Small Interfering RNA Effectively Inhibits the Expression of SARS Coronavirus Membrane Gene at Two Novel Targeting Sites

Abstract
:1. Introduction
2. Results and Discussion
2.1. The selection of the targeted sites for M gene specific siRNAs
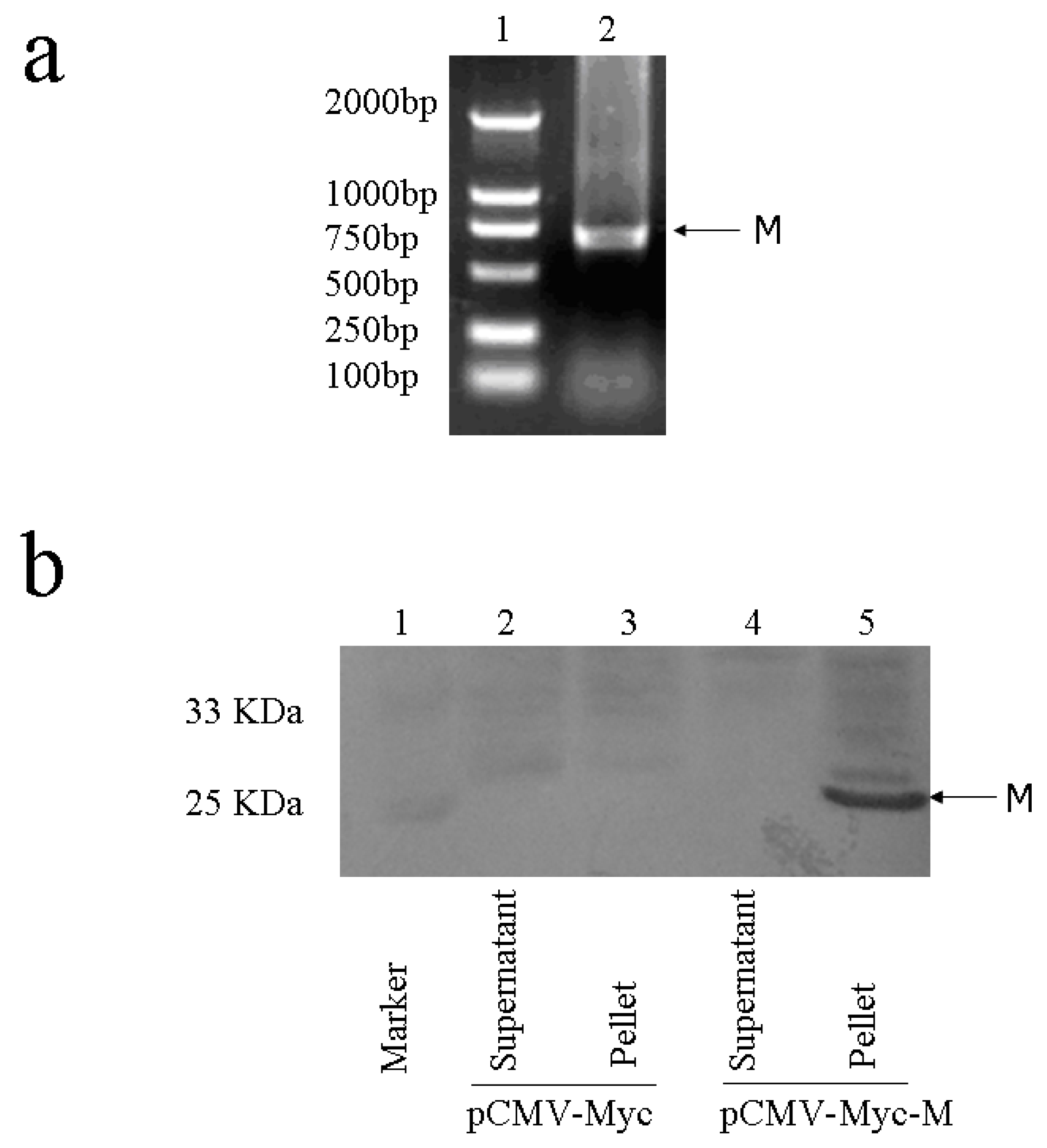
2.2. Cloning and expression analysis of M gene
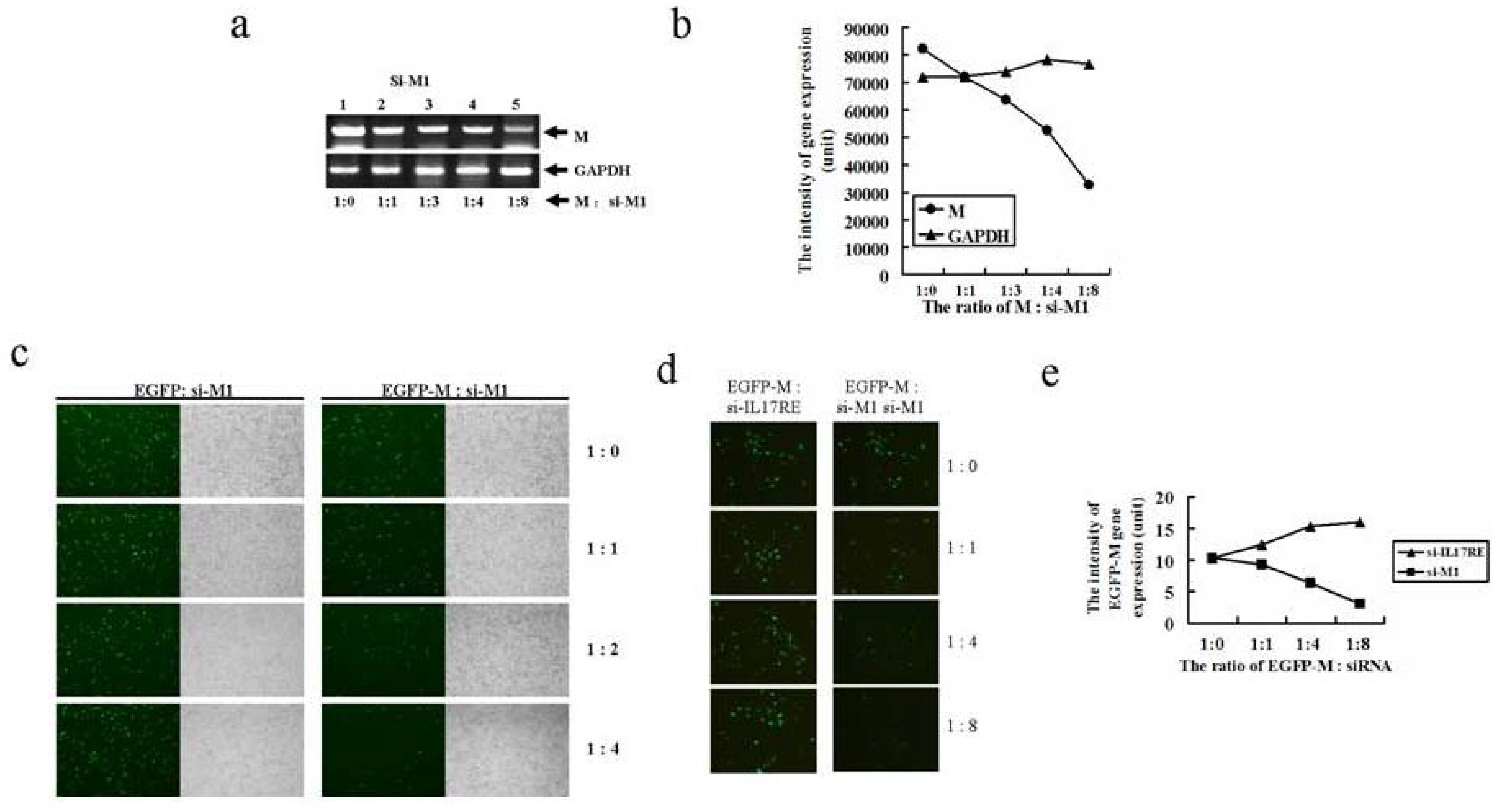
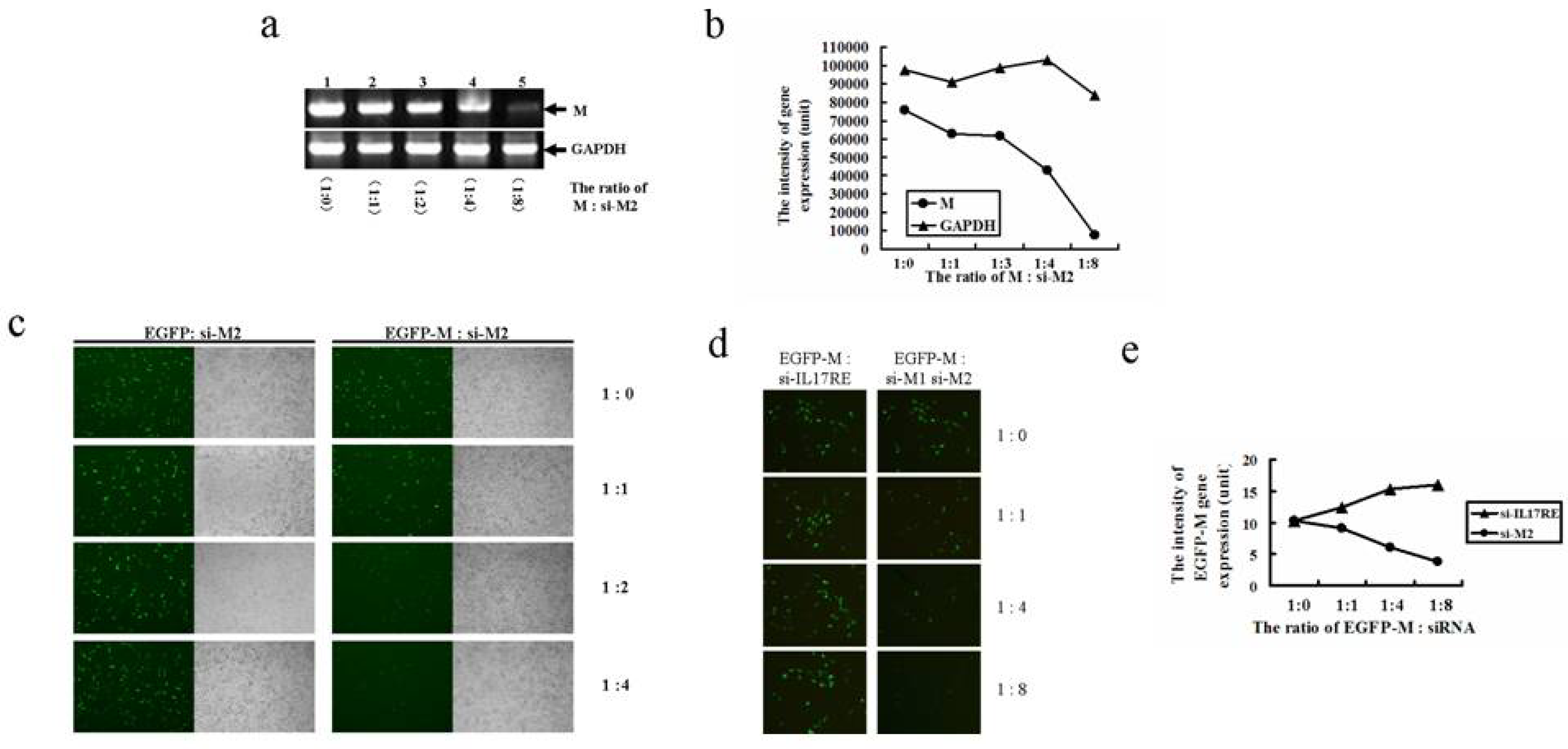
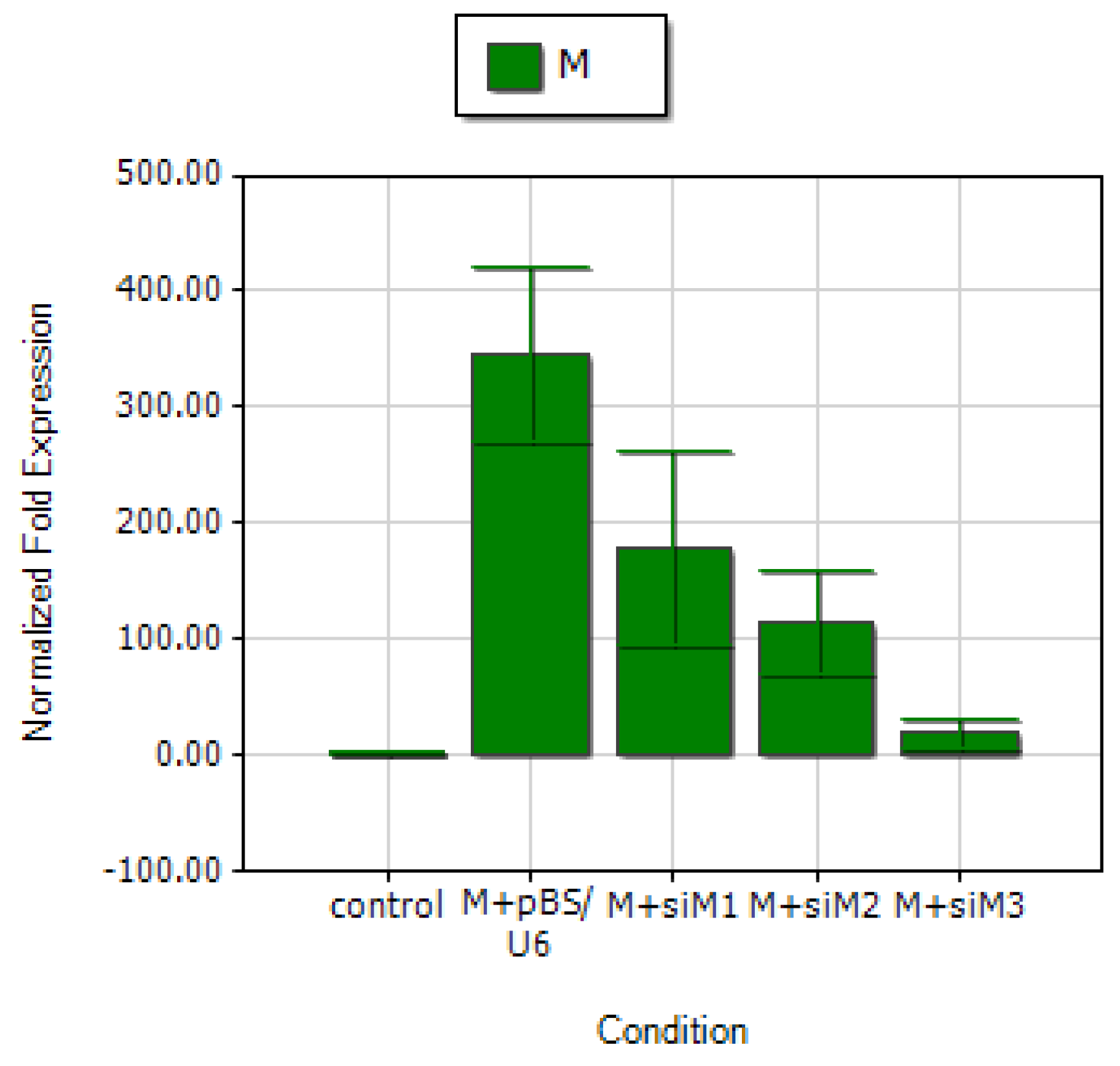
2.3. Two novel siRNAs effectively inhibits M gene expression
2.4. siRNA targeting the 3′ portion of M gene is a more potent inhibitor
2.5. M gene specific siRNAs effectively inhibited M gene mediated upregulation of interferon β gene expression
3. Experimental
3.1. Cell culture and antibodies
3.2. Plasmid construction
3.3. Reverse transcription-polymerase chain reaction (RT-PCR) and real time quantitative RT-PCR (qRT-PCR)
3.4. Transient transfection
3.5. Western blot analysis
3.6. Flow cytometric analysis
4. Conclusions
Acknowledgments
References
- Masters, P.S. The molecular biology of coronaviruses. Advan. Virus Res. 2006, 66, 193–292. [Google Scholar]
- Frieman, M.; Baric, R. Mechanisms of severe acute respiratory syndrome pathogenesis and innate immunomodulation. Microbiol. Mol. Biol. Rev. 2008, 72, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Dandekar, A.A. Immunopathogenesis of coronavirus infections: implications for SARS. Nat. Rev. Immunol. 2005, 5, 917–927. [Google Scholar] [PubMed]
- Narayanan, K.; Huang, C.; Makino, S. SARS coronavirus accessory proteins. Virus Res. 2008, 133, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Stertz, S.; Reichelt, M.; Spiegel, M.; Kuri, T.; Martinez-Sobrido, L.; Garcia-Sastre, A.; Weber, F.; Kochs, G. The intracellular sites of early replication and budding of SARS-coronavirus. Virology 2007, 361, 304–315. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, Z.Y.; Kong, W.P.; Nabel, G.J. Generation of synthetic severe acute respiratory syndrome coronavirus pseudoparticles: implications for assembly and vaccine production. J. Virol. 2004, 78, 12557–12565. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.K.; Chang, S.C.; Huang, C.C.; Lee, T.T.; Hsiao, C.W.; Kou, Y.H.; Chen, I.Y.; Chang, C.K.; Huang, T.H.; Chang, M.F. Assembly of severe acute respiratory syndrome coronavirus RNA packaging signal into virus-like particles is nucleocapsid dependent. J. Virol. 2005, 79, 13848–13855. [Google Scholar] [CrossRef] [PubMed]
- Siu, Y.L.; Teoh, K.T.; Lo, J.; Chan, C.M.; Kien, F.; Escriou, N.; Tsao, S.W.; Nicholls, J.M.; Altmeyer, R.; Peiris, J.S.; Bruzzone, R.; Nal, B. The M, E, and N structural proteins of the severe acute respiratory syndrome coronavirus are required for efficient assembly, trafficking, and release of virus-like particles. J. Virol. 2008, 82, 11318–11330. [Google Scholar] [CrossRef] [PubMed]
- McBride, C.E.; Li, J.; Machamer, C.E. The cytoplasmic tail of the severe acute respiratory syndrome coronavirus spike protein contains a novel endoplasmic reticulum retrieval signal that binds COPI and promotes interaction with membrane protein. J. Virol. 2007, 81, 2418–2428. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Ye, L.; Timani, K.A.; Li, S.; Zen, Y.; Zhao, M.; Zheng, H.; Wu, Z. Peptide domain involved in the interaction between membrane protein and nucleocapsid protein of SARS-associated coronavirus. J. Biochem. Mol. Biol. 2005, 38, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wu, D.; Shen, C.; Chen, K.; Shen, X.; Jiang, H. Severe acute respiratory syndrome coronavirus membrane protein interacts with nucleocapsid protein mostly through their carboxyl termini by electrostatic attraction. Int. J. Biochem. Cell Biol. 2006, 38, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Gao, J.; Zheng, H.; Li, B.; Kong, L.; Zhang, Y.; Wang, W.; Zeng, Y.; Ye, L. The membrane protein of SARS-CoV suppresses NF-kappaB activation. J. Med. Virol. 2007, 79, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.L.; Kok, K.H.; Ng, M.H.; Poon, V.K.; Yuen, K.Y.; Zheng, B.J.; Jin, D.Y. Severe acute respiratory syndrome coronavirus M protein inhibits type I interferon production by impeding the formation of TRAF3.TANK.TBK1/IKKepsilon complex. J. Biol. Chem. 2009, 284, 16202–16209. [Google Scholar] [CrossRef] [PubMed]
- Li, B.J.; Tang, Q.; Cheng, D.; Qin, C.; Xie, F.Y.; Wei, Q.; Xu, J.; Liu, Y.; Zheng, B.J.; Woodle, M.C.; Zhong, N.; Lu, P.Y. Using siRNA in prophylactic and therapeutic regimens against SARS coronavirus in Rhesus macaque. Nat. Med. 2005, 11, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Qin, Z.L.; Zhao, P.; Cao, M.M.; Qi, Z.T. siRNAs targeting terminal sequences of the SARS-associated coronavirus membrane gene inhibit M protein expression through degradation of M mRNA. J. Virol. Methods 2007, 145, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Yang, D.H.; Xiong, J.; Jia, J.; Huang, B.; Jin, Y.X. Inhibition of genes expression of SARS coronavirus by synthetic small interfering RNAs. Cell Res. 2005, 15, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, M.; Zhao, W.; Zhang, J.; Zhang, X.; Wang, K.; Gu, C.; Wu, K.; Li, Y.; Zheng, C.; Xiao, G.; Yan, H.; Guo, D.; Tien, P.; Wu, J. Isolation of virus from a SARS patient and genome-wide analysis of genetic mutations related to pathogenesis and epidemiology from 47 SARS-CoV isolates. Virus Genes 2005, 30, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.J.; Guan, Y.; Tang, Q.; Du, C.; Xie, F.Y.; He, M.L.; Chan, K.W.; Wong, K.L.; Lader, E.; Woodle, M.C.; Lu, P.Y.; Li, B.; Zhong, N. Prophylactic and therapeutic effects of small interfering RNA targeting SARS-coronavirus. Antivir. Ther. 2004, 9, 365–374. [Google Scholar] [PubMed]
- Lee, Y.N.; Chen, L.K.; Ma, H.C.; Yang, H.H.; Li, H.P.; Lo, S.Y. Thermal aggregation of SARS-CoV membrane protein. J. Virol. Methods 2005, 129, 152–161. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Bowie, A.G. The family of five: TIR-domain-containing adaptors in Toll-like receptor signalling. Nat. Rev. Immunol. 2007, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Sui, G.; Soohoo, C.; Affarel, B.; Gay, F.; Shi, Y.; Forrester, W.C. A DNA vector-based RNAi technology to suppress gene expression in mammalian cells. Proc. Natl. Acad. Sci. USA 2002, 99, 5515–5520. [Google Scholar] [CrossRef] [PubMed]
- Li, T.S.; Li, X.N.; Chang, Z.J.; Fu, X.Y.; Liu, L. Identification and functional characterization of a novel interleukin 17 receptor: a possible mitogenic activation through ras/mitogen-activated protein kinase signaling pathway. Cell. Signal. 2006, 18, 1287–1298. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the plasmids are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene name | GenBank ID | Forward primer | Reverse primer | Size of product (bp) |
|---|---|---|---|---|
| β-actin† GAPDH INFβ SARS-M † β-actin-R†† SARS-M-R†† | BC009275 NM_002046 NM_002176 AY278491 BC009275 AY278491 | 5′-cacactgtgcccatctacga-3′ 5’-tcttcaccaccatggagaag-3’ 5′-atgaccaacaagtgtctcct-3′ 5′-tatagaattctggcagacaacggtactatt-3′ 5′-tccatcatgaagtgtgacgt-3′ 5′-tgctgtgatcattcgtggtc-3′ | 5′-ctgcttgctgatccacatct-3′ 5’-ctgcttcaccaccttcttga-3’ 5′-ttcagtttcggaggtaacct-3′ 5′-tataggtaccgtcacttactgtactagcaaagc-3′ 5′-ctcaggaggagcaatgatct-3′ 5′-tacggtagcggttgtatgca-3′ | 600 489 564 686 161 178 |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, Y.; Cao, Y.-L.; Yang, F.; Zhang, Y.; Wang, S.-H.; Liu, L. Small Interfering RNA Effectively Inhibits the Expression of SARS Coronavirus Membrane Gene at Two Novel Targeting Sites. Molecules 2010, 15, 7197-7207. https://doi.org/10.3390/molecules15107197
Wang Y, Cao Y-L, Yang F, Zhang Y, Wang S-H, Liu L. Small Interfering RNA Effectively Inhibits the Expression of SARS Coronavirus Membrane Gene at Two Novel Targeting Sites. Molecules. 2010; 15(10):7197-7207. https://doi.org/10.3390/molecules15107197
Chicago/Turabian StyleWang, Ying, Ying-Li Cao, Fan Yang, Yun Zhang, Shu-Hui Wang, and Li Liu. 2010. "Small Interfering RNA Effectively Inhibits the Expression of SARS Coronavirus Membrane Gene at Two Novel Targeting Sites" Molecules 15, no. 10: 7197-7207. https://doi.org/10.3390/molecules15107197