Molecular Characterization and Tandem Mass Spectrometry of the Lectin Extracted from the Seeds of Dioclea sclerocarpa Ducke

Abstract
:1. Introduction
2. Results and Discussion





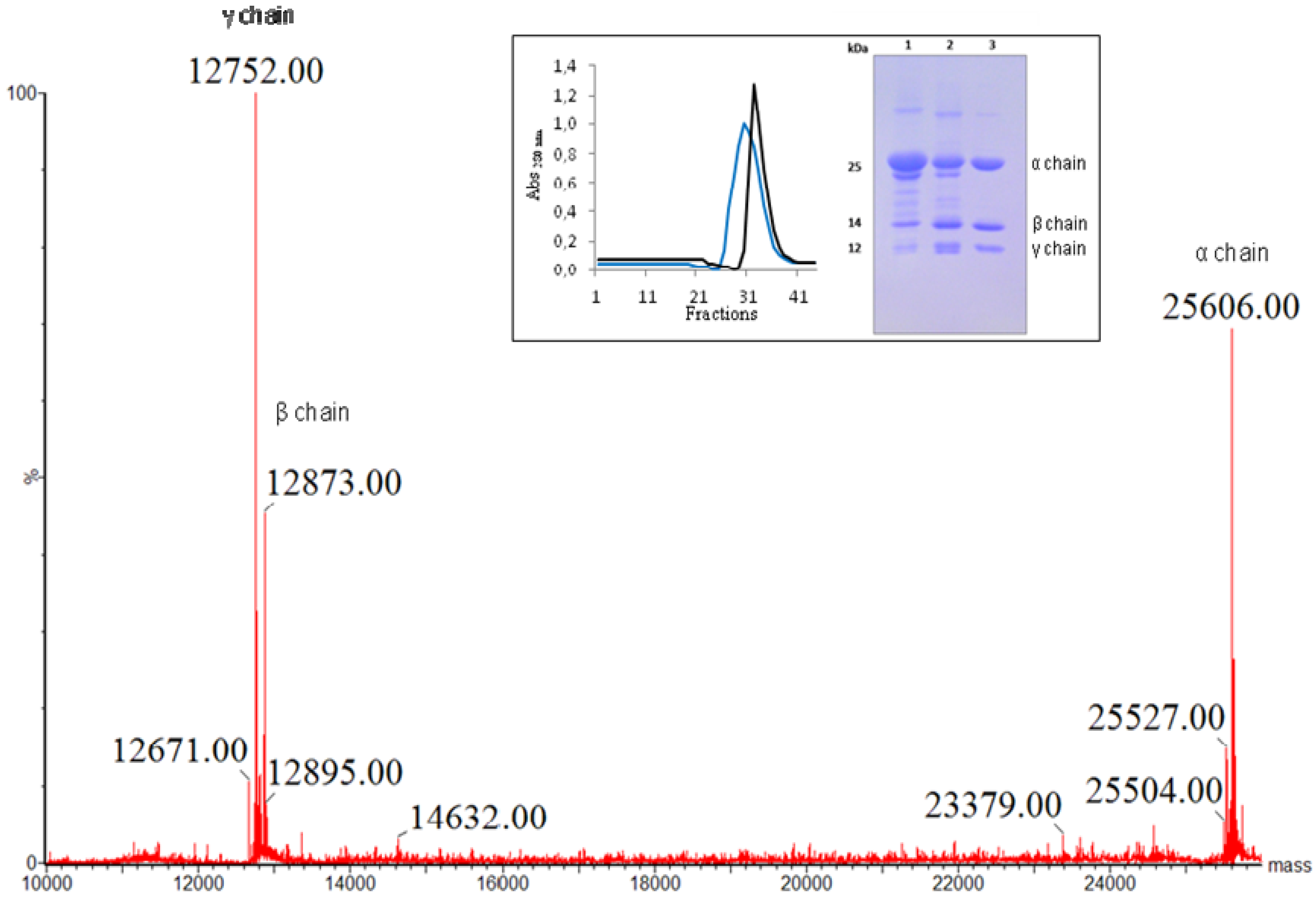{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Experimental mass (Da) | Sequence |
|---|---|---|
| T1 | 3254.5981 | ADTIVAVELDSYPNTDIGDPSYPHIGIDIK |
| T2 | 1004.5582 | SIRSKSTAR |
| T3 * | 863.3566 | WNMQTGK |
| T4 | 1373.7257 | VGTVHISYNSVAK |
| T5 | 3243.5935 | LSAVVSYSGSSSTTVSYDVDLNNVLPEWVR |
| T6 | 1108.5526 | VGLSATTGLYK |
| T7 | 1512.6473 | ETNTILSWSFTSK |
| T8 | 1845.6696 | TNSIADENSLHFSFHK |
| T9 | 4044.8477 | FSQNPKDLILQGDAFTDSDGNLQLTK |
| T10 | 1345.6222 | VSSSGDPQGNSVGR |
| T11 | 1472.7642 | ALFYAPVHIWEK |
| T12 * | 1715.8588 | SAVVASFDATFTFLIK |
| T13 | 2864.1970 | SPDREPADGITFFIANTDTSIPSGSGGR |
| T14 | 958.4971 | LLGLFPDAN |
| Q1 | 2342.9988 | DSYPNTDIGDPNYPHIGIDIK |
| Q2 | 1533.7388 | NMQTGKVGTVHISY |
| Q3 | 1392.7252 | NSVAKRLSAVVSY |
| Q4 | 974.4125 | SGSSSTTVSY |
| Q5 | 1312.6182 | DVDLNNVLPEW |
| Q6 | 1090.5066 | KETNTILSW |
| Q7 | 2137.9294 | SFTSKLKTNSIADENSLHF |
| Q8 | 1446.7222 | SFHKFSQNPKDL |
| Q9 | 2246.9380 | SQNPKDLILQGDAFTDSDGNL |
| Q10 | 1999.9325 | QLTKVSSSGDPQGNSVGRALF |
| Q11 | 884.4222 | YAPVHIW |
| Q12 | 1370.6240 | KSAVVASFDATF |
| Q13 | 1544.6987 | IKSPDREPADGITF |
| Q14 | 1804.9670 | FIANTDTSIPSGSGGRLL |

3. Experimental
3.1. Materials
3.2. Protein Extraction Procedure
3.3. Hemagglutinating Activity
3.4. Sugar Specificity
3.5. Protein Purification
3.6. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis
3.7. Lectin Metal Dependence
3.8. Carbohydrate Content Analysis
3.9. Circular Dichroism and Thermostability
3.10. MW Determination by Mass Spectrometry
3.11. Protein Digestion and Tandem Mass Spectrometry Analysis
3.12. Primary Structure Analysis
4. Conclusions
Acknowledgments
References and Notes
- van Damme, E.J.M.; Peumans, W.J.; Barre, A.; Rougé, P. Plant lectins: A composite of several distinct families of structurally and evolutionary related proteins with diverse biological roles. CRCCrit. Rev. Plant Sci. 1998, 17, 575–692. [Google Scholar]
- Kocourek, J.; Horejsi, V. A note of the recent discussion on definition of the term “lectin”. In Lectins: Biology, Biochemistry, Clinical Biochemistry; Bog-Hansen, T.C., Spengler, G.A., Eds.; Walter de Gruyter: Berlin, Germany, 1983; Volume 3, pp. 3–6. [Google Scholar]
- Moreira, R.A.; Ainouz, I.L.; Oliveita, J.T.A.; Cavada, B.S. Plant lectins: Chemical and biological aspects. Mem. Inst. Oswaldo Cruz 1991, 86, 212–218. [Google Scholar]
- Peumans, W.; Barre, A.; Hao, Q.; Rougé, P.; van Damme, E.J.M. Higher plants developed structurally different motifs to recognize foreign glycans. Trends Glycosci. Glycotechnol. 2000, 12, 83–101. [Google Scholar] [CrossRef]
- Sharon, N.; Lis, H. History of lectins: from hemagglutinins to biological recognition molecules. Glycobiology 2004, 14, 53–62. [Google Scholar] [CrossRef]
- Sharon, N. Lectins: Carbohydrate-specific reagents and biological recognition molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef]
- Loris, R.; Halmeryck, T.; Bouckaert, J.; Wyns, L. Legume Lectin Structure. Biochim. Biophys. Acta 1998, 1383, 9–36. [Google Scholar]
- Ferreira, R.R.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Gomes, J.C. Characteristics of the histamine release from hamster cheek pouch mast cells stimulated by lectins from Brazilian beans And Concanavalin A. Inflamm. Res. 1996, 45, 442–447. [Google Scholar] [CrossRef]
- Asseury, A.M.S.; Shimbuya, M.D.; Martins, G.J.; Souza, M.L.P.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Ribeiro, R.A.; Flores, C.A. Antiinflammatory effect of glucose-mannose binding lectins isolated from brazilian beans. Mediators Inflamm. 1997, 6, 201–210. [Google Scholar] [CrossRef]
- Alencar, N.M.N.; Teixeira, E.H.; Asseury, A.M.; Cavada, B.S.; Flores, C.A.; Ribreiro, R.A. Leguminous lectins as tools for studying the role of sugar residues in leukocyte recruitment. Mediators Inflamm. 1998, 8, 107–113. [Google Scholar]
- Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Crisostomo, C.V.; Silva, L.M.; Moreira, R.A.; Oliveira, J.T.A. Lectin from Dioclea guianensis var. lasiophylla Duke seeds mobilization during germination and seedlings growth in the dark. Rev. Bras. Fisiol. Veg. 1994, 6, 21–25. [Google Scholar]
- Sanz-Aparicio, J.; Hermoso, J.; Grangeiro, T.B.; Calvette, J.J.; Cavada, B.S. The crystal structure of Canavalia brasiliensis lectin suggests a correlation between its quaternary conformation and its distinct biological properties from Concanavalin A. FEBS Lett. 1997, 405, 114–118. [Google Scholar] [CrossRef]
- Moreira, R.A.; Monteiro, A.C.O.; Horta, A.C.G.; Oliveira, J.T.A.; Cavada, B.S. Isolation and characterization of Dioclea altissima Var. Megacarpa seed lectin. Phytochemstry 1997, 46, 139–144. [Google Scholar]
- Cavada, B.S.; Grangeiro, T.B.; Ramos, M.V.; Cordeiro, E.F.; Oliveira, J.T.A.; Moreira, R.A. Isolation and partial characterization of a lectin from Dioclea rostrata Benth seeds. Rev. Bras. Fisiol. Veg. 1996, 8, 31–36. [Google Scholar]
- Moreira, R.A.; Cordeiro, E.F.; Ramos, M.V.; Grangeiro, T.B.; Martins, J.L.; Oliveira, J.T.A.; Cavada, B.S. Isolation and partial characterization of a lectin from seeds of Dioclea violacea. Rev. Bras. Fisiol. Veg. 1996, 8, 23–29. [Google Scholar]
- Vasconcellos, I.M.; Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A. Purification and partial characterization of a lectin from the seed of Dioclea guianensis. J. Food Biochem. 1991, 15, 137–154. [Google Scholar] [CrossRef]
- Moreira, R.A.; Barros, A.C.H.; Stewart, J.C.; Pusztai, A. Isolation and characterization of a lectin from the seeds of Dioclea grandiflora (Mart.). Planta 1983, 158, 63–69. [Google Scholar]
- Cavada, B.S.; Moreira, R.A.; Oliveira, J.T.A.; Grangeiro, T.B. Primary structures and functions of plant lectins. Rev. Bras. Fisiol. Veg. 1993, 5, 193–201. [Google Scholar]
- Moreira, R.A.; Ainouz, I.L.; de Oliveira, J.T.A.; Cavada, B.S. Plant lectins, chemical and biological aspects. Mem. Inst. Oswaldo. Cruz 1991, 86 suppl. П, 211–218. [Google Scholar] [CrossRef]
- Gouet, P.; Robert, X.; Courcelle, E. ESPript/ENDscript: Extracting and rendering sequence and 3D information from atomic structure of proteins. Nucleic Acids Res. 2003, 31, 3320–3323. [Google Scholar] [CrossRef]
- Naismith, J.H.; Emmerich, C.; Habash, J.; Harrop, S.J.; Helliwell, J.R.; Hunter, W.N.; Raftery, J.; Kalb, A.J.; Yariv, J. Refined structure of concanavalin A complexed with methyl alpha-D-mannopyranoside at 2.0 Å resolution and comparison with the saccharide-free structure. Acta Crystallogr. D Biol. Crystallogr. 1994, 50, 847–858. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Biochemistry 1976, 72, 248–254. [Google Scholar]
- Moreira, R.A.; Perrone, J.C. Purification and partial characterization of a lectin from Phaseolus vulgaris. Plant Physiol. 1977, 59, 783–787. [Google Scholar] [CrossRef]
- Grant, G.; McKenzie, N.H.; Moreira, R.A.; Pusztai, A. Dioclea grandiflora and Dioclea sclerocarpa seeds. A nutritional study. Plant Foods Hum. Nutr. 1986, 36, 47–61. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Ferrige, A.G.; Seddon, M.J.; Green, B.N.; Jarvis, S.A.; Skilling, J. Disentangling electrospray spectra with maximum entropy. Rapid Commun. Mass Spectrom. 1992, 6, 707–711. [Google Scholar]
- Altschul, S.F.; Maddem, T.L.; Scharffer, A.A.; Zhang, L.; Zhang, Z.; Miller, W.; Lipman, D.L. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Corpet, F. Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res. 1988, 16, 10881–10890. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar]
- Sample Availability: Contact the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Correia, J.L.A.; Nascimento, A.S.F.d.; Cajazeiras, J.B.; Gondim, A.C.S.; Pereira, R.I.; Sousa, B.L.d.; Silva, A.L.C.d.; Garcia, W.; Teixeira, E.H.; Nascimento, K.S.d.; et al. Molecular Characterization and Tandem Mass Spectrometry of the Lectin Extracted from the Seeds of Dioclea sclerocarpa Ducke. Molecules 2011, 16, 9077-9089. https://doi.org/10.3390/molecules16119077
Correia JLA, Nascimento ASFd, Cajazeiras JB, Gondim ACS, Pereira RI, Sousa BLd, Silva ALCd, Garcia W, Teixeira EH, Nascimento KSd, et al. Molecular Characterization and Tandem Mass Spectrometry of the Lectin Extracted from the Seeds of Dioclea sclerocarpa Ducke. Molecules. 2011; 16(11):9077-9089. https://doi.org/10.3390/molecules16119077
Chicago/Turabian StyleCorreia, Jorge Luis Almeida, Antônia Sâmia Fernandes do Nascimento, João Batista Cajazeiras, Ana Cláudia Silva Gondim, Ronniery Ilario Pereira, Bruno Lopes de Sousa, André Luiz Coelho da Silva, Wanius Garcia, Edson Holanda Teixeira, Kyria Santiago do Nascimento, and et al. 2011. "Molecular Characterization and Tandem Mass Spectrometry of the Lectin Extracted from the Seeds of Dioclea sclerocarpa Ducke" Molecules 16, no. 11: 9077-9089. https://doi.org/10.3390/molecules16119077