Antiproliferative Effect and Ultrastructural Alterations Induced by Psilostachyin on Trypanosoma cruzi

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
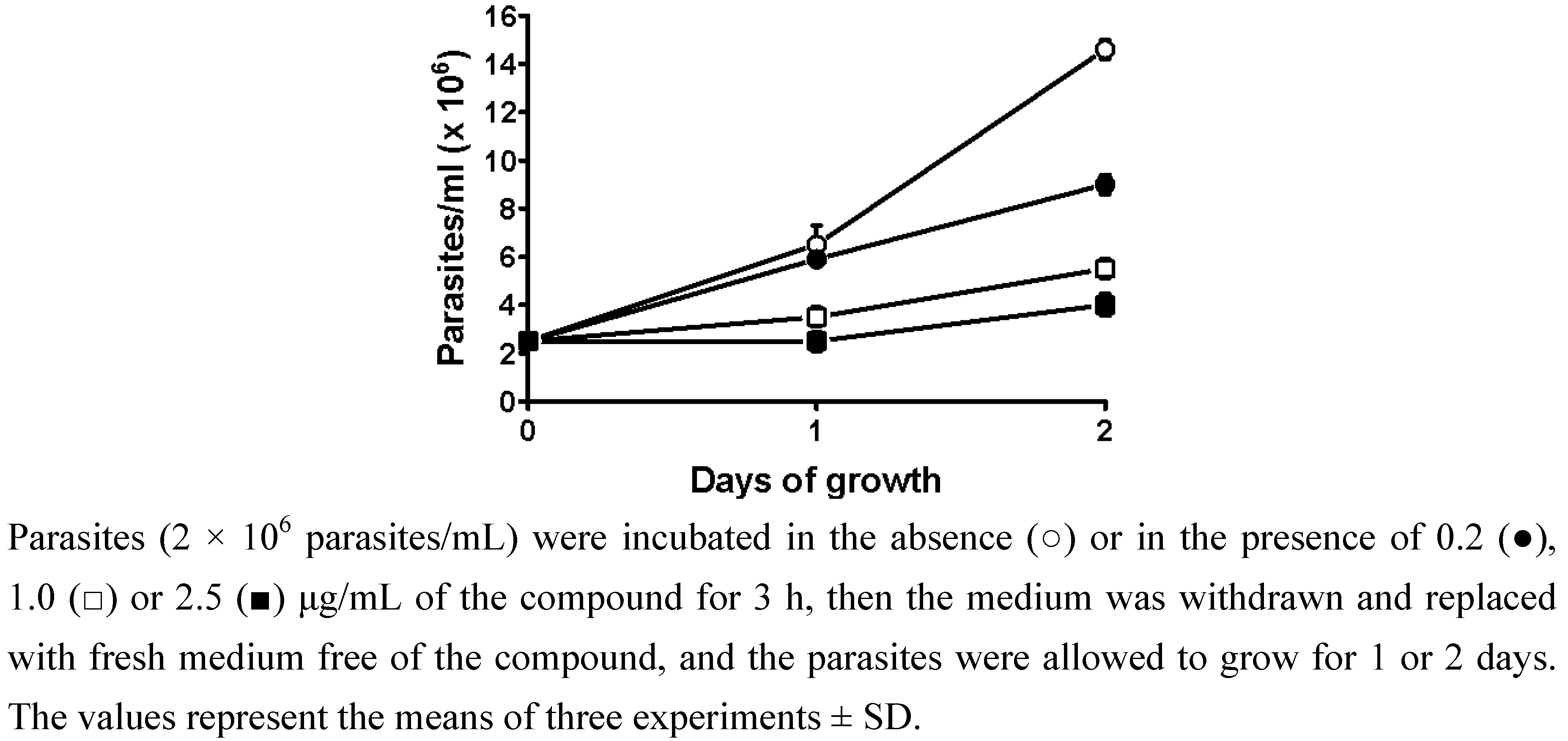
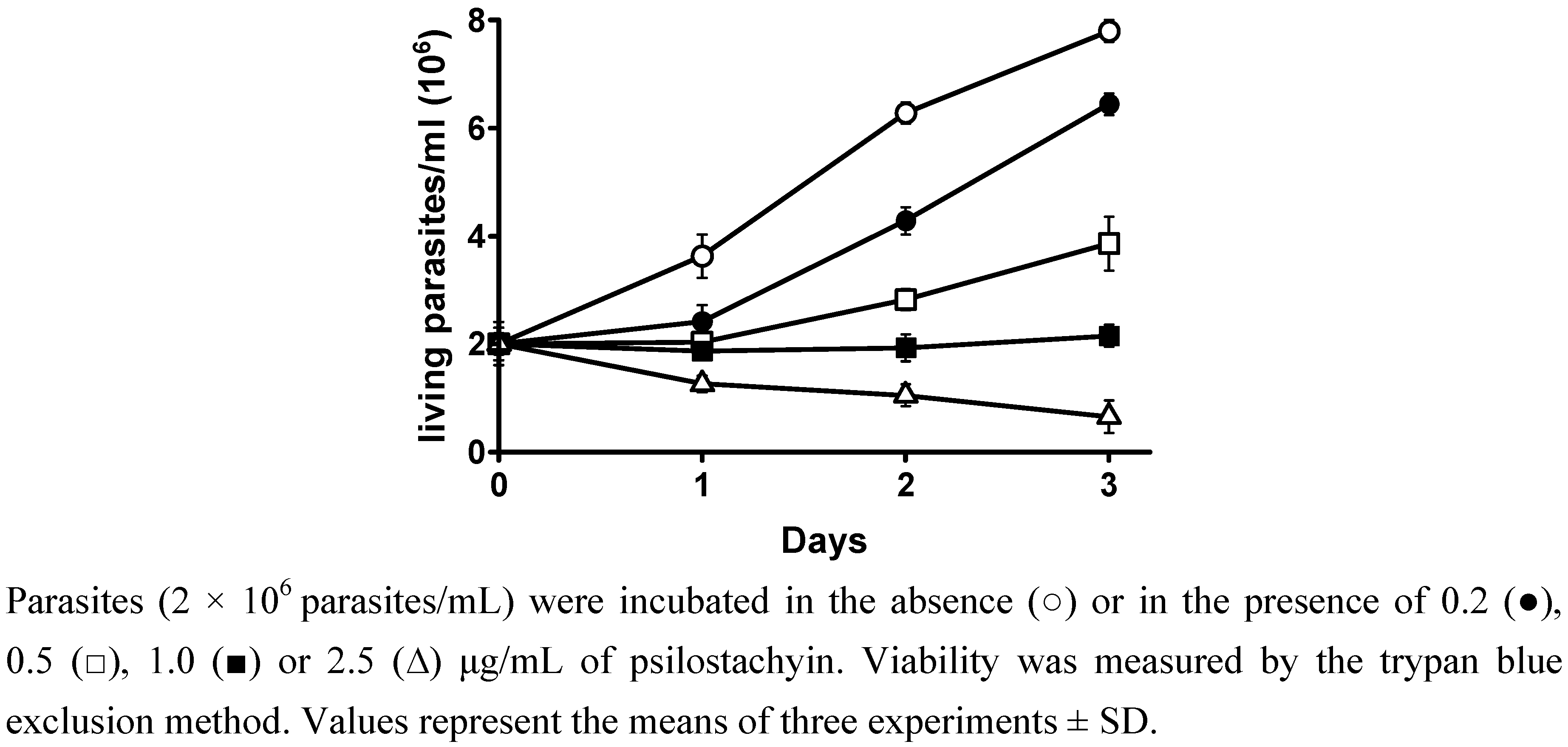
2.1. Viability and reversibility assay


2.2. Cytotoxicity assay on mammalian cells
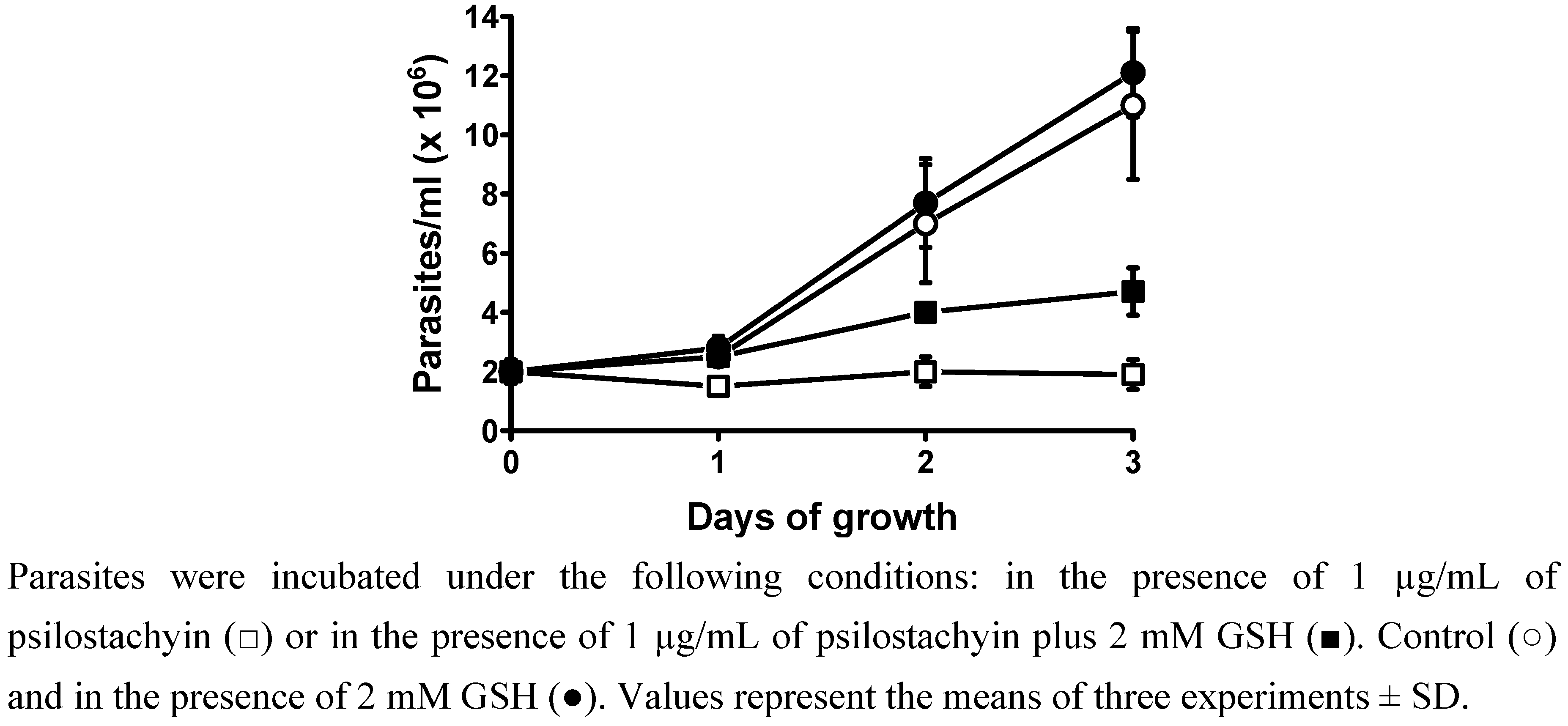
2.3. Effect of psilostachyin on T. cruzi epimastigotes in the presence of glutathione (GSH)

2.4. Transmission electron microscopy

3. Experimental
3.1. Isolation and identification of psilostachyin
3.2. Culture of Trypanosoma cruzi epimastigotes
3.3. Animals
3.4. Viability and reversibility assays
3.5. Cytotoxicity assay
3.6. Effect of psilostachyin on T. cruzi epimastigotes in the presence of GSH
3.7. Transmission electron microscopy
4. Conclusions
Acknowledgements
- Sample Availability: Samples of the compound are available from the corresponding author.
References and Notes
- World Health Organization. Control of Chagas disease. Expert Committee. Second Report; Technical Report Series, N° 905, WHO: Geneva, Switzerland, 2002.
- Amato-Neto, V. Etiological treatment for infection by Trypanosoma cruzi. Mem. Inst. Oswaldo Cruz 1999, 94, 337–339. [Google Scholar] [CrossRef]
- Castro, J.A.; de Meca, M.M.; Bartel, L.C. Toxic side effects of drugs used to treat Chagas´ disease (American trypanosomiasis). Hum. Exp. Toxicol. 2006, 25, 471–479. [Google Scholar] [CrossRef]
- Chan-Bacab, M.J.; Peña-Rodríguez, L.M. Plant natural products with leishmanicidal activity. Nat. Prod. Rep. 2001, 18, 674–688. [Google Scholar] [CrossRef]
- Hoet, S.; Opperdoes, F.; Brun, R.; Quetin-Leclercq, J. Natural products active against African trypanosomes: a step towards new drugs. Nat. Prod. Rep. 2004, 21, 353–364. [Google Scholar] [CrossRef]
- Tasdemir, D.; Kaiser, M.; Brun, R.; Yardley, V.; Schmidt, T.J.; Tosun, F.; Rüedi, P. Antitrypanosomal and antileishmanial activities of flavonoids and their analogues: in vitro, in vivo, structure-activity relationship, and quantitative structure-activity relationship studies. Antimicrob. Agents Chemother. 2006, 50, 1352–1364. [Google Scholar] [CrossRef]
- Salem, M.M.; Werbovetz, K.A. Natural products from plants as drug candidates and lead compounds against leishmaniasis and trypanosomiasis. Curr. Med. Chem. 2006, 13, 2571–2598. [Google Scholar] [CrossRef]
- Picman, A.K. Biological activities of sesquiterpene lactones. Biochem. Syst. Ecol. 1986, 14, 255–281. [Google Scholar] [CrossRef]
- Jisaka, M.; Ohigashi, H.; Takegawa, K.; Huffman, M.A.; Koshimizu, K. Antitumoral and antimicrobial activities of bitter sesquiterpene lactones of Vernonia amygdalina, a possible medicinal plant used by wild chimpanzees. Biosci. Biotechnol. Biochem. 1993, 57, 833–834. [Google Scholar]
- Quintero, A.; Pelcastre, A.; Solano, J.D. Antitumoral activity of new pyrimidine derivatives of sesquiterpene lactones. J. Pharm. Pharm. Sci. 1999, 2, 108–112. [Google Scholar]
- Brengio, S.D.; Belmonte, S.A.; Guerreiro, E.; Giordano, O.S.; Pietrobon, E.O.; Sosa, M.A. The sesquiterpene lactone dehydroleucodine (DhL) affects the growth of cultured epimastigotes of Trypanosoma cruzi. J. Parasitol. 2000, 86, 407–412. [Google Scholar]
- Jimenez-Ortiz, V.; Brengio, S.D.; Giordano, O.; Tonn, C.; Sanchez, M.; Burgos, M.H.; Sosa, M.A. The trypanocidal effect of sesquiterpene lactones helenalin and mexicanin on cultured epimastigotes. J. Parasitol. 2005, 91, 170–174. [Google Scholar] [CrossRef]
- Schmidt, T.; Nour, A.M.M.; Kahlid, S.A.; Kaiser, M.; Brun, R. Quantitative structure-antiprotozoal activity relationships of sesquiterpene lactones. Molecules 2009, 14, 2062–2076. [Google Scholar] [CrossRef]
- Sülsen, V.; Frank, F.; Cazorla, S.; Anesini, C.; Malchiodi, E.; Freixa, B.; Vila, R.; Muschietti, L.; Martino, V. Trypanocidal and leishmanicidal activities of sesquiterpene lactones from Ambrosia tenuifolia Sprengel (Asteraceae). Antimicrob. Agents Chemother. 2008, 52, 2415–2419. [Google Scholar]
- Schmeda-Hirschmann, G.; Bohlmann, F.; Jakupovic, J. The constituents of Ambrosia tenuifolia and Pectis odorata. Rev. Latinoam. Quim. 1986, 17, 200–202. [Google Scholar]
- Izumi, E.; Morello, L.G.; Ueda-Nakamura, T.; Yamada-Ogatta, S.F.; Dias Filho, B.P.; Garcia Cortez, D.A.; Piloto Ferreira, I.C.; Morgado-Diaz, J.A.; Vataro-Nakamura, C. Trypanosoma cruzi: antiprotozoal activity of parthenolide obtained from Tanacetum parthenium (L.) Schultz Bip. (Asteraceae, Compositae) against epimastigote and amastigote forms. Exp. Parasitol. 2007, 118, 324–330. [Google Scholar]
- Lazardi, K.; Urbina, J.A.; De Souza, W. Ultrastructural alterations induced by two ergosterol biosynthesis inhibitors, ketoconazole and terbinafine, on epimastigotes and amastigotes of Trypanosoma (Schizotrypanun) cruzi. Antimicrob. Agents Chemother. 1990, 34, 2097–2105. [Google Scholar] [CrossRef]
- Xu, C.; Ray, D.S. Isolation of proteins associated with kinetoplast DNA networks in vivo. Proc. Natl. Acad. Sci. USA 1993, 90, 1786–1789. [Google Scholar] [CrossRef]
- Cavalcanti, D.P.; Fragoso, S.P.; Goldenberg, S.; De Souza, W.; Motta, M.C. The effect of topoisomerase II inhibitors on the kinetoplast ultrastructure. Parasitol. Res. 2004, 94, 439–448. [Google Scholar] [CrossRef]
- National Research Council. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, D.C., USA, 1996.
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sülsen, V.; Barrera, P.; Muschietti, L.; Martino, V.; Sosa, M. Antiproliferative Effect and Ultrastructural Alterations Induced by Psilostachyin on Trypanosoma cruzi. Molecules 2010, 15, 545-553. https://doi.org/10.3390/molecules15010545
Sülsen V, Barrera P, Muschietti L, Martino V, Sosa M. Antiproliferative Effect and Ultrastructural Alterations Induced by Psilostachyin on Trypanosoma cruzi. Molecules. 2010; 15(1):545-553. https://doi.org/10.3390/molecules15010545
Chicago/Turabian StyleSülsen, Valeria, Patricia Barrera, Liliana Muschietti, Virginia Martino, and Miguel Sosa. 2010. "Antiproliferative Effect and Ultrastructural Alterations Induced by Psilostachyin on Trypanosoma cruzi" Molecules 15, no. 1: 545-553. https://doi.org/10.3390/molecules15010545