Emodin-8-O-β-D-Glucoside from Polygonum Amplexicaule D. Don var. Sinense Forb. Promotes Proliferation and Differentiation of Osteoblastic MC3T3-E1 Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Result and Discussion
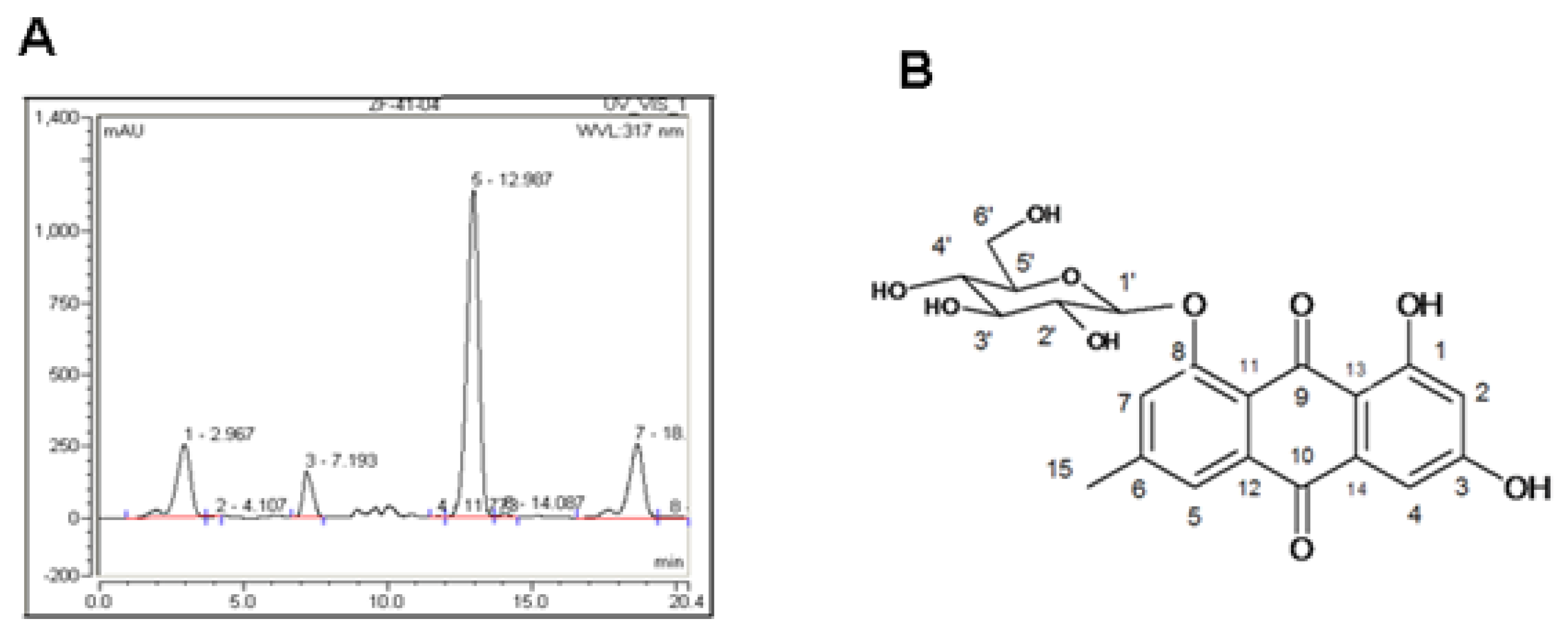
2.1. Indentification of PAF Compounds
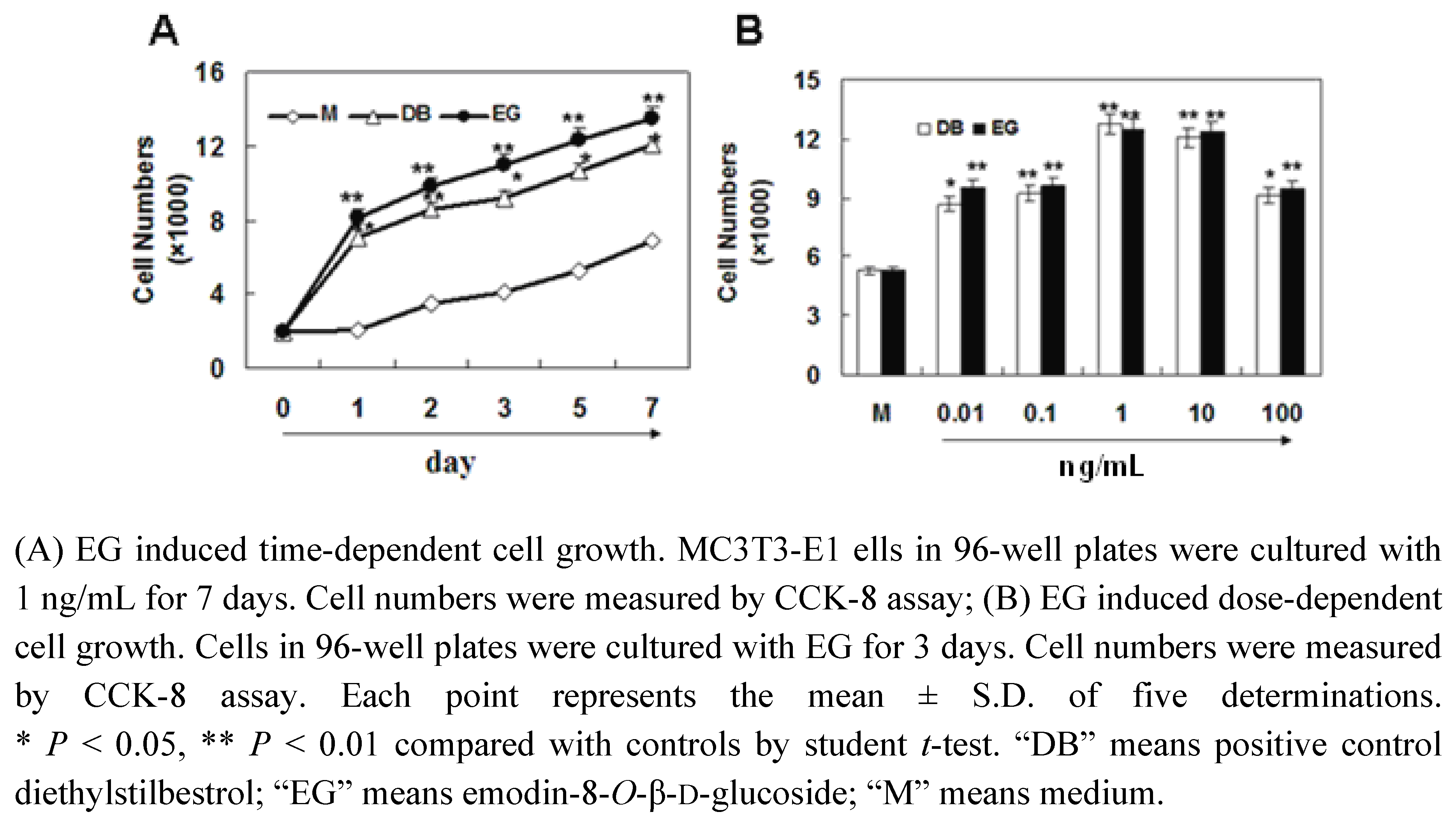
2.2. The Effect of EG on Proliferation of MC3T3-E1 Cells
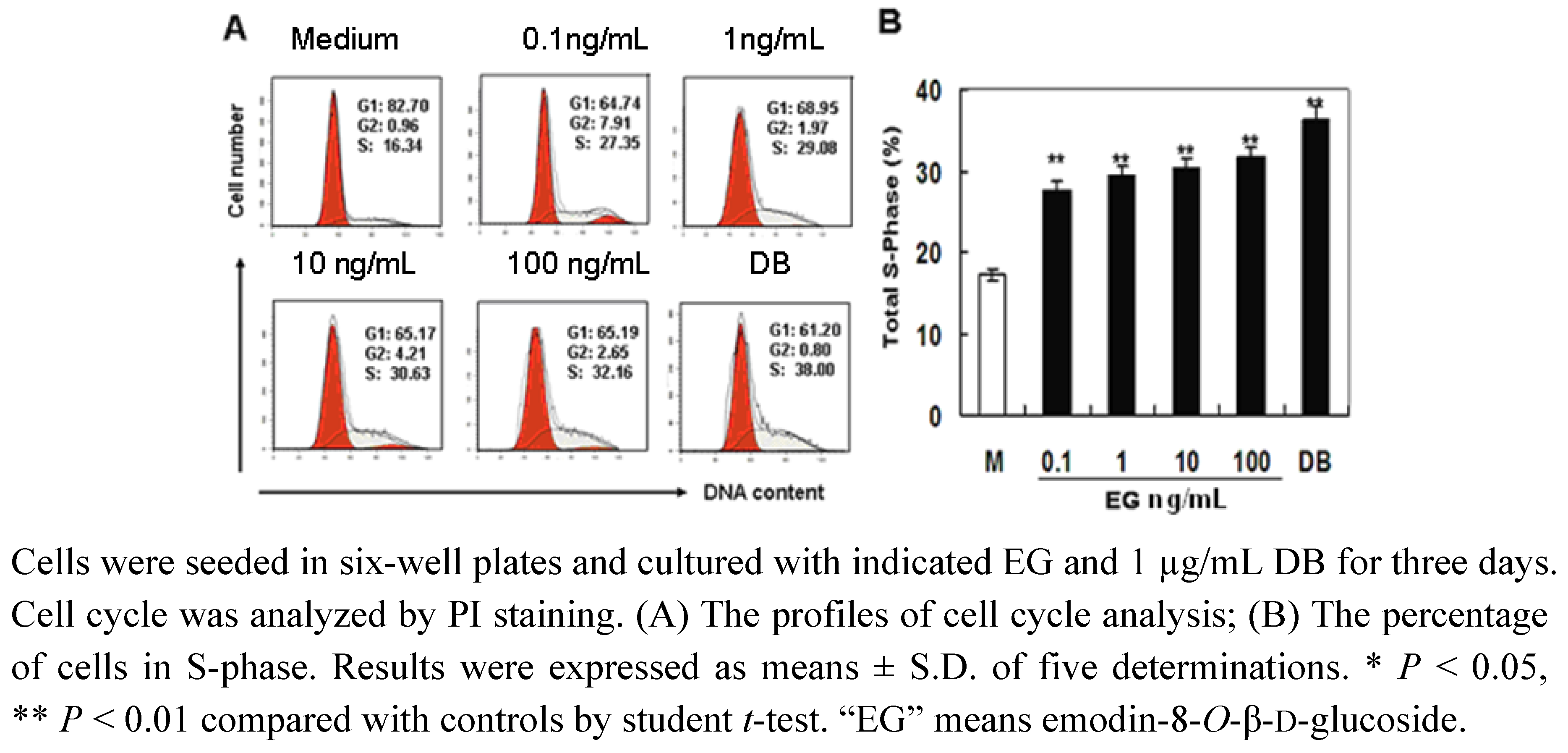
2.3. The Effect of EG on Cell Cycle Progression
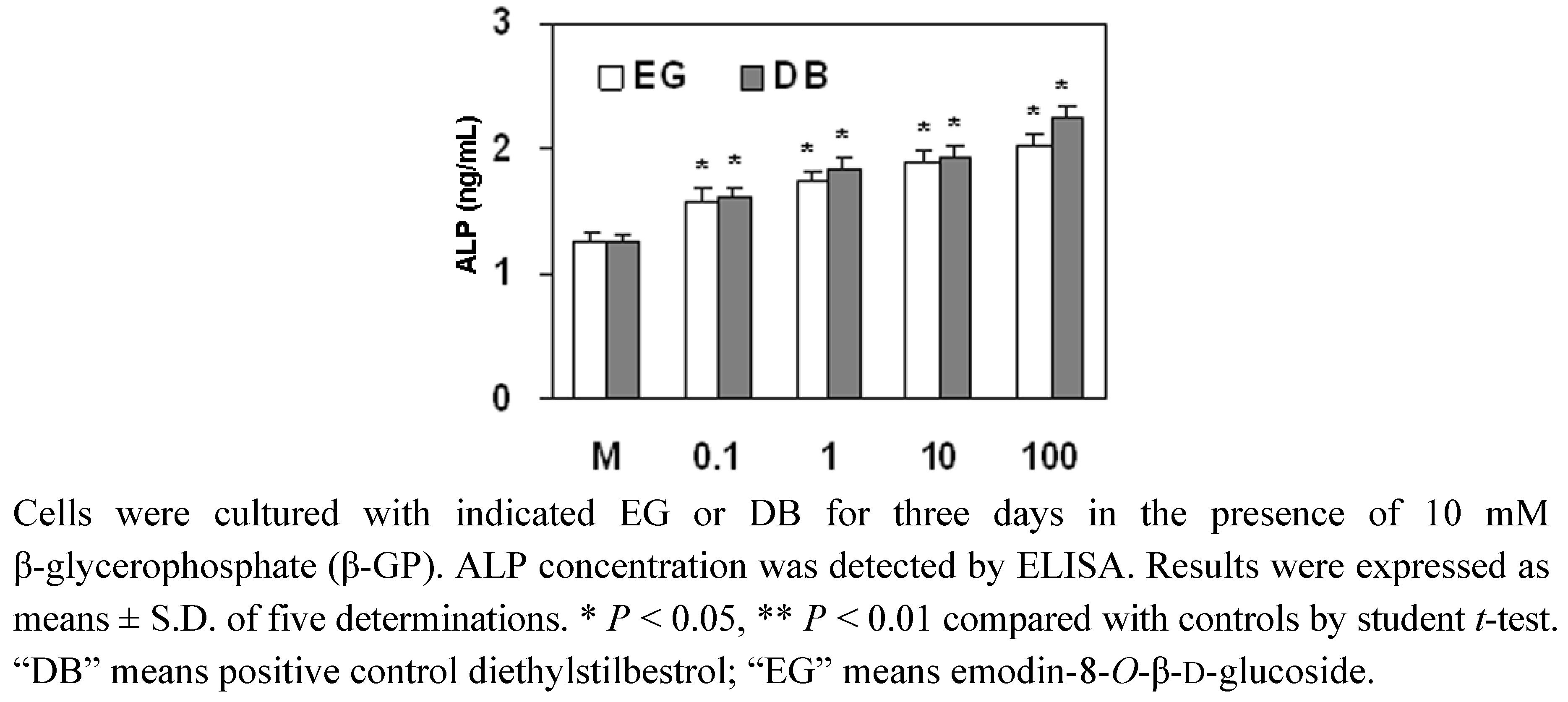
2.4. The Effect of EG on ALP Expression in MC3T3-E1 Cells
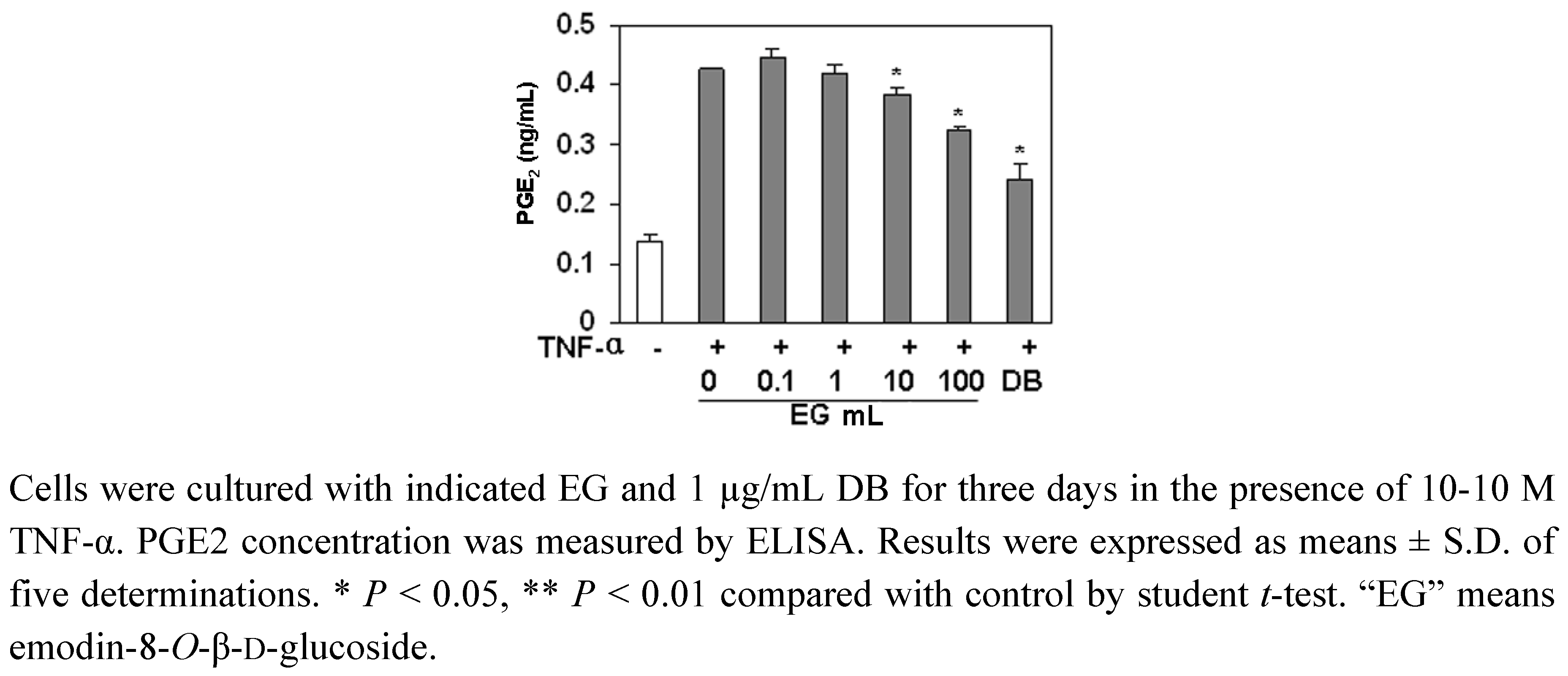
2.5. The Effect of EG on PGE2 Production in MC3T3-E1 Cells
3. Experimental
3.1. Materials
3.2. Plant Material
3.3. Extraction and Isolation Procedures
3.4. Sample Preparation
3.5. Cell Culture
3.6. Cell Proliferative CCK-8 Assay
3.7. Cell Cycle Analysis
3.8. Alkaline Phosphatase (ALP) Activity Assay
3.9. Prostaglandin (PG) E2 Assay
3.10. Statistics
4. Conclusions
Acknowledgements
References and Notes
- Brixen, K.T.; Christensen, P.M.; Ejersted, C.; Langdahl, B.L. Teriparatide (biosythetic human parathyroid hormone 1-34): a new paradign in the treatment of osteoporosis. Basic Clin. Pharmacol. Toxicol. 2004, 94, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, D.; Ma, H.; Liu, J. Neuroprotective effects of emodin-8-O-beta-d-glucoside in vivo and in vitro. Eur. J. Pharmacol. 2007, 577, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Quarlers, J.D.; Yohay, D.A.; Lever, L.W.; Caton, R.; Wenstrup, R.J. Distinct proliferative and differentiated stages of mouse MC3T3-E1 cells in culture: an in vitro model of osteoblast development. J. Bone Miner. Res. 1999, 27, 683–689. [Google Scholar]
- Zhao, J.; Ohba, S.; Komiyama, Y.; Shinkai, M.; Chung, U.I.; Nagamune, T. Icariin: a potential osteoinductive compound for bone tissue engineering. Tissue Eng. Pt. A 2010, 16, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.Y.; Lau, K.S.; Jiang, B.; Kennelly, E.J.; Kronenberg, F.; Kung, A.W. Ethanolic extract of Actaea racemosa (black cohosh) potentiates bone nodule formation in MC3T3-E1 preosteoblast cells. Bone 2008, 43, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.W.; Suh, S.J.; Lee, T.K.; Ha, K.T.; Kim, J.K.; Kim, K.H.; Kim, D.I.; Jeon, J.H.; Moon, T.C.; Kim, C.H. Effect of safflower seeds supplementation on stimulation of the proliferation, differentiation and mineralization of osteoblastic MC3T3-E1 cells. J. Ethnopharmacol. 2008, 115, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.K.; Kim, K.S.; Byun, Y.S.; Suh, S.J.; Jim, U.H.; Kim, K.H.; Lee, I.S.; Kim, C.H. Effects of Ulmus davidiana planch on mineralization, bone morphogenetic protein-2, alkaline phosphatase, type I collagen, and collagenase-1 in bone cells. In vitro Cell Dev. Biol. Anim. 2006, 42, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.C.; Lee, J.W.; Yoon, C.H.; Kim, H.M.; Kim, C.H. Drynariae Rhizoma promotes osteoblast differentiation and mineralization in MC3T3-E1 cells through regulation of bone morphogenetic protein-2, alkaline phosphatase, type I collagen and collagenase-1. Toxicol. In Vitro 2004, 18, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.C; Lee, J.W.; Yoon, C.H.; Lee, Y.C.; Chung, K.H.; Kim, M.G.; Kim, C.H. Stimulative effects of Drynariae Rhizoma extracts on the proliferation and differentiation of osteoblastic MC3T3-E1 cells. J. Ethnopharmacol. 2005, 9, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.S.; Koh, G.; Park, C.Y.; Woo, J.T.; Kim, S.W.; Kim, J.W.; Park, I.K.; Kim, Y.S. Soybean isoflavones inhibit tumor necrosis factor-alpha-induced apoptosis and the production of interleukin-6 and prostaglandin E2 in osteoblastic cells. Phytochemistry 2003, 63, 209–215. [Google Scholar] [PubMed]
- Choi, E.M.; Suh, K.S.; Kim, Y.S.; Choue, R.W.; Koo, S.J. Soybean ethanol extract increases the function of osteoblastic MC3T3-E1 cells. Phytochemistry 2001, 56, 733–739. [Google Scholar] [CrossRef]
- Hu, Y.M.; Liu, C.; Cheng, K.W.; Sung, H.H.Y.; Williams, L.D.; Yang, Z.L.; Ye, W.C. Sesquiterpenoids from Homalomen occulta affect osteoblast proliferation, differentiation and mineralization in vitro. Phytochemistry 2008, 69, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wang, M.; Rosen, R.T.; Ho, C.T. 2, 2′-Dipheny-1-picryl-hydrazyl-radical- scavenging active components from Polygonum multiflorum Thunb. J. Agric. Food Chem. 1999, 47, 2226–2228. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, R.; Xie, C.; Wei, X.; Zhang, M.; Zhang, X.; Flick, L.M.; Schwarz, E.M.; O’Keefe, R.J. PGE2 signaling through the EP4 receptor on fibroblasts upregulates RANKL and stimulates osteolysis. J. Bone Miner. Res. 2009, 24, 1753–1762. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Gao, Y.H. Inhibitory effect of genistein on bone resorption in tissue culture. Biochem. Pharmacol. 1998, 55, 71–76. [Google Scholar] [CrossRef]
- Mizutani, K.; Ikeda, K.; Kawai, Y.; Yamori, Y. Resveratrol stimulates the proliferation and differentiation of osteoblastic MC3T3-E1 cells. Biochem. Biophys. Res. Commun. 1998, 253, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.O.; Kook, S.H.; Choi, K.C.; Jang, Y.S.; Jeon, Y.M.; Kim, J.G.; Lee, K.Y.; Kim, J.; Chung, M.S.; Chung, G.H.; Lee, J.C. Quercetin, a bioflavonoid, accelerates TNF-alpha-induced growth inhibition and apoptosis in MC3T3-E1 osteoblastic cells. Eur. J. Pharmacol. 2006, 529, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, H.; Zhou, P.; Kim, H.J.; Novack, D.V.; Ross, F.P.; Teitelbaum, S.L. M-CSF mediates TNF-induced inflammatory osteolysis. J. Clin. Invest. 2005, 115, 3418–3427. [Google Scholar] [CrossRef] [PubMed]
- Kwan, T.S; Padrines, M.; Théoleyre, S.; Heymann, D.; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar]
- Igarashi, K.; Hirafuji, M.; Adachi, H.; Shinoda, H.; Mitani, H. Effects of bisphosphonates on alkaline phosphatase activity, mineralization, and prostaglandin E2 synthesis in the clonal osteoblast-like cell line MC3T3-E1. Prostaglandins Leukot Essent Fatty Acids 1997, 56, 121–125. [Google Scholar] [CrossRef]
- Kim, H.S.; Shin, T.H.; Yang, S.R.; Seo, M.S.; Kim, D.J.; Kang, S.K.; Park, J.H.; Kang, K.S. Implication of NOD1 and NOD2 for the differentiation of multipotent mesenchymal stem cells derived from human umbilical cord blood. PLoS One 2010, 10, e15369. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, I.; Yamaguchi, T.; Yano, S.; Yamauchi, M.; Sugimoto, T. Metformin enthances the differentiation and mineralization of osteoblastic MC3T3-E1 cells via AMP kinase activation as well as eNOS and BMP-2 expression. Biochem. Biophys. Res. Commun. 2008, 375, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Pozharski, E. Fluorometric analysis of individual cationic lipid-DNA complexes. Methods Mol. Biol. 2010, 606, 385–391. [Google Scholar] [PubMed]
- Choi, E.M. The licorice root derived isoflavan glabridin increases the function of osteoblastic MC3T3-E1 cells. Biochem. Pharmacol. 2005, 70, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Amos, A.F.; Trevor, W.S.; Robert, A.S. Responses of differentiated MC3T3-E1 osteoblast-like cells to reactive oxygen species. Eur. J. Pharmacol. 2008, 587, 35–41. [Google Scholar]
- Kajii, T.; Suzuki, K.; Yoshikawa, M.; Imai, T.; Matsumoto, A.; Nakamura, S. Long-term effects of prostaglandin E2 on the mineralization of a clonal osteoblastic cell line (MC3T3-E1). Arch. Oral Biol. 1999, 44, 233–241. [Google Scholar] [CrossRef]





© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Xiang, M.-X.; Xu, Z.; Su, H.-W.; Hu, J.; Yan, Y.-J. Emodin-8-O-β-D-Glucoside from Polygonum Amplexicaule D. Don var. Sinense Forb. Promotes Proliferation and Differentiation of Osteoblastic MC3T3-E1 Cells. Molecules 2011, 16, 728-737. https://doi.org/10.3390/molecules16010728
Xiang M-X, Xu Z, Su H-W, Hu J, Yan Y-J. Emodin-8-O-β-D-Glucoside from Polygonum Amplexicaule D. Don var. Sinense Forb. Promotes Proliferation and Differentiation of Osteoblastic MC3T3-E1 Cells. Molecules. 2011; 16(1):728-737. https://doi.org/10.3390/molecules16010728
Chicago/Turabian StyleXiang, Mei-Xian, Zong Xu, Han-Wen Su, Jinyue Hu, and Yun-Jun Yan. 2011. "Emodin-8-O-β-D-Glucoside from Polygonum Amplexicaule D. Don var. Sinense Forb. Promotes Proliferation and Differentiation of Osteoblastic MC3T3-E1 Cells" Molecules 16, no. 1: 728-737. https://doi.org/10.3390/molecules16010728