LYRM1, a Gene that Promotes Proliferation and Inhibits Apoptosis during Heart Development

Abstract
:1. Introduction
2. Results and Discussion
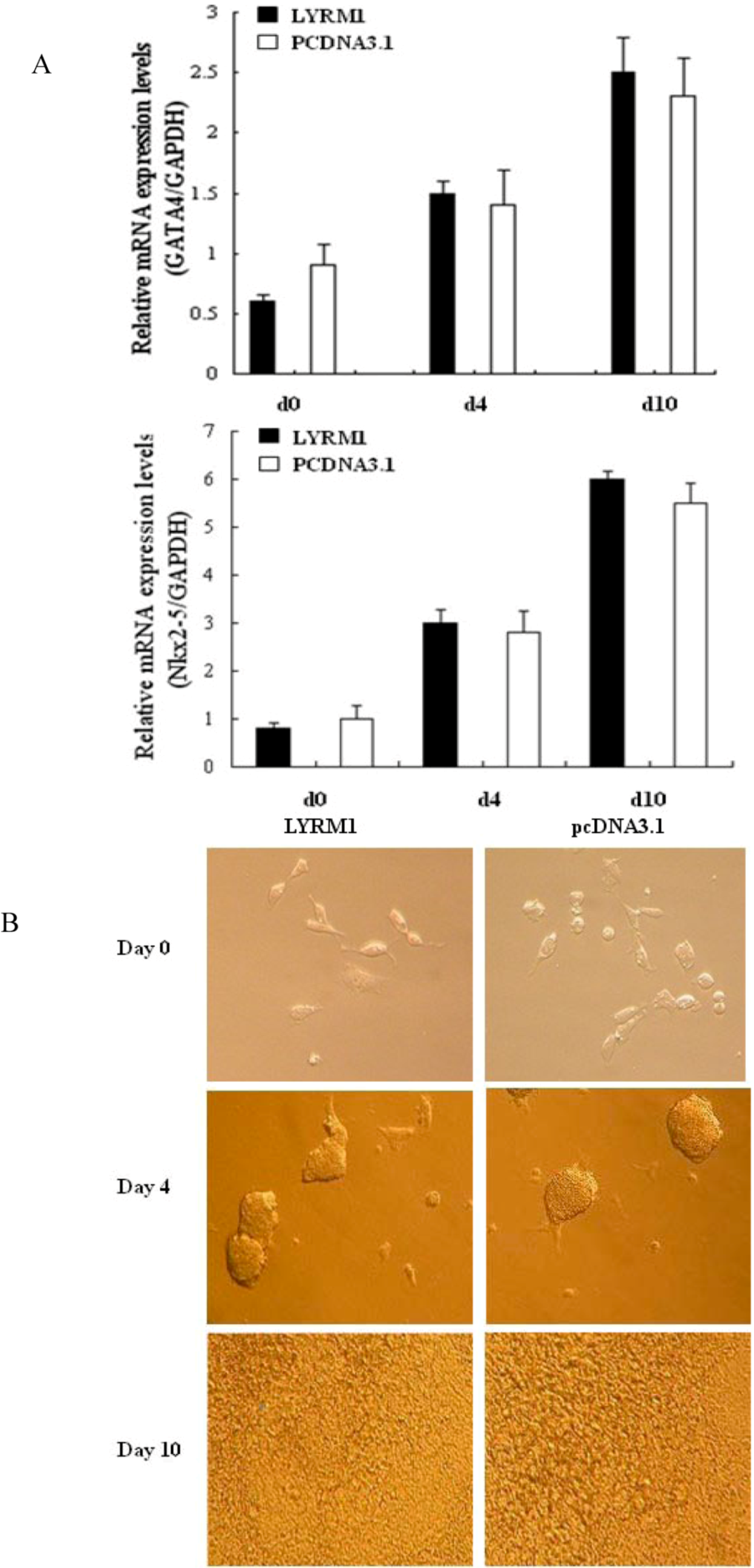
2.1. Effects of LYRM1 on cell differentiation
2.2. Effect of LYRM1 on cell proliferation


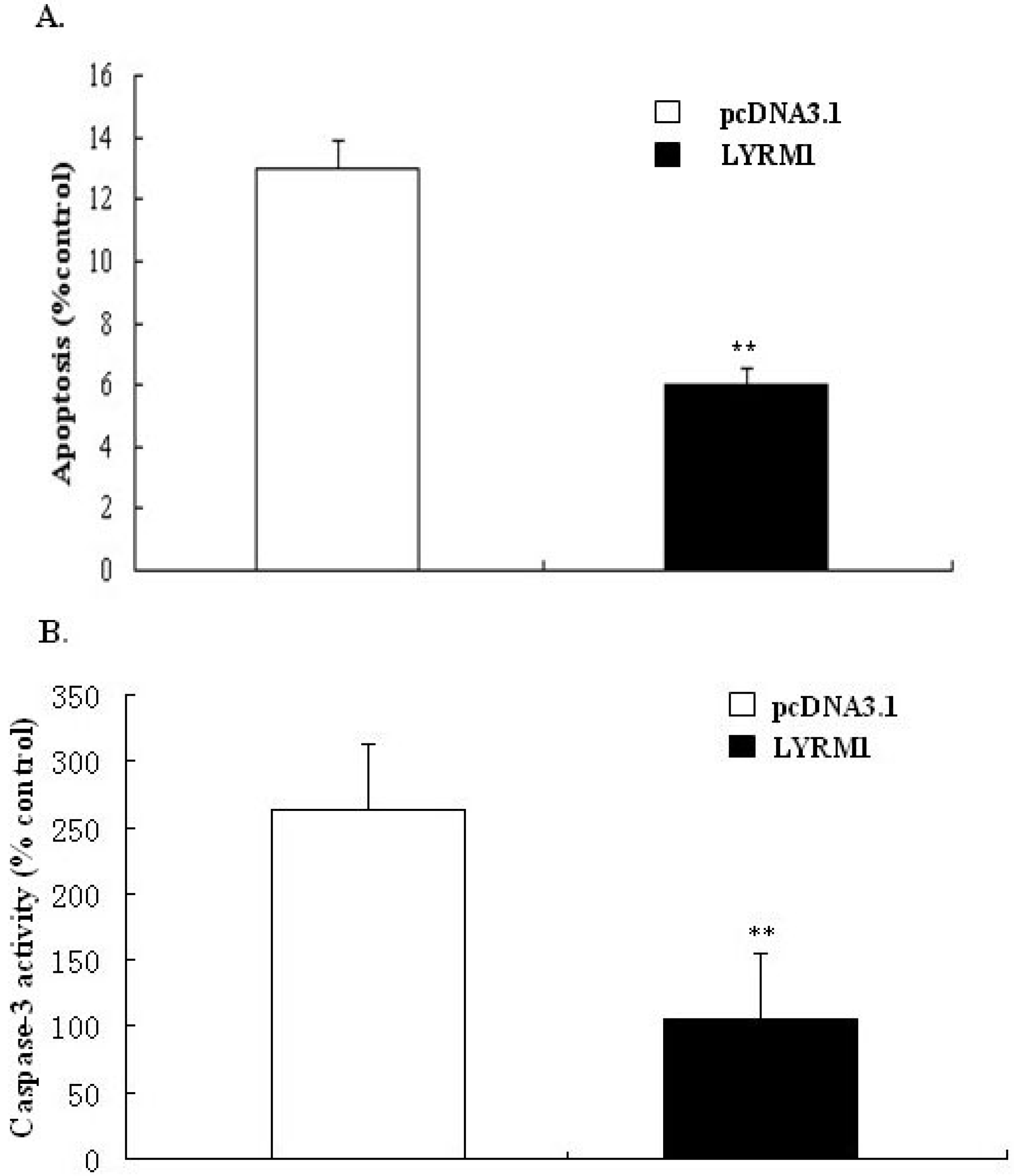
2.3. Effect of LYRM1 on cell apoptosis

2.4. Discussion
3. Experimental
3.1. Cell culture and differentiation of P19 cells
3.2. Quantitative real-time RT-PCR

{kind=link}
{kind=link}
{kind=link}
| Gene name | Product size | Reverse and forward primers (5'-3') |
| GATA4 | 136bp | F:5'-CCTGCGGCCTCTACATGA-3' |
| R:5'-AGGGTCTCACCAGCAGGA-3' | ||
| Nkx2-5 | 222bp | F:5'-CCTGCGGCCTCTACATGA-3' |
| R:5'-AGGGTCTCACCAGCAGGA-3' | ||
| GAPDH | 237bp | F:5'-TTCACCACCATGGAGAAGGC-3' |
| R:5'-GGCATGGACTGTGGTCATGA-3' |
3.3. Establishment of a stable cell line overexpressing LYRM1
3.4. MTT assay
3.5. Cell cycle assay
3.6. Apoptosis assay
3.7. Measurement of caspase-3 activity
3.8. Statistical analysis
4. Conclusions
Acknowledgements
- Sample Availability: Contact the authors.
References
- Sadowski, S.L. Congenital cardiac disease in the newborn infant: Past, present, and future. Crit. Care. Nurs. Clin. North Am. 2009, 21, 37–48. [Google Scholar] [CrossRef]
- Gorenflo, M.; Gu, H.; Xu, Z. Peri-operative pulmonary hypertension in paediatric patients: Current strategies in children with congenital heart disease. Cardiology 2010, 116, 10–17. [Google Scholar] [CrossRef]
- Huang, J.B.; Liu, Y.L.; Sun, P.W.; Lv, X.D.; Du, M.; Fan, X.M. Molecular mechanisms of congenital heart disease. Cardiovasc Pathol. 2010, 19, e183–e193. [Google Scholar] [CrossRef]
- Qiu, J.; Gao, C.L.; Zhang, M.; Chen, R.H.; Chi, X.; Liu, F.; Zhang, C.M.; Ji, C.B.; Chen, X.H.; Zhao, Y.P.; Li, X.N.; Tong, M.L.; Ni, Y.H.; Guo, X.R. LYRM1, a novel gene promotes proliferation and inhibits apoptosis of preadipocytes. Eur. J. Endocrinol. 2009, 160, 177–184. [Google Scholar]
- Bentham, J.; Bhattacharya, S. Genetic mechanisms controlling cardiovascular development. Ann. N Y Acad. Sci. 2008, 1123, 10–19. [Google Scholar]
- Joziasse, I.C.; van de Smagt, J.J.; Smith, K.; Bakkers, J.; Sieswerda, G.J.; Mulder, B.J.; Doevendans, P.A. Genes in congenital heart disease: atrioventricular valve formation. Basic Res. Cardiol. 2008, 103, 216–227. [Google Scholar] [CrossRef]
- Srivastava, D. Genetic regulation of cardiogenesis and congenital heart disease. Annu. Rev. Pathol. 2006, 1, 199–213. [Google Scholar] [CrossRef]
- Fiorina, P.; Corradi, D.; Pinelli, S.; Maestri, R.; Lagrasta, C.; Buscaglia, M.; Davalli, A.; Folli, F.; Astorri, E. Apoptotic/mytogenic pathways during human heart development. Int. J. Cardiol. 2004, 96, 409–417. [Google Scholar] [CrossRef]
- Heyden, M.A.; Kempen, M.J.; Tsuji, Y.; Rook, M.B.; Jongsma, H.J.; Opthof, T. P19 embryonal carcinoma cells: A suitable model system for cardiac electrophysiological differentiation at the molecular and functional level. Cardiovasc. Res. 2003, 58, 410–422. [Google Scholar] [CrossRef]
- Heyden, M.A.; Defize, L.H. Twenty one years of P19 cells: what an embryonal carcinoma cell line taught us about cardiomyocyte differentiation. Cardiovasc. Res. 2003, 58, 292–302. [Google Scholar] [CrossRef]
- Grepin, C.; Nemer, G.; Nemer, M. Enhanced cardiogenesis in embryonic stem cells overexpressing the GATA-4 transcription factor. Development 1997, 124, 2387–2395. [Google Scholar]
- Peterkin, T.; Gibson, A.; Loose, M.; Patient, R. The roles of GATA-4, -5 and -6 in vertebrate heart development. Semin. Cell Dev. Biol. 2005, 16, 83–94. [Google Scholar]
- Pikkarainen, S.; Tokola, H.; Kerkela, R.; Ruskoaho, H. GATA transcription factors in the developing and adult heart. Cardiovasc. Res. 2004, 63, 196–207. [Google Scholar] [CrossRef]
- Lints, T.J.; Parsons, L.M.; Hartley, L.; Lyons, I.; Harvey, R.P. 5: A novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993, 119, 419–431. [Google Scholar]
- Komuro, I.; Izumo, S. Csx: A murine homeobox-containing gene specifically expressed in the developing heart. Proc. Natl. Acad. Sci. USA 1993, 90, 8145–8149. [Google Scholar] [CrossRef]
- Ueyama, T.; Kasahara, H.; Ishiwata, T.; Nie, Q.; Izumo, S. Myocardin expression is regulated by Nkx2.5, and its function is required for cardiomyogenesis. Mol. Cell Biol. 2003, 23, 9222–9232. [Google Scholar]
- Durocher, D.; Charron, F.; Warren, R.; Schwartz, R.T.; Nemer, M. The cardiac transcription factors Nkx2-5 and GATA-4 are mutual cofactors. EMBO J. 1997, 16, 5687–5696. [Google Scholar] [CrossRef]
- Brown, CO., 3rd.; Chi, X.; Garcia-Gras, E.; Shirai, M.; Feng, X.H.; Schwartz, R.T. The cardiac determination factor, Nkx2-5, is activated by mutual cofactors GATA-4 and Smad1/4 via a novel upstream enhancer. J. Biol. Chem. 2004, 279, 10659–10669. [Google Scholar]
- Jiang, Y.; Drysdale, T.A.; Evans, T. A role for GATA-4/5/6 in the regulation of Nkx2.5 expression with implications for patterning of the precardiac field. Dev. Biol. 1999, 216, 57–71. [Google Scholar] [CrossRef]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutelingsperger, C. A novel assay for apoptosis: Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labeled annexin-V. J. Immunol. Methods 1995, 180, 39–52. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhu, C.; Liu, Y.-Q.; Chen, F.-K.; Hu, D.-L.; Yu, Z.-B.; Qian, L.-M. LYRM1, a Gene that Promotes Proliferation and Inhibits Apoptosis during Heart Development. Molecules 2010, 15, 6974-6982. https://doi.org/10.3390/molecules15106974
Zhu C, Liu Y-Q, Chen F-K, Hu D-L, Yu Z-B, Qian L-M. LYRM1, a Gene that Promotes Proliferation and Inhibits Apoptosis during Heart Development. Molecules. 2010; 15(10):6974-6982. https://doi.org/10.3390/molecules15106974
Chicago/Turabian StyleZhu, Chun, Yao-Qiu Liu, Fu-Kun Chen, De-Liang Hu, Zhang-Bin Yu, and Ling-Mei Qian. 2010. "LYRM1, a Gene that Promotes Proliferation and Inhibits Apoptosis during Heart Development" Molecules 15, no. 10: 6974-6982. https://doi.org/10.3390/molecules15106974