4255 | 3.0 | 631.4064 | 13.9 | ||||||
| n | 615.4225 | 1.8 | 631.4030 | 8.5 | ||||||
| C35H62O7 | 594.4496 | ht | Na5, K2 | 617.4331 | 617.4393 | 10.1 | K5,Na8 | 633.4250 | 633.4133 | 18.5 |
| p | 617.4350 | 6.9 | 633.4246 | 17.8 | ||||||
| n | 617.4407 | 2.2 | 633.4278 | 22.9 | ||||||
| C35H64O7 | 596.4652 | ht | Na6, K3 | 619.4530 | 619.4550 | 3.2 | K6, Na9 | 635.4382 | 635.4289 | 14.6 |
| p | 619.4510 | 6.4 | 635.4381 | 14.4 | ||||||
| n | 619.4552 | 0.1 | 635.4406 | 18.4 | ||||||
| C35H60O8 | 608.4288 | * | Na7, K4 | 631.4152 | 631.4186 | 5.3 | K7 | 647.4011 | 647.3925 | 13.2 |
| C35H62O8 | 610.4445 | ht | Na8, K5 | 633.4250 | 633.4342 | 14.6 | K8, Na11 | 649.4149 | 649.4082 | 10.4 |
| p | 633.4246 | 15.2 | 649.4095 | 2.1 | ||||||
| n | 633.4278 | 10.2 | 649.4142 | 9.3 | ||||||
| C35H64O8 | 612.4601 | ht | Na9, K6 | 635.4382 | 635.4499 | 18.4 | K9, Na12 | 651.4291 | 651.4238 | 8.1 |
| p | 635.4381 | 18.5 | 651.4296 | 9.3 | ||||||
| n | 635.4406 | 14.6 | 651.4344 | 16.2 | ||||||
| C37H66O7 | 622.4809 | p | Na10 | 645.4689 | 645.4706 | 2.6 | K10 | 661.4538 | 661.4446 | 13.9 |
| C35H62O9 | 626.4394 | ht | Na11, K8 | 649.4149 | 649.4292 | 21.9 | K11 | 665.4069 | 665.4031 | 5.8 |
| n | 649.4142 | 23.0 | 665.4083 | 7.8 | ||||||
| C35H64O9 | 628.4550 | ht | Na12, K9 | 651.4291 | 651.4448 | 24.1 | K12 | 667.4178 | 667.4187 | 1.5 |
| p | 651.4296 | 16.0 | 667.4142 | 2.4 | ||||||
| C37H64O8 | 636.4601 | * | Na13 | 659.4312 | 659.4499 | 28.3 | K13 | 675.4328 | 675.4238 | 13.2 |
| C37H66O8 | 638.4758 | * | Na14 | 661.4609 | 661.4655 | 7.0 | K14 | 677.4458 | 677.4395 | 9.3 |
| C37H68O8 | 640.4914 | * | Na15 | 663.4749 | 663.4812 | 9.5 | K15 | 679.4530 | 679.4551 | 3.2 |
Treatment of data
Study of complex crude extracts
- - Refluxed H2O extract of ground leaves (Figure 1);
- - Cup of herbal tea prepared from leaves, according to traditional recipe; In this particular case, the amount of CH2Cl2 extract obtained was low (~1 mg) and ACGs were undetectable with TLC or HPLC-UV, rendering our method particularly useful;
- - Lyophilized pulp from Senegal;
- - Bottled nectar from Venezuela.

Interest of the method, chemotaxonomic significance

Experimental
General
Plant material and preparation of samples
MALDI-TOF experiments and data analysis
Conclusions
Acknowledgements
- Samples Availability: Not available.
References and Notes
- Caparros-Lefèbvre, D.; Elbaz, A. Possible relation of atypical Parkinsonism in the French West Indies with consumption of tropical plants: A case-control study. Lancet 1999, 354, 281–286. [Google Scholar] [CrossRef]
- Caparros-Lefèbvre, D.; Sergeant, N.; Lees, A.; Camuzat, A.; Daniel, S.; Lannuzel, A.; Brice, A.; Tolosa, E.; Delacourte, A.; Duyckaerts, C. Guadeloupean Parkinsonism: A cluster of progressive supranuclear palsy-like tauopathy. Brain 2002, 125, 801–811. [Google Scholar] [CrossRef]
- Lannuzel, A.; Höglinger, G.U.; Verhaeghe, S.; Gire, L.; Belson, S.; Escobar-Khondiker, M.; Poullain, P.; Oertel, W.H.; Hirsch, E.C.; Dubois, B.; Ruberg, M. Atypical Parkinsonism in Guadeloupe: A common risk factor for two closely related phenotypes? Brain 2007, 130, 816–827. [Google Scholar]
- De Cock, V.C.; Lannuzel, A.; Verhaeghe, S.; Roze, E.; Ruberg, M.; Derenne, J.P.; Willer, J.C.; Vidailhet, M.; Arnulf, I. REM sleep behavior disorder in patients with Guadeloupean Parkinsonism, a Tauopathy. Sleep 2007, 30, 1026–1032. [Google Scholar]
- Apartis, E.; Gaymard, B.; Verhaeghe, S.; Roze, E.; Vidailhet, M.; Lannuzel, A. Predominant cortical dysfunction in Guadeloupean parkinsonism. Brain 2008, 131, 2701–2709. [Google Scholar] [CrossRef]
- Angibaud, G.; Gaultier, C.; Rascol, O. Atypical parkinsonism and Annonaceae consumption in New Caledonia. Mov. Disord. 2004, 19, 603–604. [Google Scholar] [CrossRef]
- Camuzat, A.; Romana, M.; Dürr, A.; Feingold, J.; Brice, A.; Ruberg, M.; Lannuzel, A. The PSP-associated MAPT H1 subhaplotype in Guadeloupean atypical parkinsonism. Mov Disord. 2008, 23, 2384–2391. [Google Scholar] [CrossRef]
- Chaudhuri, K.R.; Hu, M.T.M.; Brooks, D.J. Atypical parkinsonism in Afro-Caribbean and Indian origin immigrants to the U.K. Mov. Disord. 2000, 15, 18–23. [Google Scholar] [CrossRef]
- Steele, J.C.; Caparros-Lefèbvre, D.; Lees, A.; Sacks, O. Progressive supranuclear palsy and its relation to pacific foci of the parkinsonism-dementia complex and Guadeloupean Parkinsonism. Parkinsonism Relat. Disord. 2002, 9, 39–54. [Google Scholar] [CrossRef]
- Schapira, A.H.V. Mitochondrial dysfunction in Parkinson’s disease. Cell Death Differ. 2007, 14, 1261–1266. [Google Scholar] [CrossRef]
- Cavé, A.; Figadère, B.; Laurens, A.; Cortes, D. Acetogenins from Annonaceae. In Progress in the Chemistry of Organic Natural Products; Herz, W., Kirby, G.W., Moore, R.E., Steglich, W., Tamm, C., Eds.; Springer: Vienna, New York, NY, USA, 1997; pp. 81–288. [Google Scholar]
- Bermejo, A.; Figadère, B.; Zafra-Polo, M.C.; Barrachina, I.; Estornell, E.; Cortes, D. Acetogenins from Annonceae: Recent progress in isolation, synthesis and mechanisms of action. Nat. Prod. Rep. 2005, 22, 269–303. [Google Scholar] [CrossRef]
- Lannuzel, A.; Michel, P.P.; Höglinger, G.U.; Champy, P.; Jousset, A.; Medja, F.; Lombès, A.; Darios, F.; Gleye, C.; Laurens, A.; Hocquemiller, R.; Hirsch, E.C.; Ruberg, M. The mitochondrial complex I inhibitor annonacin is toxic to mesencephalic dopaminergic neurons by impairment of energy metabolism. Neuroscience 2003, 121, 287–296. [Google Scholar] [CrossRef]
- Champy, P.; Höglinger, G.U.; Féger, J.; Gleye, C.; Hocquemiller, R.; Laurens, A.; Guérineau, V.; Laprévote, O.; Medja, F.; Lombès, A.; Michel, P.P.; Lannuzel, A.; Hirsch, E.C.; Ruberg, M. Annonacin, a lipophilic inhibitor of mitochondrial complex I, induces nigral and striatal neurodegeneration in rats: Possible relevance for atypical parkinsonism in Guadeloupe. J. Neurochem. 2004, 88, 63–69. [Google Scholar]
- Escobar-Khondiker, M.; Höllerhage, M.; Michel, P.P.; Muriel, M.P.; Champy, P.; Yagi, T.; Lannuzel, A.; Hirsch, E.C.; Oertel, W.H.; Ruberg, M.; Höglinger, G.U. Annonacin, a natural mitochondrial complex I inhibitor, causes Tau pathology in cultured neurons. J. Neurosci. 2006, 27, 7828–7837. [Google Scholar]
- Höllerhage, M.; Matusch, A.; Champy, P.; Lombès, A.; Ruberg, M.; Oertel, W.H.; Höglinger, G.U. Natural lipophilic inhibitors of mitochondrial complex I are candidate toxins for sporadic tau pathologies. Exp. Neurol. 2009, 14, 4387–4395. [Google Scholar]
- Champy, P.; Melot, A.; Guérineau, V.; Gleye, C.; Höglinger, G.U.; Ruberg, M.; Lannuzel, A.; Laprévote, O.; Laurens, A.; Hocquemiller, R. Quantification of acetogenins in Annona muricata linked to atypical parkinsonism in Guadeloupe. Mov. Disord. 2005, 20, 1629–1633. [Google Scholar] [CrossRef]
- Champy, P.; Escobar-Khondiker, M.; Bajin ba Ndop, I.; Yamada, E.; Lannuzel, A.; Laprévote, O.; Ruberg, M.; Höglinger, G.U. Atypical parkinsonism induced by Annonaceae: Where are we yet? Planta Med. 2008, 74, 936–937. [Google Scholar]
- Pomper, K.W.; Lowe, J.D.; Crabtree, S.B.; Keller, W.J. Identification of Annonaceous Acetogenins in the ripe fruit of the North American pawpaw (Asimina triloba). J. Agric. Food Chem. 2009, 57, 8339. [Google Scholar] [CrossRef]
- Chen, Y.Y.; Chang, F.R.; Yen, H.F.; Wu, Y.C. Epomusenins A and B, two acetogenins from the fruits of Rollinia mucosa. Phytochemistry 1996, 42, 1081–1083. [Google Scholar] [CrossRef]
- Liaw, C.C.; Chang, F.R.; Wu, M.J.; Wu, Y.C. A novel constituent from Rollinia mucosa, rollicosin, and a new approach to develop Annonaceous acetogenins as potential antitumor agents. J. Nat. Prod. 2003, 66, 279–281. [Google Scholar]
- McLaughlin, J.L. Paw Paw and Cancer: Annonaceous acetogenins from discovery to commercial products. J. Nat. Prod. 2008, 71, 1311–1321. [Google Scholar] [CrossRef]
- Gu, Z.M.; Zhou, D.; Lewis, N.; Wu, J.; Shi, G.; McLaughlin, J.L. Isolation of new bioactive Annonaceous acetogenins from Rollinia mucosa guided by liquid chromatography/mass spectrometry. Bioorg. Med. Chem. 5, 1911–1916.
- Gu, Z.M.; Zhou, D.; Wu, J.; Shi, G.; Zeng, L.; McLaughlin, J.L. Screening for Annonaceous acetogenins in bioactive plants extracts by liquid chromatography/mass spectrometry. J. Nat. Prod. 60, 242–248.
- Martin, J.M.; Madigosky, S.R.; Gu, Z.M.; Zhou, D.; Wu, J.; McLaughlin, J.L. Chemical defense in the zebra swallowtail butterfly, Eurytides marcellus, involving Annonaceous acetogenins. J. Nat. Prod. 1999, 62, 2–4. [Google Scholar] [CrossRef]
- Koomen, J.M.; Russell, W.K.; Tichy, S.E.; Russell, D.H. Accurate mass measurement of DNA oligonucleotide ions using high-resolution time-of-flight mass spectrometry. J. Mass Spectrom. 2002, 37, 357–371. [Google Scholar] [CrossRef]
- Laprévote, O.; Girard, C.; Das, B.C.; Cortes, D.; Cavé, A. Formation of gas-phase lithium complexes from acetogenins and their analysis by fast atom bombardment mass spectrometry. Tetrahedron Lett. 1992, 33, 5237–5240. [Google Scholar] [CrossRef]
- Laprévote, O.; Girard, C.; Das, B.C.; Cortes, D.; Cavé, A. Desorption of lithium complexes of acetogenins by fast atom bombardment: Application to semi-quantitative analysis of crude plant extracts. Analusis 1993, 21, 207–210. [Google Scholar]
- Melot, A.; Fall, D.; Gleye, C; Champy, P. Apolar Annonaceous acetogenins from the fruit pulp of Annona muricata. Molecules 2009, 14, 4387–4395. [Google Scholar] [CrossRef]
- Duret, P.; Laurens, A.; Hocquemiller, R.; Cortes, D.; Cavé, A. Isoacetogenins, artifacts issued from translactonization from Annonaceous acetogenins. Heterocycles 1994, 39, 741–749. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Champy, P.; Guérineau, V.; Laprévote, O. MALDI-TOF MS Profiling of Annonaceous Acetogenins in Annona muricata Products for Human Consumption. Molecules 2009, 14, 5235-5246. https://doi.org/10.3390/molecules14125235
Champy P, Guérineau V, Laprévote O. MALDI-TOF MS Profiling of Annonaceous Acetogenins in Annona muricata Products for Human Consumption. Molecules. 2009; 14(12):5235-5246. https://doi.org/10.3390/molecules14125235
Chicago/Turabian StyleChampy, Pierre, Vincent Guérineau, and Olivier Laprévote. 2009. "MALDI-TOF MS Profiling of Annonaceous Acetogenins in Annona muricata Products for Human Consumption" Molecules 14, no. 12: 5235-5246. https://doi.org/10.3390/molecules14125235