Enhancement of Diepoxin ζ Production by Yeast Extract and Its Fractions in Liquid Culture of Berkleasmium-Like Endophytic Fungus Dzf12 from Dioscorea zingiberensis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
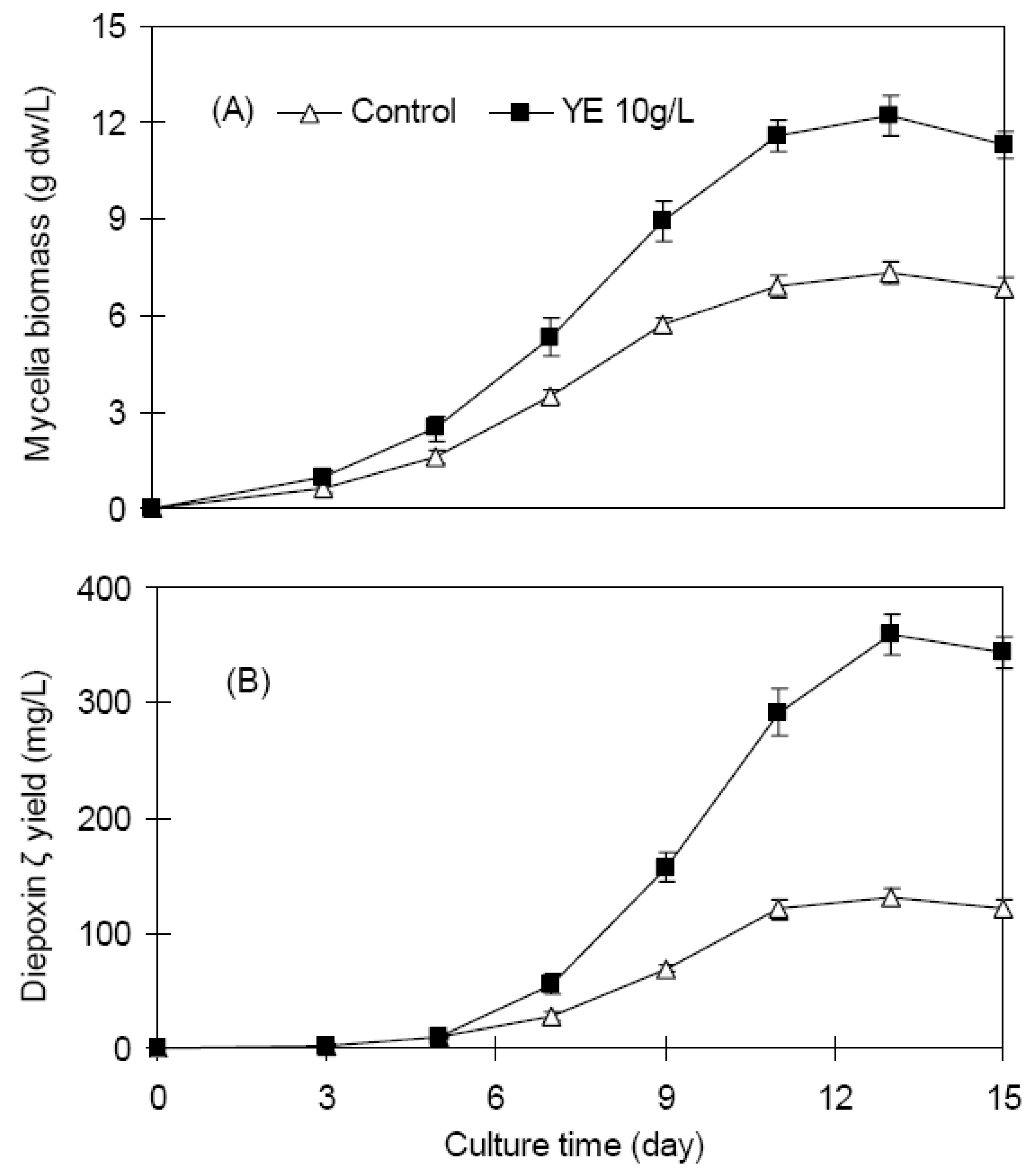
2.1. Mycelia growth and diepoxin ζ accumulation of endophyte Dzf12 in batch culture
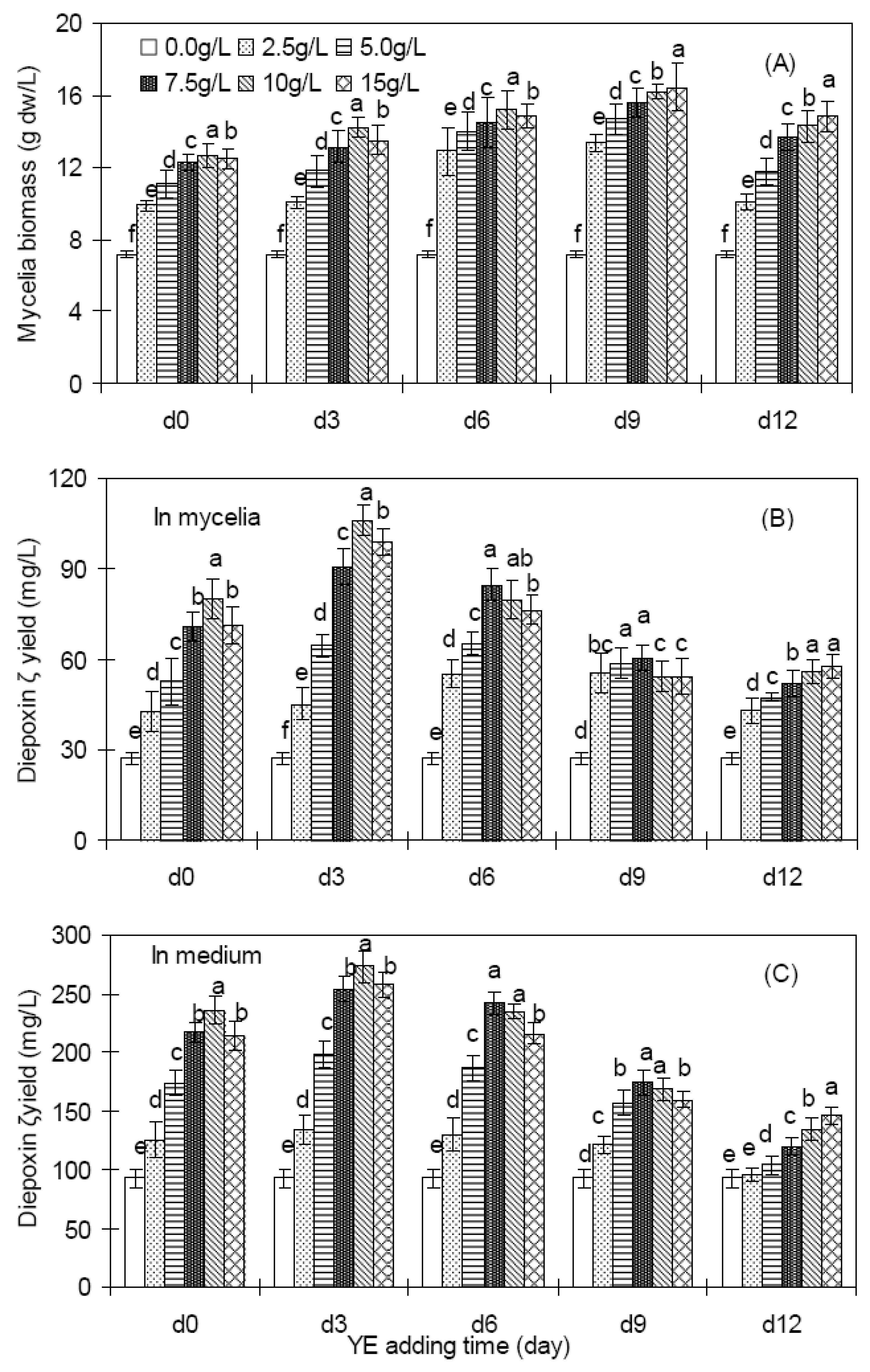
2.2. Effects of YE on mycelia growth and diepoxin ζ production of endophyte Dzf12
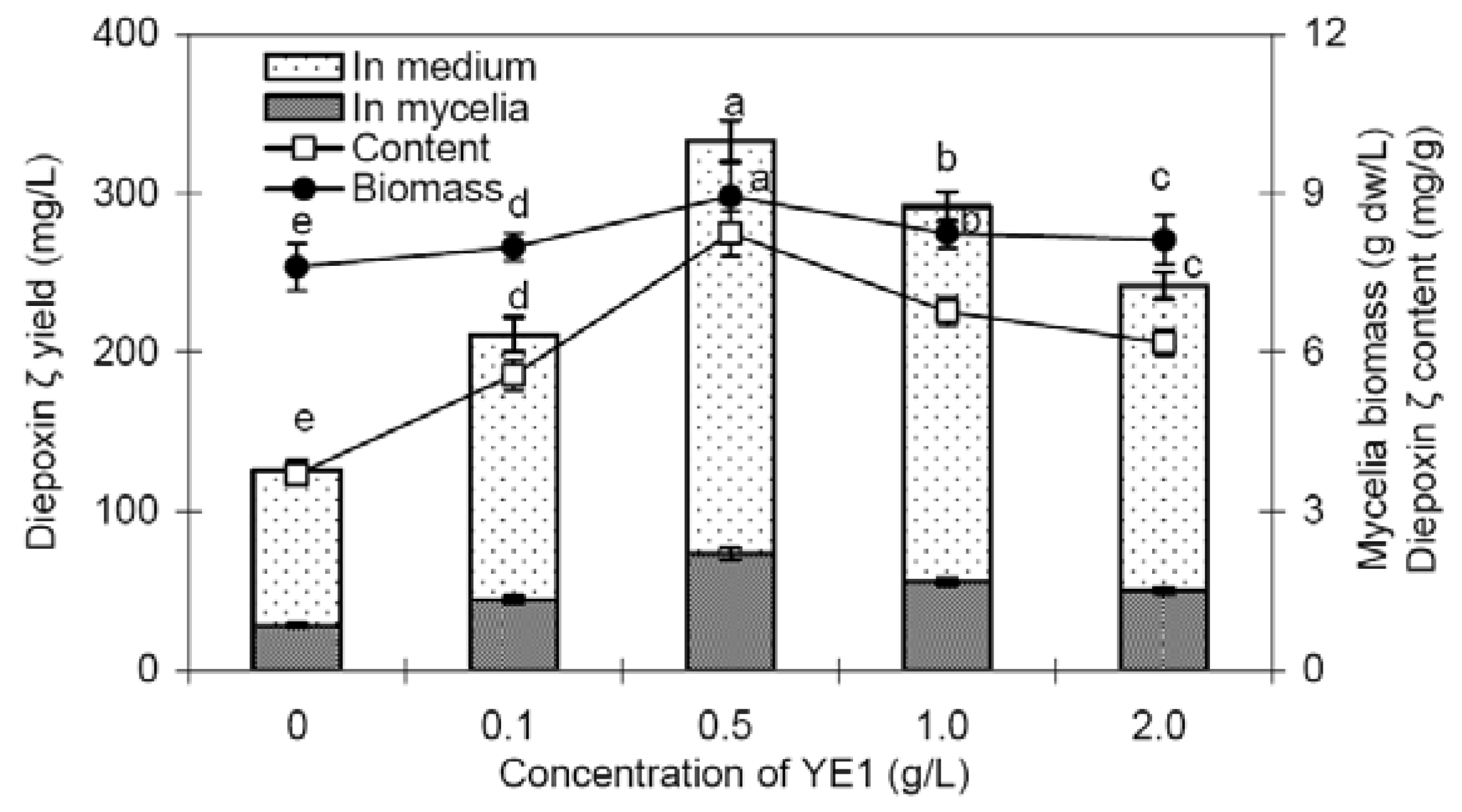
2.3. Effects of fractions YE1 and YE2 on mycelia growth and diepoxin ζ production of endophyte Dzf12
3. Experimental
3.1. Endophytic fungus and culture conditions
3.2. Preparation and application of YE and its fractions
3.3. Measurement of biomass and diepoxin ζ content
3.4. Statistical analysis
4. Conclusions
Acknowledgements
References
- Strobel, G.; Daisy, B.; Castillo, U.; Harper, J. Natural products from endophytic microorganisms. J. Nat. Prod. 2004, 67, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Gunatilaka, A.A.L. Natural products from plant-associated microorganism: distribution, structural diversity, bioactivity, and implications of their occurrence. J. Nat. Prod. 2006, 69, 509–526. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, C.; Cabrera, R.; Reina, M.; Gonzalez-Coloma, A. Fungal endophytes and their role in plant protection. Curr. Org. Chem. 2007, 11, 707–720. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, L.; Li, L.; Zheng, C.; Guo, L.; Li, W.; Sun, P.; Qin, L. Recent developments and future prospects of antimicrobial metabolites produced by endophytes. Microbiol. Res. 2010, 165, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.W.; Song, Y.C.; Tan, R.X. Biology and chemistry of endophytes. Nat. Prod. Rep. 2006, 23, 753–771. [Google Scholar] [CrossRef] [PubMed]
- Strobel, G. Plant-associated microorganisms (endophytes) as a new source of bioactive natural products. Med. Plant Biotechnol. 2007, 1, 49–72. [Google Scholar]
- Suryanarayanan, T.S.; Thirunavukkarasu, N.; Govindarajulu, M.B.; Sasse, F.; Jansen, R.; Murali, T.S. Fungal endophytes and bioprospecting. Fungal Biol. Rev. 2009, 23, 9–19. [Google Scholar] [CrossRef]
- Verma, V.C.; Kharwar, R.N.; Strobel, G.A. Chemical and functional diversity of natural products from plant associated endophytic fungi. Nat. Prod. Commun. 2009, 4, 1511–1532. [Google Scholar] [PubMed]
- Zhou, L.; Zhao, J.; Xu, L.; Huang, Y.; Ma, Z.; Wang, J.; Jiang, W. Antimicrobial compounds produced by plant endophytic fungi. In Fungicides: Chemistry, Environmental Impact and Health Effects; De Costa, P., Bezerra, P., Eds.; Nova Science Publishers: New York, NY, USA, 2009; pp. 91–119. [Google Scholar]
- Zhou, L.; Zhao, J.; Shan, T.; Cai, X.; Peng, Y. Spirobisnaphthalenes from fungi and their biological activities. Mini-Rev. Med. Chem. 2010, 10, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Ogishi, H.; Chiba, N.; Mikawa, T.; Sakaki, T.; Miyaji, S.; Sezaki, M. Novel antibiotic MK 3018 substance-useful as antimicrobial agent. Jpn. Pat. 01294686, 28 November 1989. [Google Scholar]
- Schlingmann, G.; West, R.R.; Milne, L.; Pearce, C.J.; Carter, G.T. Diepoxins, novel fungal metabolites with antibiotic activity. Tetrahedron Lett. 1993, 34, 7225–7228. [Google Scholar] [CrossRef]
- Chu, M.; Truumees, I.; Patel, M.G.; Gullo, V.P.; Blood, C.; King, I.; Pai, J.K.; Puar, M.S. A novel class of antitumor metabolites from the fungus Nattrassia mangiferae. Tetrahedron Lett. 1994, 35, 1343–1346. [Google Scholar] [CrossRef]
- Krohn, K.; Michel, A.; Florke, U.; Aust, H.-J.; Draeger, S.; Schulz, B. Palmarumycins C1-C16 from Coniothyrium sp.: isolation, structure elucidation and biological activity. Liebigs Ann. Chem. 1994, 11, 1099–1108. [Google Scholar] [CrossRef]
- Petersen, F.; Moerker, T.; Vanzanella, F.; Peter, H.H. Production of cladospirone bisepoxide, a new fungal metabolite. J. Antibiot. 1994, 47, 1098–1103. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Shan, T.; Li, P.; Huang, Y.; Xu, L.; Zhou, L.; Wang, M.; Jiang, W. Spirobisnaphthalenes from the endophytic fungus Dzf12 of Dioscorea zingiberensis and their antimicrobial activities. Nat. Prod. Commun. 2009, 4, 1469–1472. [Google Scholar] [PubMed]
- Zhou, L.; Yang, C.; Li, J.; Wang, S.; Wu, J. Heptasaccharide and octasaccharide isolated from Paris polyphylla var. yunnanensis and their plant growth-regulatory activity. Plant Sci. 2003, 165, 571–575. [Google Scholar]
- Zhou, L.; Wu, J. Development and application of medicinal plant tissue cultures for production of drugs and herbal medicinals in China. Nat. Prod. Rep. 2006, 23, 789–810. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Cao, X.; Zhang, R.; Peng, Y.; Zhao, S.; Wu, J. Stimulation of saponin production in Panax ginseng hairy roots by two oligosaccharides from Paris polyphylla var. yunnanensis. Biotechnol. Lett. 2007, 29, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, D.K.; Boruah, H.P.D.; Saikia, R.; Bora, T.C. Optimization of process parameters for improved production of bioactive metabolite by a novel endophytic fungus Fusarium sp. DF2 isolated from Taxus wallichiana of North East India. World J. Microbiol. Biotechnol. 2008, 24, 79–87. [Google Scholar] [CrossRef]
- Zhang, R.; Li, P.; Xu, L.; Chen, Y.; Sui, P.; Zhou, L.; Li, J. Enhancement of diosgenin production in Dioscorea zingiberensis cell culture by oligosaccharide elicitor from its endophytic fungus Fusarium oxysporum Dzf17. Nat. Prod. Commun. 2009, 4, 1459–1462. [Google Scholar] [PubMed]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Enhanced beauvericin production with in situ adsorption in mycelial liquid culture of Fusarium redolens Dzf2. Process Biochem. 2009, 44, 1063–1067. [Google Scholar] [CrossRef]
- Putalun, W.; Luealon, W.; De-Eknamkul, W.; Tanaka, H.; Shoyama, Y. Improvement of artemisinin production by chitosan in hairy root cultures of Artemisia annua. Biotechnol. Lett. 2007, 29, 1143–1146. [Google Scholar] [CrossRef] [PubMed]
- Prakash, G.; Srivastava, A.K. Statistical elicitor optimization studies for the enhancement of azadirachtin production in bioreactor Azadirachta indica cell cultivation. Biochem. Eng. J. 2008, 40, 218–226. [Google Scholar] [CrossRef]
- Zhao, J.; Zhou, L.; Wu, J. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol. 2010, 87, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Li, P.; Zhao, J.; Yin, C.; Zhou, L. Endophytic fungi from Dioscorea zingiberensis and their effects on the growth and diosgenin production of the host plant cultures. Nat. Prod. Res. Dev. 2010, 22, 11–15. [Google Scholar]
Sample Availability: The compound diepoxin ζ is available from the authors. |





© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhao, J.; Zheng, B.; Li, Y.; Shan, T.; Mou, Y.; Lu, S.; Li, P.; Zhou, L. Enhancement of Diepoxin ζ Production by Yeast Extract and Its Fractions in Liquid Culture of Berkleasmium-Like Endophytic Fungus Dzf12 from Dioscorea zingiberensis. Molecules 2011, 16, 847-856. https://doi.org/10.3390/molecules16010847
Zhao J, Zheng B, Li Y, Shan T, Mou Y, Lu S, Li P, Zhou L. Enhancement of Diepoxin ζ Production by Yeast Extract and Its Fractions in Liquid Culture of Berkleasmium-Like Endophytic Fungus Dzf12 from Dioscorea zingiberensis. Molecules. 2011; 16(1):847-856. https://doi.org/10.3390/molecules16010847
Chicago/Turabian StyleZhao, Jianglin, Bingbing Zheng, Yan Li, Tijiang Shan, Yan Mou, Shqiong Lu, Peiqin Li, and Ligang Zhou. 2011. "Enhancement of Diepoxin ζ Production by Yeast Extract and Its Fractions in Liquid Culture of Berkleasmium-Like Endophytic Fungus Dzf12 from Dioscorea zingiberensis" Molecules 16, no. 1: 847-856. https://doi.org/10.3390/molecules16010847