The Dose-Effect of Icariin on the Proliferation and Osteogenic Differentiation of Human Bone Mesenchymal Stem Cells

Abstract
:1. Introduction
2. Results and Discussion
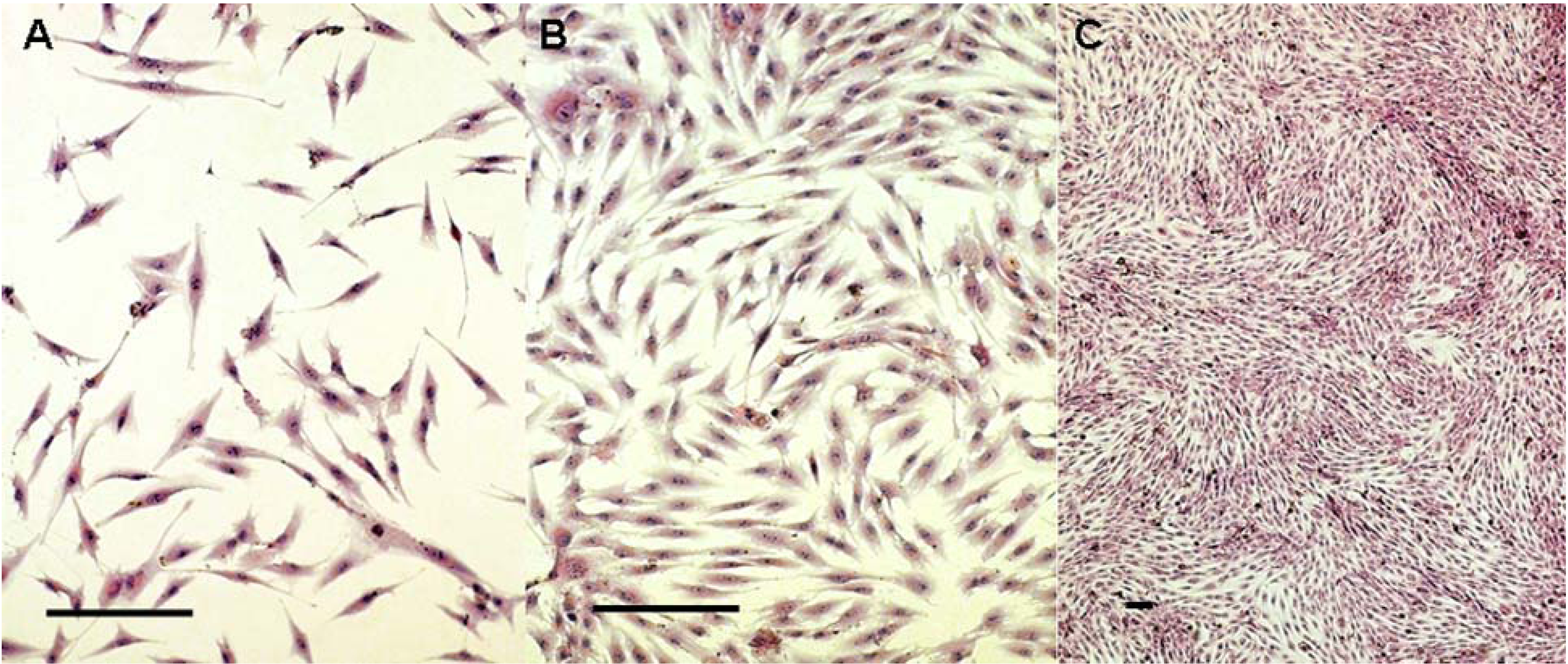
2.1. Cells Observation
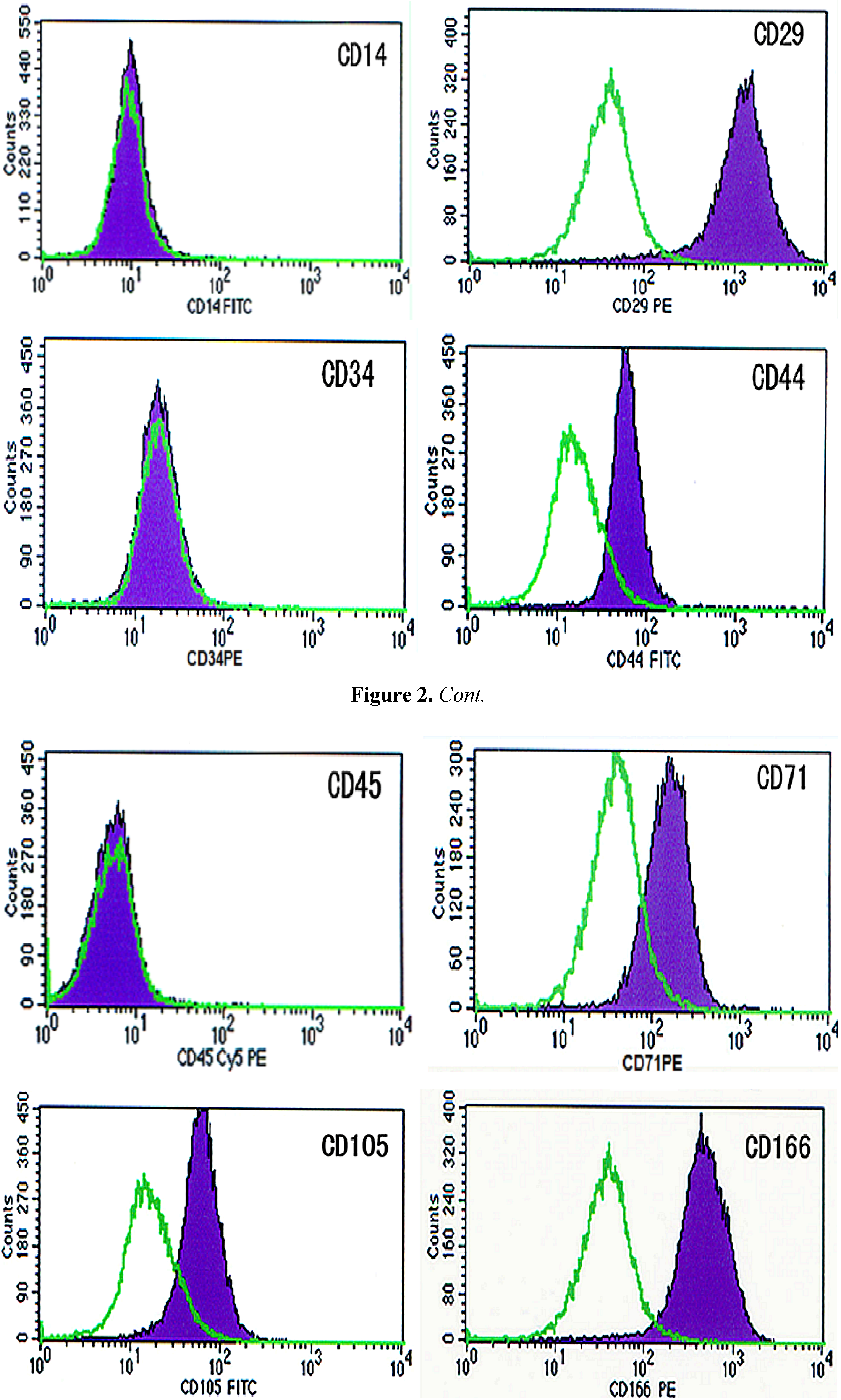
2.2. Identification of BMSCs


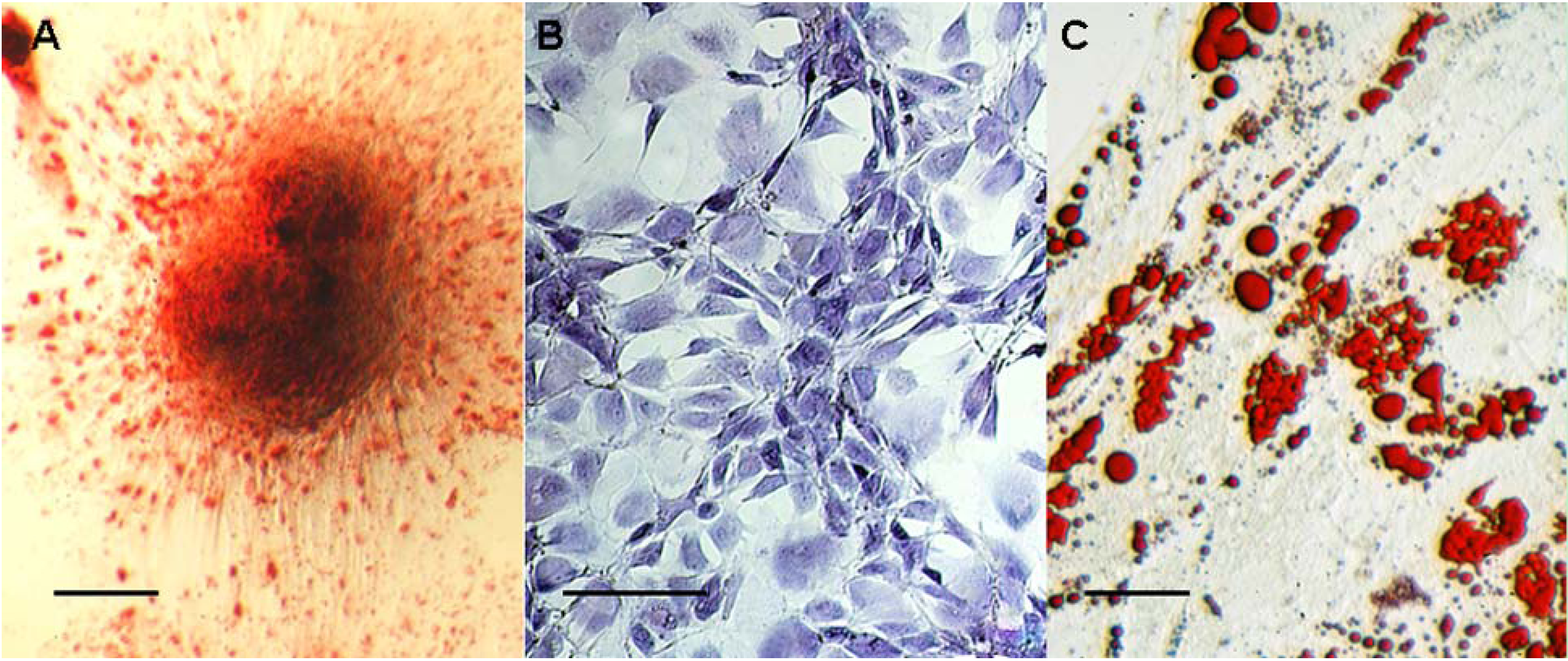
2.3. hBMSC Differentiation Assay

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
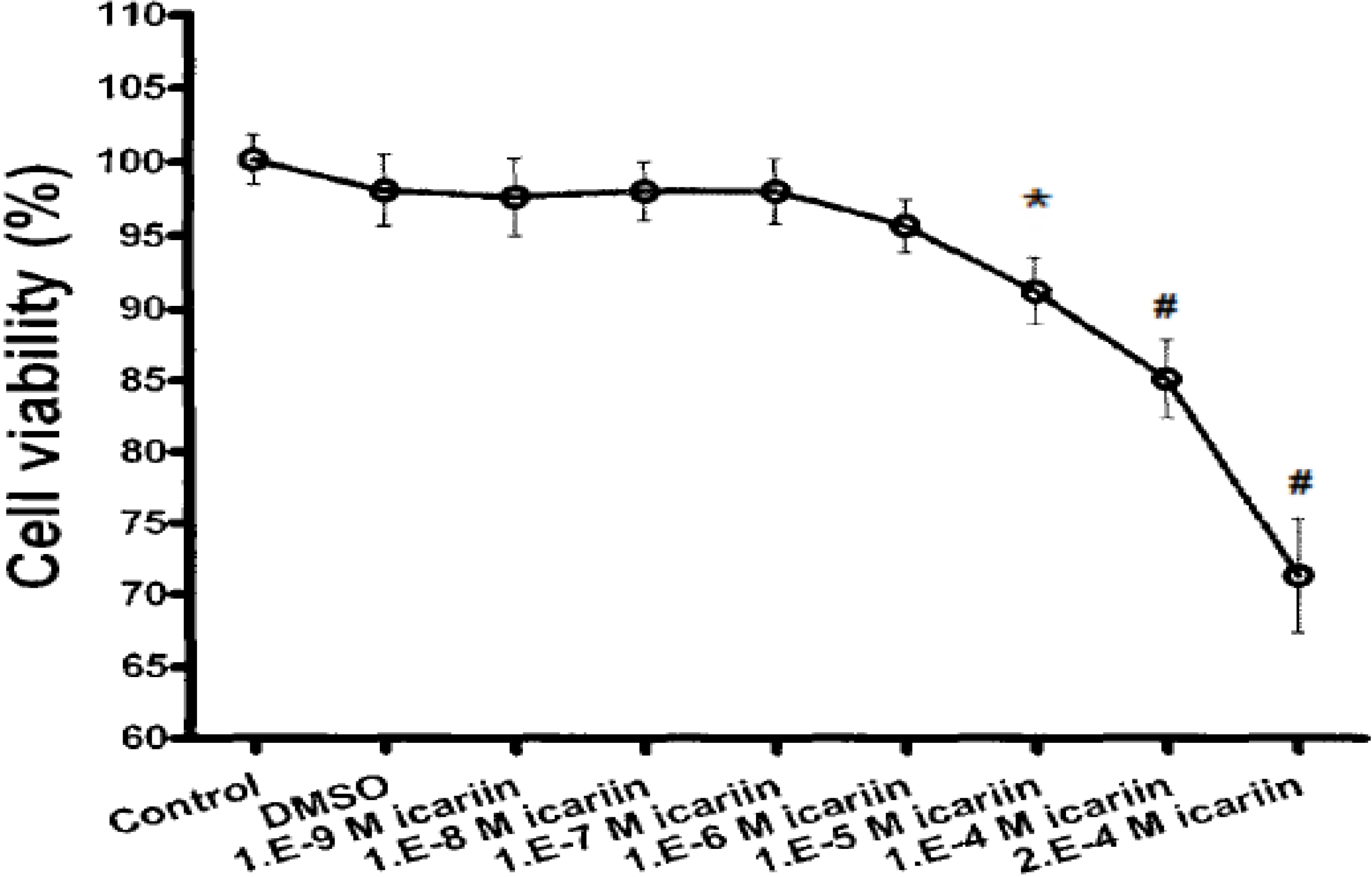
2.4. Cytotoxicity of Icariin


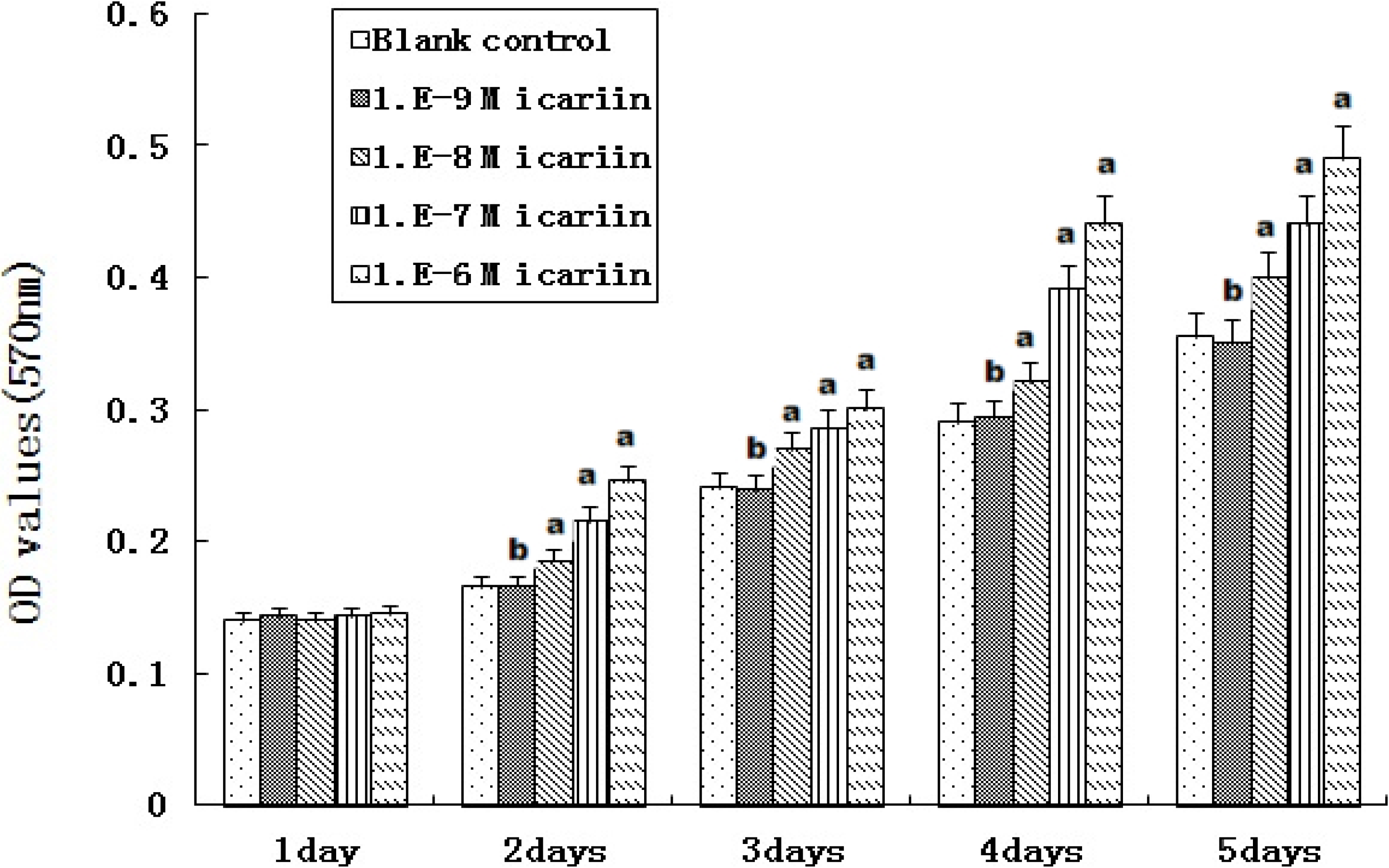
2.5. Proliferation of hBMSCs Stimulated by Icariin
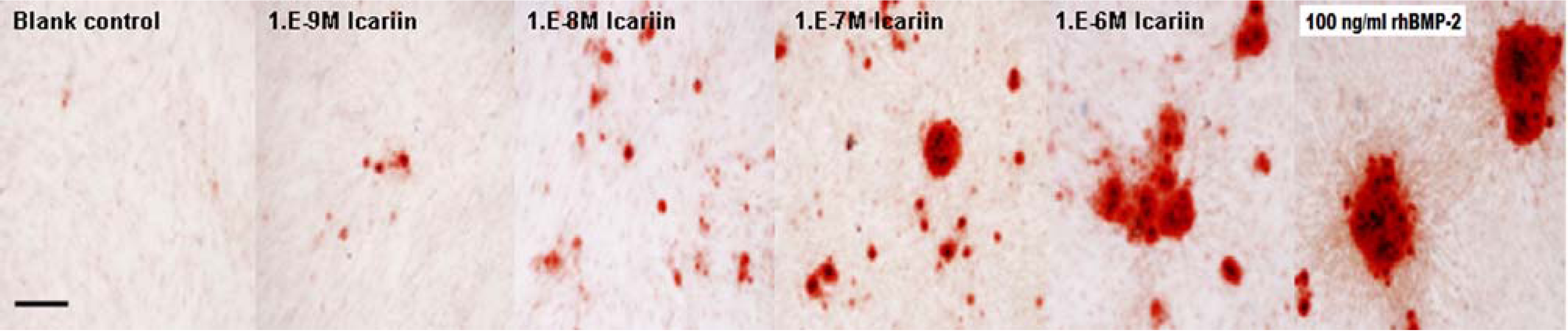
2.6. Osteogenic Differentiation of hBMSCs Stimulated by Icariin


3. Experimental
3.1. Isolation and Expansion of hBMSCs
3.2. Phenotype Identification of Cells
3.3. Inducing Differentiation of Cells
3.4. Cytotoxicity Test of Icariin
3.5. Proliferation of hBMSCs Stimulated by Icariin
3.6. Osteogenic Differentiation of hBMSCs Stimulated by Icariin
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References and Notes
- Kretlow, J.D.; Mikos, A.G. Review: Mineralization of synthetic polymer scaffolds for bone tissue engineering. Tissue Eng. 2007, 13, 927–938. [Google Scholar] [CrossRef]
- Hofmann, A.; Ritz, U.; Verrier, S.; Eglin, D.; Alini, M.; Fuchs, S.; Kirkpatrick, C.J.; Rommens, P.M. The effect of human osteoblasts on proliferation and neo-vessel formation of human umbilical vein endothelial cells in a long-term 3D co-culture on polyurethane scaffolds. Biomaterials 2008, 29, 4217–4226. [Google Scholar] [CrossRef]
- Caterson, E.J.; Nesti, L.J.; Danielson, K.G.; Tuan, R.S. Human marrow-derived mesenchymal progenitor cells: Isolation, culture expansion, and analysis of differentiation. Mol. Biotechnol. 2002, 20, 245–256. [Google Scholar] [CrossRef]
- Lee, J.Y.; Seol, Y.J.; Kim, K.H.; Lee, Y.M.; Park, Y.J.; Rhyu, I.C.; Chung, C.P.; Lee, S.J. Transforming growth factor (TGF)-beta1 releasing tricalcium phosphate/chitosan microgranules as bone substitutes. Pharm. Res. 2004, 21, 1790–1796. [Google Scholar] [CrossRef]
- Bi, L.; Cheng, W.; Fan, H.; Pei, G. Reconstruction of goat tibial defects using an injectable tricalcium phosphate/chitosan in combination with autologous platelet-rich plasma. Biomaterials 2010, 31, 3201–3211. [Google Scholar] [CrossRef]
- Okada, M.; Sangadala, S.; Liu, Y.; Yoshida, M.; Reddy, B.V.; Titus, L.; Boden, S.D. Development and optimization of a cell-based assay for the selection of synthetic compounds that potentiate bone morphogenetic protein-2 activity. Cell Biochem. Funct. 2009, 27, 526–534. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Z.G.; Li, C.; Hu, S.J.; Liu, L.; Wang, J.P.; Mei, Q.B. Du-Zhong (Eucommia ulmoides Oliv.) cortex extract prevent OVX-induced osteoporosis in rats. Bone 2009, 45, 553–559. [Google Scholar] [CrossRef]
- Zhao, J.; Ohba, S.; Komiyama, Y.; Shinkai, M.; Chung, U.I.; Nagamune, T. Icariin: A potential osteoinductive compound for bone tissue engineering. Tissue Eng. Part A 2010, 16, 233–243. [Google Scholar] [CrossRef]
- Jiang, F.; Wang, X.L.; Wang, N.L.; Yao, X.S. Two new flavonol glycosides from Epimedium koreanum Nakai. J. Asian Nat. Prod. Res. 2009, 11, 401–409. [Google Scholar] [CrossRef]
- Hsieh, T.P.; Sheu, S.Y.; Sun, J.S.; Chen, M.J.; Liu, M.H. Icariin isolated from Epimedium pubescens regulates osteoblasts anabolism through BMP-2, SMAD4, and Cbfa1 expression. Phytomedicine 2010, 17, 414–423. [Google Scholar] [CrossRef]
- Yin, X.X.; Chen, Z.Q.; Liu, Z.J.; Ma, Q.J.; Dang, G.T. Icariine stimulates proliferation and differentiation of human osteoblasts by increasing production of bone morphogenetic protein 2. Zhonghua Yi Xue Za Zhi (Taipei) 2007, 120, 204–210. [Google Scholar]
- He, W.; Li, Z.L.; Cui, Y.L.; Yi, B.; Liang, C.; Wang, X.X.; Li, Y.; Wang, X. Effect of icariin on the mRNA expressions of Cbfalpha1, BMP2, BMP4 in rat osteoblasts. Beijing Da Xue Xue Bao 2009, 41, 669–673. [Google Scholar]
- Mareddy, S.; Crawford, R.; Brooke, G.; Xiao, Y. Clonal isolation and characterization of bone marrow stromal cells from patients with osteoarthritis. Tissue Eng. 2007, 13, 819–829. [Google Scholar] [CrossRef]
- Zhao, J.; Ohba, S.; Shinkai, M.; Chung, U.I.; Nagamune, T. Icariin induces osteogenic differentiation in vitro in a BMP- and Runx2-dependent manner. Biochem. Biophys. Res. Commun. 2008, 369, 444–448. [Google Scholar] [CrossRef]
- Nian, H.; Ma, M.H.; Nian, S.S.; Xu, L.L. Antiosteoporotic activity of icariin in ovariectomized rats. Phytomedicine 2009, 16, 320–326. [Google Scholar] [CrossRef]
- Chen, K.M.; Ge, B.F.; Ma, H.P.; Liu, X.Y.; Bai, M.H.; Wang, Y. Icariin, a flavonoid from the herb Epimedium enhances the osteogenic differentiation of rat primary bone marrow stromal cells. Pharmazie 2005, 60, 939–942. [Google Scholar]
- Xiao, Q.; Chen, A.; Guo, F. Effects of Icariin on expression of OPN mRNA and type I collagen in rat osteoblasts in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2005, 25, 690–692. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fan, J.-J.; Cao, L.-G.; Wu, T.; Wang, D.-X.; Jin, D.; Jiang, S.; Zhang, Z.-Y.; Bi, L.; Pei, G.-X. The Dose-Effect of Icariin on the Proliferation and Osteogenic Differentiation of Human Bone Mesenchymal Stem Cells. Molecules 2011, 16, 10123-10133. https://doi.org/10.3390/molecules161210123
Fan J-J, Cao L-G, Wu T, Wang D-X, Jin D, Jiang S, Zhang Z-Y, Bi L, Pei G-X. The Dose-Effect of Icariin on the Proliferation and Osteogenic Differentiation of Human Bone Mesenchymal Stem Cells. Molecules. 2011; 16(12):10123-10133. https://doi.org/10.3390/molecules161210123
Chicago/Turabian StyleFan, Jun-Jun, Liang-Guo Cao, Tao Wu, De-Xin Wang, Dan Jin, Shan Jiang, Zhi-Yong Zhang, Long Bi, and Guo-Xian Pei. 2011. "The Dose-Effect of Icariin on the Proliferation and Osteogenic Differentiation of Human Bone Mesenchymal Stem Cells" Molecules 16, no. 12: 10123-10133. https://doi.org/10.3390/molecules161210123