Phytoceramide Shows Neuroprotection and Ameliorates Scopolamine-Induced Memory Impairment

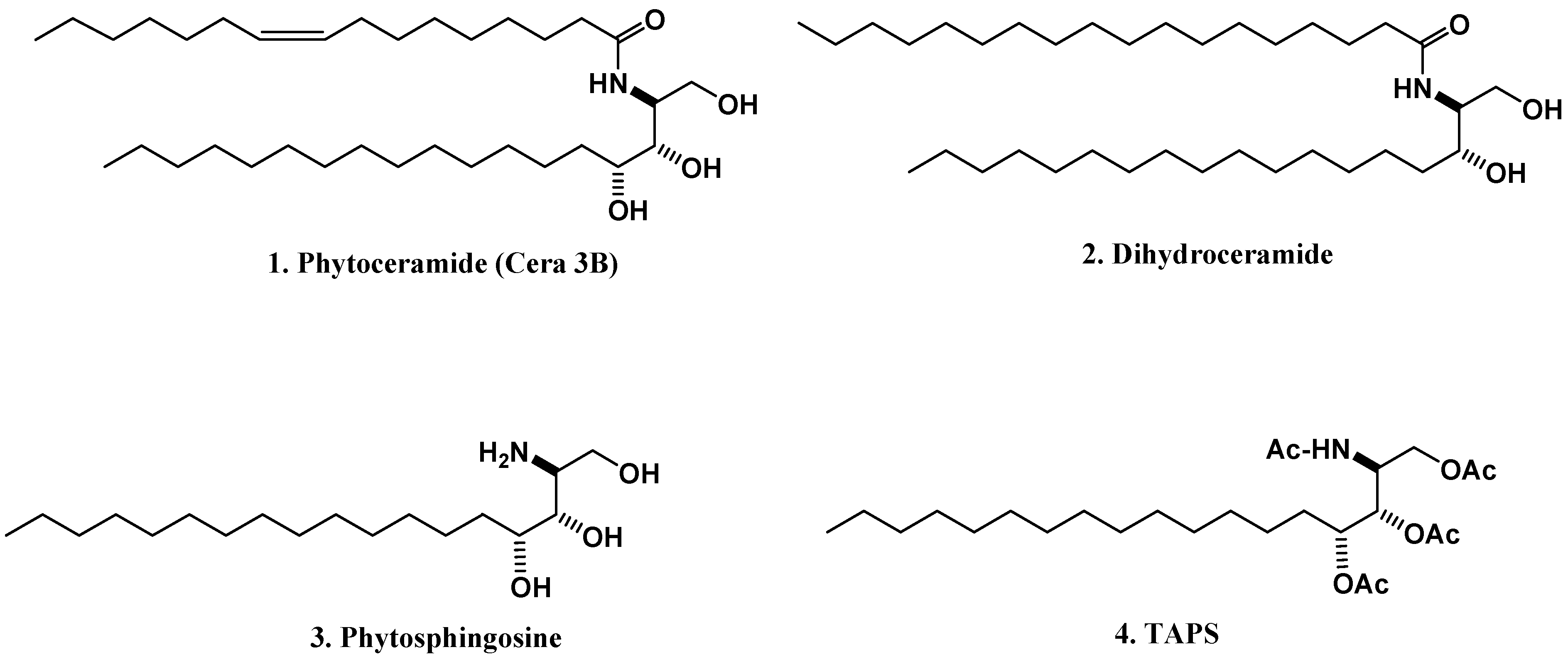{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. Results and Discussion
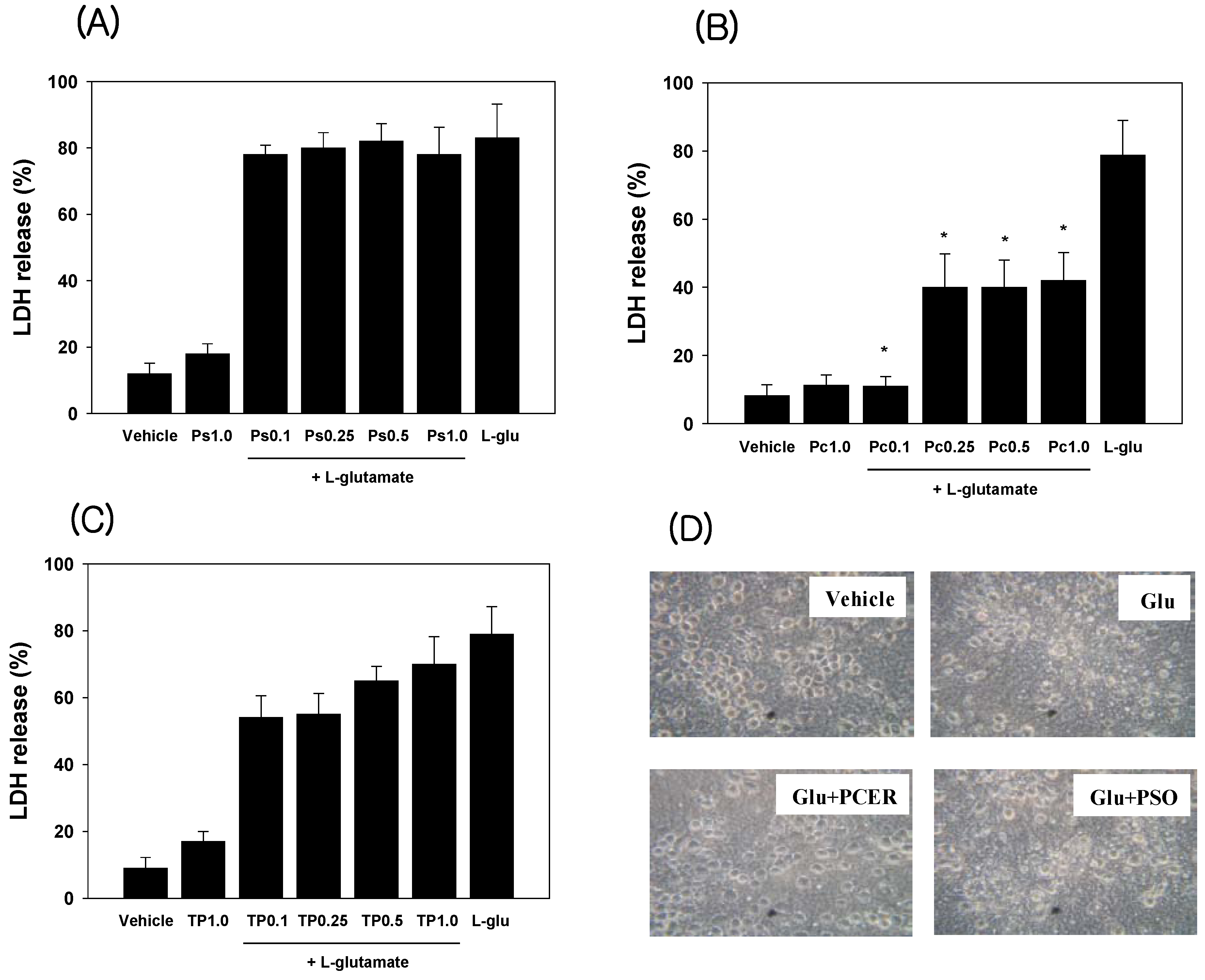
2.1. Neuroprotection in Cultured Neuronal Cells

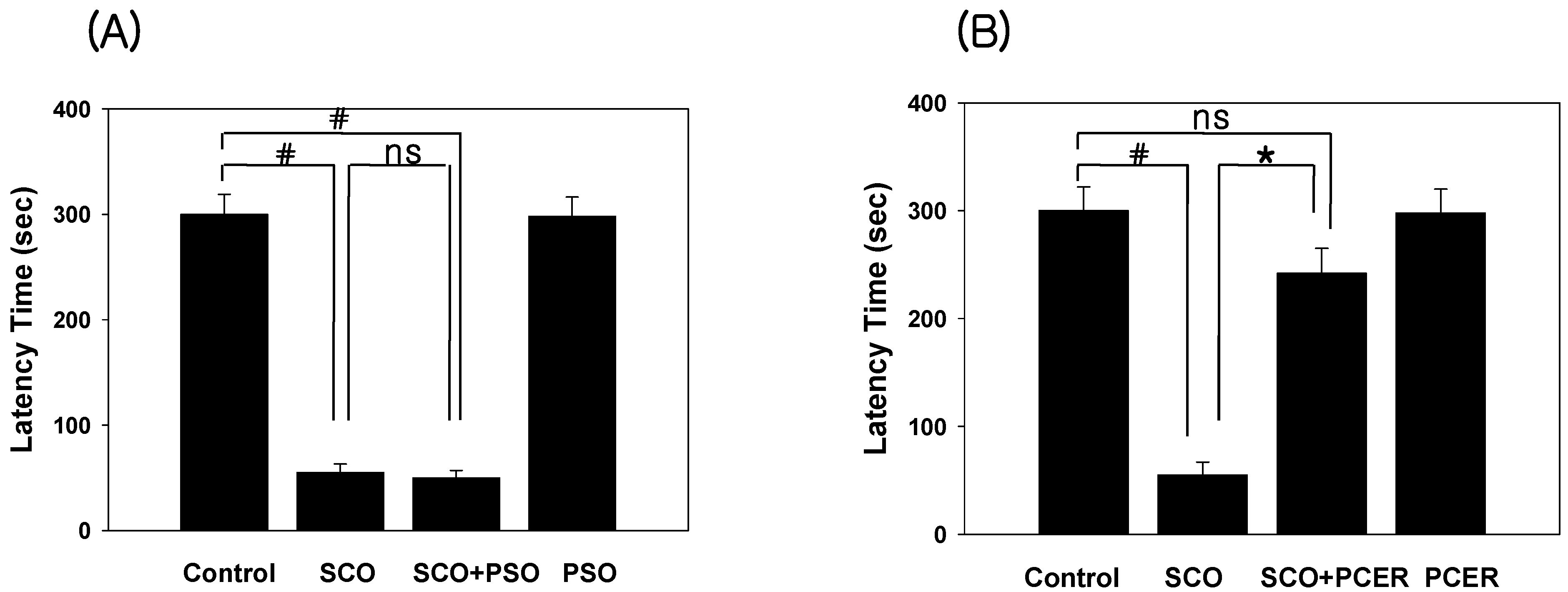
2.2. Amelioration of the Scopolamine-Induced Memory Deficits in the Passive Avoidance Test

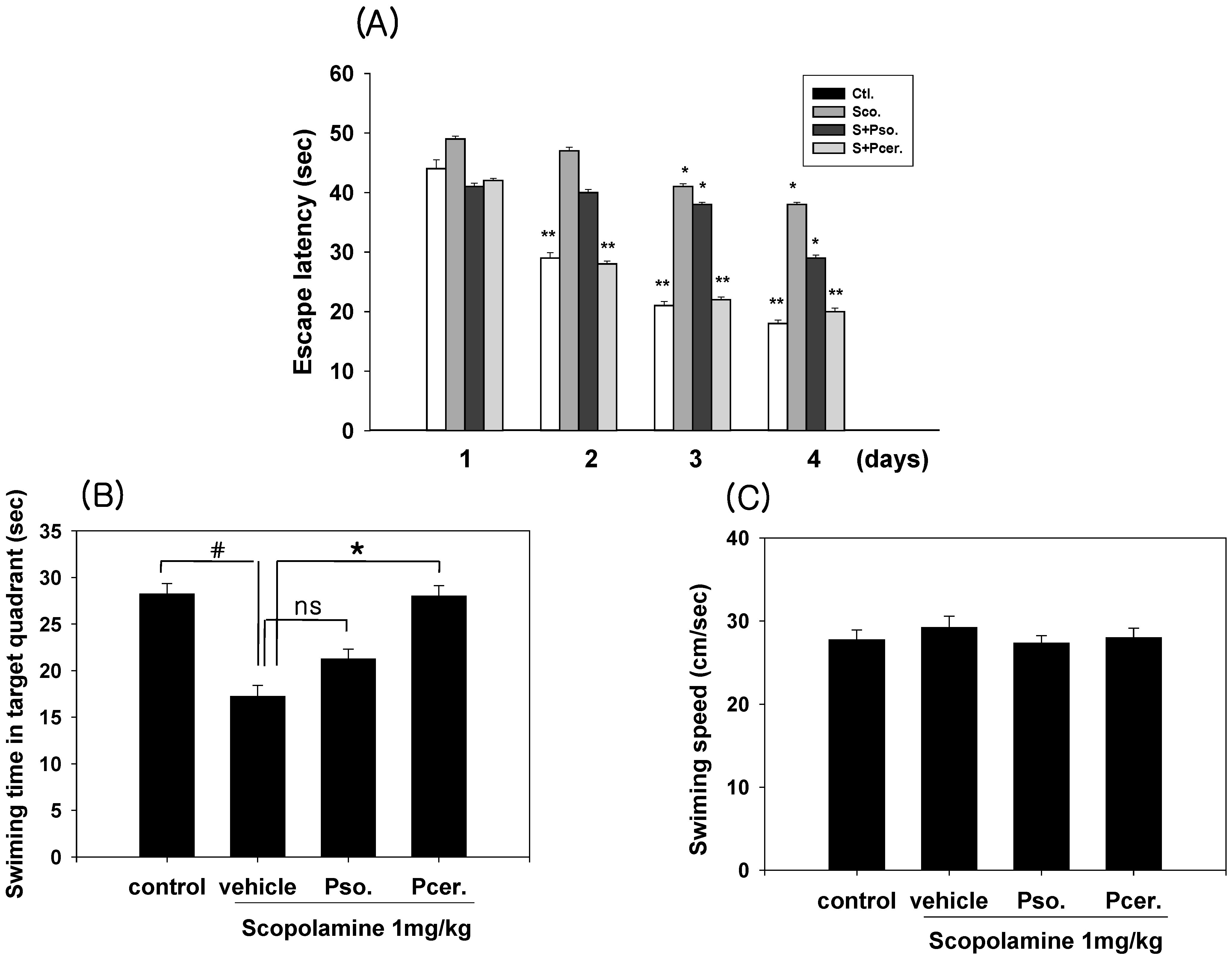
2.3. Amelioration of the Scopolamine-Induced Memory Deficits in Morris Water Maze Test

3. Experimental
3.1. Materials and Methods
3.2. Cortical Neuronal Cell Culture
3.3. Lactate Dehydrogenase (LDH) Assay
3.4. Passive Avoidance Test
3.5. Morris Water Maze Test
3.6. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Mergenthaler, P.; Dirnagl, U.; Meisel, A. Pathophysiology of stroke: Lessons from animal models. Metab. Brain Dis. 2004, 19, 151–167. [Google Scholar] [CrossRef]
- Richard, M.J.P.; Connell, B.J.; Khan, B.V.; Saleh, T.M. Cellular mechanisms by which lipoic acid confers protection during the early stages of cerebral ischemia: A possible role for calcium. Neurosci. Res. 2011, 69, 299–307. [Google Scholar] [CrossRef]
- Bartus, R.T.; Dean, R.L.; Beer, B.; Lippa, A.S. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar]
- Dolezal, V.; Tucek, S. Positive and negative effects of tacrin and methoxytacrine on the metabolism of acetylcholine in brain cortical prisms incubated under “resting” conditions. J. Neurochem. 1991, 56, 1207–1215. [Google Scholar] [CrossRef]
- Ennaceur, A.; Meliani, K. Effects of physostigmine and scopolamine on rats’ performances in object-recognition and radial-maze tests. Psychopharmacology 1992, 109, 321–330. [Google Scholar] [CrossRef]
- Elena, I.; de Chaves, P. Sphingolipids in apoptosis, survival and regeneration in the nervous system. Biochim. Biophys. Acta 2006, 1758, 1995–2015. [Google Scholar]
- Spiegel, S.; Milstien, S. Sphingosine-1-phosphate: An enigmatic signalling lipid. Nat. Rev. Mol. Cell Biol. 2003, 4, 397–407. [Google Scholar] [CrossRef]
- Anne, G.B.; Dimitri, P.; Leyre, B.; Virginie, G.; Marie-Franc, O.A.; Marie-Lise, M.; Marie-Bernadette, D.; Olivier, C. Critical role for sphingosine kinase-1 in regulating survival of neuroblastoma cells exposed to amyloid-β peptide. Mol. Pharmacol. 2007, 72, 341–349. [Google Scholar] [CrossRef]
- Garcia, J.; Shea, J.; Alvarez-Vasquez, F.; Quershi, A.; Luberto, C.; Voit, E.O.; Del Poeta, M. Mathmatical modeling of pathogenicity of Cryptococcus neoformans. Mol. Syst. Biol. 2008, 4, 1–13. [Google Scholar]
- Dickson, R.C.; Lester, R.L. Yeast sphingolipids. Biochim. Biophys. Acta 1999, 1426, 347–357. [Google Scholar]
- Mao, C.; Xu, R.; Szulc, Z.M.; Bielawska, A.; Galadari, S.H.; Obeid, L.M. Cloning and characterization of a novel human alkaline ceramidase: A mammalian enzyme that hydrolyzes phytoceramide. J. Biol. Chem. 2001, 276, 26577–26588. [Google Scholar]
- Koh, J.Y.; Choi, D.W. Quantitative determination of glutamate mediated cortical neuronal injury in cell culture by lactate dehydrogenase efflux assay. J. Neurosci. Methods 1987, 20, 83–90. [Google Scholar] [CrossRef]
- Rubaj, A.; Zgodzinski, W.; Sieklucka-Dziuba, M. The influence of adenosine A3 receptor agonist: IB-MECA, on scopolamine- and MK-801-induced memory impairment. Behav. Brain Res. 2003, 141, 11–17. [Google Scholar] [CrossRef]
- Hannun, Y.A. The sphingomyelin cycle and second messenger function of ceramide. J. Biol. Chem. 1994, 269, 3125–3128. [Google Scholar]
- Tang, N.; Ong, W.Y.; Zhang, E.M.; Chen, P.; Yeo, J.F. Differential effects of ceramide species on exocytosis in rat PC12 cells. Exp. Brain Res. 2007, 183, 241–247. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Paola, R.D.; Genovese, T.; Mazzon, E.; Esposite, E.; Crisafulli, C.; Bramanti, P.; Salvemini, D. Anti-inflammatory and anti-apoptotic effects of fumonisin B1, an inhibitor of ceramide synthase, in a rodent model of splanchnic ischemia and reperfusion injury. J. Pharmacol. Exp. Ther. 2008, 327, 45–57. [Google Scholar] [CrossRef]
- Gangoiti, P.; Granado, M.H.; Wang, S.W.; Kong, J.Y.; Steinbrecher, U.P.; Gomez-Munoz, A. Ceramide 1-phosphate stimulates macrophage proliferation through activation of the PI3-kinase/PKB, JNK and ERK1/2 pathways. Cell Signal. 2008, 20, 726–736. [Google Scholar] [CrossRef]
- Mitoma, J.; Ito, M.; Furaya, S.; Hirabayashi, Y. Bipotential roles of ceramide in the growth of hippocampal neurons: Promotion of cell survival and dendritic outgrowth in dose- and developmental stage-dependent manners. J. Neurosci. Res. 1998, 51, 712–722. [Google Scholar] [CrossRef]
- Irie, F.; Hirabayashi, Y. Ceramide prevents motoneuronal cell death through inhibition of oxidative signal. Neurosci. Res. 1999, 35, 135–144. [Google Scholar] [CrossRef]
- LeDoux, J.E. Emotional memory systems in the brain. Behav. Brain Res. 1993, 58, 69–79. [Google Scholar] [CrossRef]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]
- Blokland, A.; Geraerts, E.; Been, M. A detailed analysis of rats’ spatial memory in a probe trial of a Morris task. Behav. Brain Res. 2004, 154, 71–75. [Google Scholar] [CrossRef]
- Bae, J.H.; Sohn, J.H.; Park, C.S.; Rhee, J.S.; Choi, E.S. Integrative transformation system for the metabolic engineering of the sphingoid base-producing yeast Pichia ciferrii. Appl. Environ. Microbiol. 2003, 69, 812–819. [Google Scholar] [CrossRef]
- Park, C.H.; Choi, S.H.; Koo, J.W.; Seo, J.H.; Kim, H.S.; Jeong, S.J.; Suh, Y.H. Novel cognitive improving and neuroprotective activities of Polygala Tenuifolia Willdenow extract, BT-11. J. Neurosci. Res. 2002, 70, 484–492. [Google Scholar] [CrossRef]
- Sample Availability: Contact to the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jung, J.-C.; Lee, Y.; Moon, S.; Ryu, J.H.; Oh, S. Phytoceramide Shows Neuroprotection and Ameliorates Scopolamine-Induced Memory Impairment. Molecules 2011, 16, 9090-9100. https://doi.org/10.3390/molecules16119090
Jung J-C, Lee Y, Moon S, Ryu JH, Oh S. Phytoceramide Shows Neuroprotection and Ameliorates Scopolamine-Induced Memory Impairment. Molecules. 2011; 16(11):9090-9100. https://doi.org/10.3390/molecules16119090
Chicago/Turabian StyleJung, Jae-Chul, Yeonju Lee, Sohyeon Moon, Jong Hoon Ryu, and Seikwan Oh. 2011. "Phytoceramide Shows Neuroprotection and Ameliorates Scopolamine-Induced Memory Impairment" Molecules 16, no. 11: 9090-9100. https://doi.org/10.3390/molecules16119090