Cleavage of Oligonucleotides Containing a P3’→N5’ Phosphoramidate Linkage Mediated by Single-Stranded Oligonucleotide Templates

Abstract
:1. Introduction
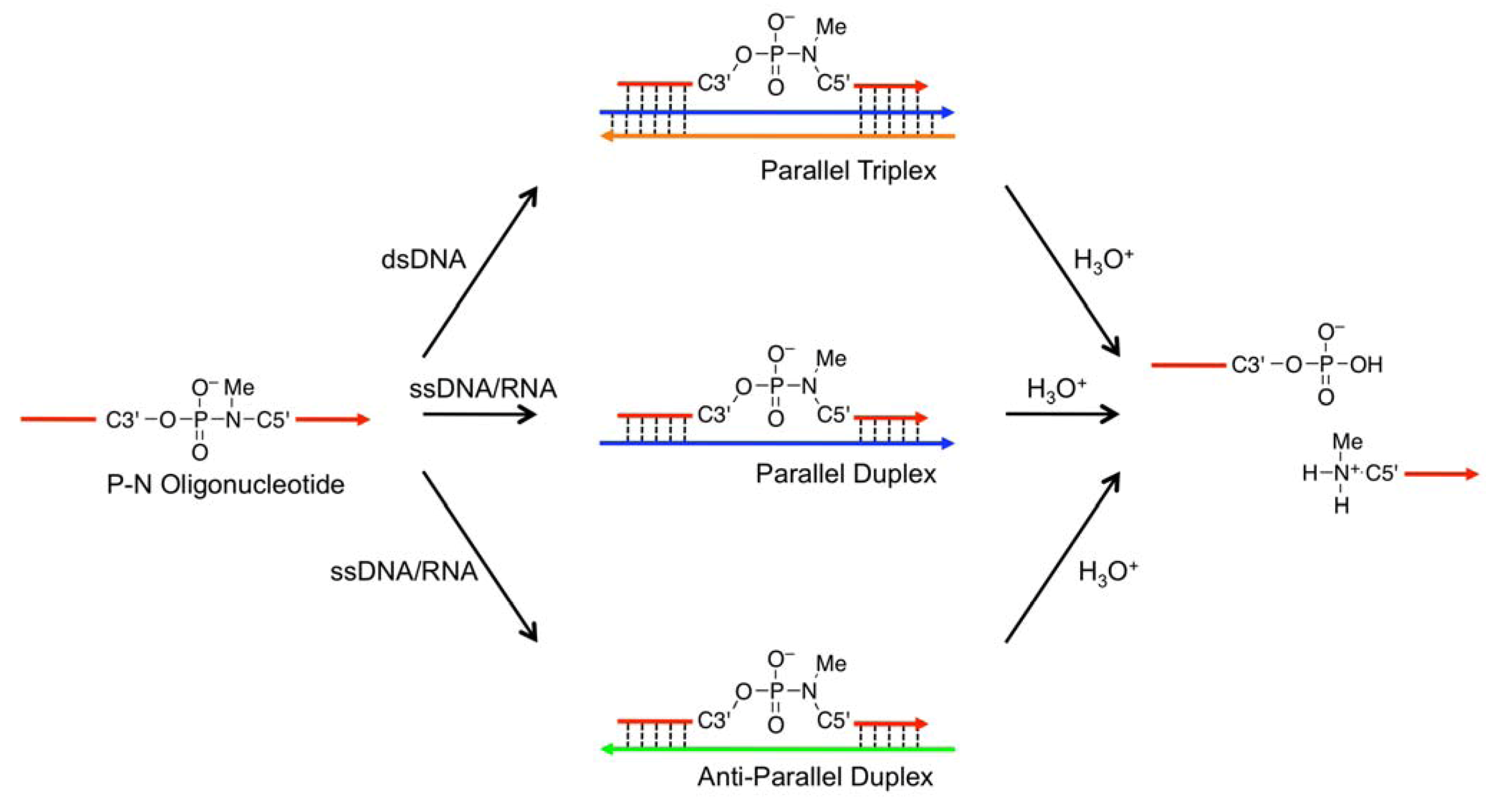
 ) as probes; the TFOs have a P3’→N5’ phosphoramidate (P-N) linkage in the backbone. This linkage was more susceptible to acid-mediated hydrolysis upon triplex formation, and the enhanced susceptibility was due to conformational strain on the P-N linkage induced by triplex formation. Previously, we examined the effects of chemical modifications that alter the microenvironment around the P-N linkage and change the extent of the conformational strain; these chemical modifications had substantial effects on the observed pseudo first-order rate constants (kobss) of the hydrolysis with the dsDNA templates [29]. These findings indicated that when the P-N linkage is subjected to sufficient strain, the linkage promptly breaks upon hybridization to the template. We hypothesized that duplex formation, like triplex formation, could induce conformational strain when oligonucleotides have a certain chemical modification and that such oligonucleotides may be selectively cleaved in the presence of single-stranded templates and, therefore, may be used as probes to detect single-stranded nucleic acids (Figure 2)., Figure 1) in the middle of a sequence with one of two chemical modifications, 2’,4’-BNA/LNA [30,31,32] (designated
) as probes; the TFOs have a P3’→N5’ phosphoramidate (P-N) linkage in the backbone. This linkage was more susceptible to acid-mediated hydrolysis upon triplex formation, and the enhanced susceptibility was due to conformational strain on the P-N linkage induced by triplex formation. Previously, we examined the effects of chemical modifications that alter the microenvironment around the P-N linkage and change the extent of the conformational strain; these chemical modifications had substantial effects on the observed pseudo first-order rate constants (kobss) of the hydrolysis with the dsDNA templates [29]. These findings indicated that when the P-N linkage is subjected to sufficient strain, the linkage promptly breaks upon hybridization to the template. We hypothesized that duplex formation, like triplex formation, could induce conformational strain when oligonucleotides have a certain chemical modification and that such oligonucleotides may be selectively cleaved in the presence of single-stranded templates and, therefore, may be used as probes to detect single-stranded nucleic acids (Figure 2)., Figure 1) in the middle of a sequence with one of two chemical modifications, 2’,4’-BNA/LNA [30,31,32] (designated  ) or 2’,5’-linked DNA [33,34] (designated
) or 2’,5’-linked DNA [33,34] (designated  ), on adjacent residues (Table 1). The reactivity of these oligonucleotides in the presence of single-stranded DNA (ssDNA) or ssRNA templates was compared with their reactivity in the presence of parallel double-stranded DNA (PDD) templates and in the absence of any template. The parallel (Hoogsteen motif) single-stranded DNA and RNA (PSD and PSR, respectively) and anti-parallel (Watson-Crick motif) single-stranded DNA and RNA (ASD and ASR) were prepared as templates (see Table 1 caption). The formation of different motifs of duplexes was expected to have different effects on the reactivity of the hydrolysis depending on the extent of the strain. (5’-amino-2’,4’-BNA), (2’,4’-BNA/LNA), and (2’,5’-linked DNA).
), on adjacent residues (Table 1). The reactivity of these oligonucleotides in the presence of single-stranded DNA (ssDNA) or ssRNA templates was compared with their reactivity in the presence of parallel double-stranded DNA (PDD) templates and in the absence of any template. The parallel (Hoogsteen motif) single-stranded DNA and RNA (PSD and PSR, respectively) and anti-parallel (Watson-Crick motif) single-stranded DNA and RNA (ASD and ASR) were prepared as templates (see Table 1 caption). The formation of different motifs of duplexes was expected to have different effects on the reactivity of the hydrolysis depending on the extent of the strain. (5’-amino-2’,4’-BNA), (2’,4’-BNA/LNA), and (2’,5’-linked DNA).

2. Results
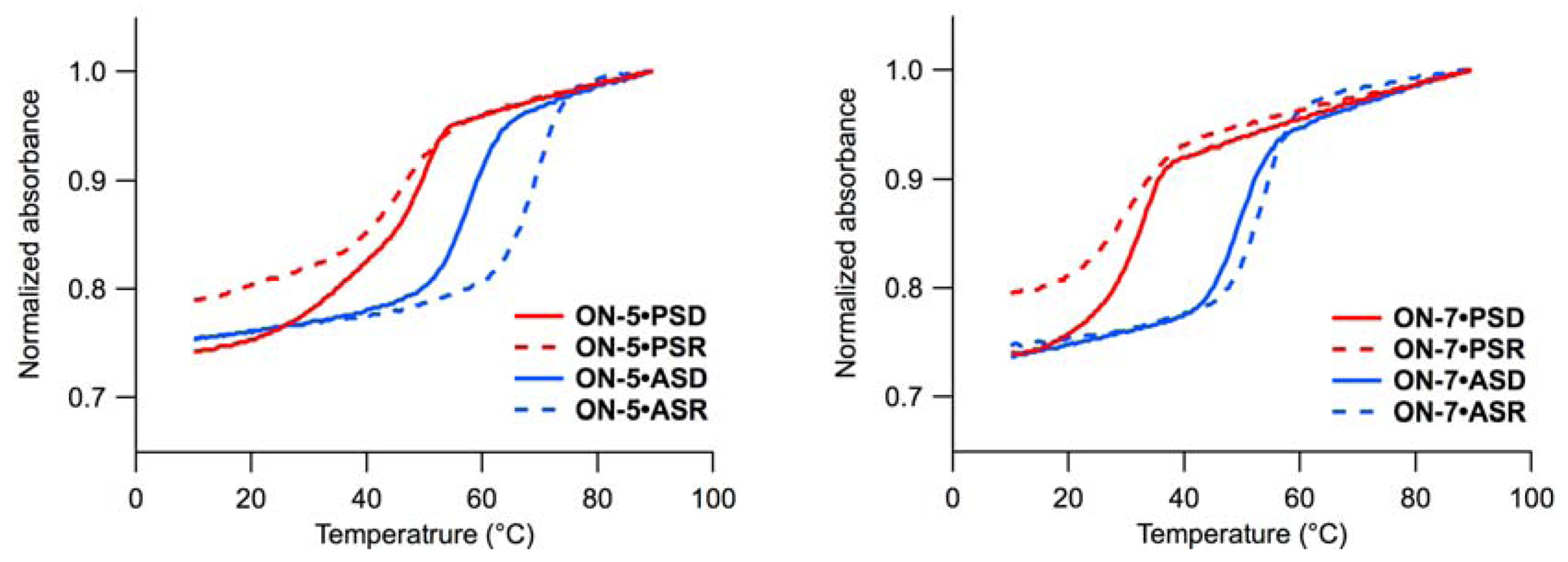
2.1. UV Melting Experiments

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ON | Sequence (5’ to 3’) b | Tm (Δ Tm) in °C with | |||
|---|---|---|---|---|---|
| PSD c | PSR c | ASD c | ASR c | ||
| ON-0 | TTTTTmCTTTmCTmCTmCT | 33 (-) | 34 (-) | 52 (-) | 54 (-) |
| ON-0 d | TTTTTmCTTTmCTmCTmCT | 48 (+15) | n.d. e | 48 (−4) | 36 (−18) |
| ON-1 | TTTTTmCT TmCTmCTmCT | 38 (+5) | 34 (±0) | 54 (+2) | 57 (+3) |
| ON-2 | TTTTTmC TmCTmCTmCT | 37 (+4) | 37 (+3) | 53 (+1) | 60 (+6) |
| ON-3 | TTTTTmCT mCTmCTmCT | 41 (+8) | 40 (+6) | 55 (+3) | 62 (+8) |
| ON-4 | TTTTTmC mCTmCTmCT | 44 (+11) | 44 (+10) | 54 (+2) | 65 (+11) |
| ON-5 | TTTT mCT TmC mCTmCT | 45 (+12) | 46 (+12) | 57 (+5) | 69 (+15) |
| ON-6 | TTTTTmC TmCTmCTmCT | 33 (±0) | 34 (±0) | 47 (−5) | 55 (+1) |
| ON-7 | TTTTTmCT mCTmCTmCT | 31 (−2) | 30 (−4) | 49 (−3) | 53 (−1) |
| ON-8 | TTTTTmC mCTmCTmCT | 31 (−2) | 29 (−5) | 43 (−9) | 51 (−3) |
, 5’-amino-2’,4’-BNA (NMe); , 2’,4’-BNA/LNA; , 2’,5’-linked DNA; mC, 5-MedC; c PSD (parallel single-stranded DNA), 5’-d(AAAAAGAAAGAGAGA)-3’; PSR (parallel single-stranded RNA), 5’-r(AAAAAGAAAGAGAGA)-3’; ASD (anti-parallel single-stranded DNA), 5’-d(AGAGAGAAAGAAAAA)-3’; ASR (anti-parallel single-stranded RNA), 5’-r(AGAGAGAAAGAAAAA)-3’; d Tm measured at pH 4.0, for detail see experimental section; e: Not determined due to low stability (Tm < 25 °C). ) (ON-2–ON-5) stabilized the duplexes in most cases [30,31,32], and the stabilizing effects were more apparent for the duplexes with PSD, PSR, and ASR than those with ASD. Comparisons between ON-2 and ON-3 Tms revealed that a positioned just 3’ of (5’-amino-2’,4’-BNA) stabilized all duplexes to a larger extent than a positioned just 5’ of . Insertion of two residues between and had greater stabilizing effects in all types of duplexes tested here (ON-4 vs. ON-5). Comparison of Tms of duplexes containing 2’,5’-linked DNA ( ) with those of ON-1 revealed that introduction of is destabilizing in most cases. The destabilization was less pronounced for the duplexes with ASR, as reported previously [35,36,37]. The melting curves of the duplexes consisting of PSD and ON-3, ON-4, or ON-8 showed some two-transition character.
) (ON-2–ON-5) stabilized the duplexes in most cases [30,31,32], and the stabilizing effects were more apparent for the duplexes with PSD, PSR, and ASR than those with ASD. Comparisons between ON-2 and ON-3 Tms revealed that a positioned just 3’ of (5’-amino-2’,4’-BNA) stabilized all duplexes to a larger extent than a positioned just 5’ of . Insertion of two residues between and had greater stabilizing effects in all types of duplexes tested here (ON-4 vs. ON-5). Comparison of Tms of duplexes containing 2’,5’-linked DNA ( ) with those of ON-1 revealed that introduction of is destabilizing in most cases. The destabilization was less pronounced for the duplexes with ASR, as reported previously [35,36,37]. The melting curves of the duplexes consisting of PSD and ON-3, ON-4, or ON-8 showed some two-transition character.2.2. Hydrolysis Experiments
| ON | kobs × 103 (s−1) in the presence of | ||||
|---|---|---|---|---|---|
| No template b | PDD b,c | PSD | ASD | ASR | |
| ON-1 | 0.027 ± 0.005 | 0.77 ± 0.03 | 0.57 ± 0.09 | 0.14 ± 0.01 | 0.17 ± <0.01 |
| ON-2 | 0.017 ± 0.006 | 0.051 ± 0.013 | 0.025 ± 0.001 | 0.020 ± 0.002 | 0.032 ± 0.001 |
| ON-3 | 0.038 ± 0.012 | 1.4 ± 0.4 | 0.60 ± 0.05 | 0.10 ± <0.01 | 0.19 ± <0.01 |
| ON-4 | 0.026 ± 0.003 | 0.044 ± 0.025 | 0.032 ± 0.003 | 0.015 ± 0.003 | n.d. d |
| ON-5 | 0.029 ± 0.004 | 1.3 ± 0.4 | 0.86 ± 0.11 | 0.24 ± 0.02 | 0.19 ± <0.01 |
| ON-6 | 0.066 ± 0.024 | n.d. d | 0.021 ± 0.004 | 0.021 ± 0.001 | 0.011 ± 0.001 |
| ON-7 | 0.022 ± 0.002 | 2.1 ± 0.1 | 0.83 ± 0.03 | 0.29 ± <0.01 | 0.18 ± <0.01 |
| ON-8 | 0.058 ± 0.003 | 0.022 ± 0.012 | 0.078 ± 0.003 | 0.035 ± 0.002 | 0.024 ± <0.001 |
| ON | kobs × 103 (s−1) in the presence of | ||
|---|---|---|---|
| No template | PSD | ASR | |
| ON-1 | 0.011 ± <0.001 | 0.27 ± 0.06 | 0.022 ± 0.001 |
| ON-2 | 0.011 ± 0.002 | 0.0067 ± 0.0006 | 0.0059 ± 0.0006 |
| ON-3 | 0.0072 ± 0.0010 | 0.19 ± 0.05 | 0.032 ± 0.002 |
| ON-4 | 0.0062 ± 0.0011 | n.d. b | n.d. b |
| ON-5 | 0.0073 ± 0.0015 | 0.26 ± 0.01 | 0.041 ± <0.001 |
| ON-6 | 0.0067 ± 0.0013 | n.d. b | n.d. b |
| ON-7 | 0.0080 ± 0.0008 | 0.68 ± 0.04 | 0.037 ± 0.005 |
| ON-8 | 0.0070 ± 0.0005 | n.d. b | n.d. b |

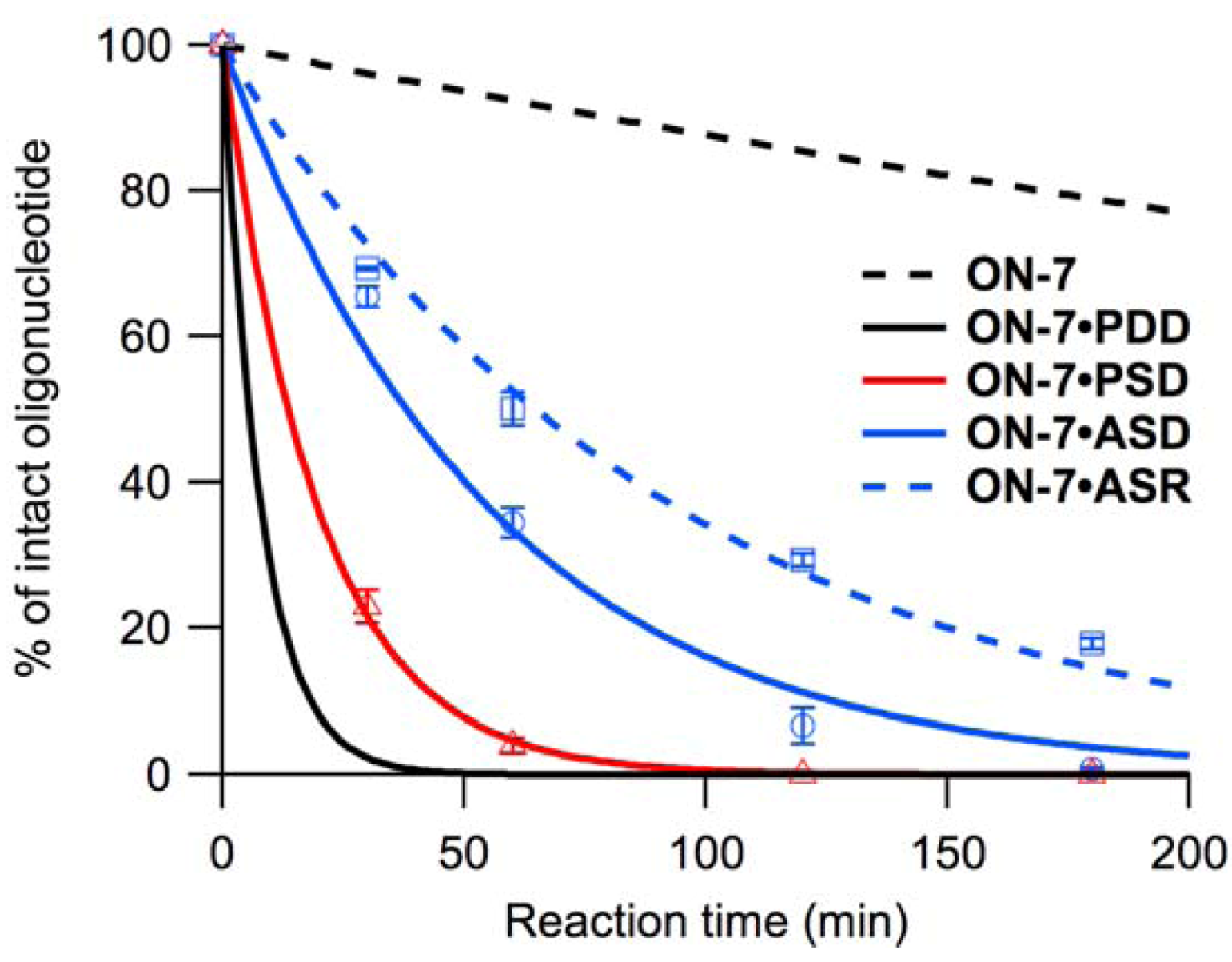
2.2.1. Reactivity on Parallel Single-Stranded DNA
to the 5’-neighboring residue of (the sequence 5’- -3’ found in ON-2 and ON-4) resulted in inactivation of hydrolysis upon hybridization to PSD (Figure S2). added to the 3’-neighboring residue of (5’-T -3’) had little effect, resulting in equivalent kobs for ON-1 and ON-3. Accelerated hydrolysis of ON-5 on PSD was observed as non-neighboring residual effects of . The sequence 5’- -3’ found in ON-6 and ON-8 eliminated the acceleration associated with hybridization to PSD, while hydrolysis of ON-7 (sequence 5’-T -3’) was accelerated (Figure 4). At 20 °C, ON-1–ON-8 were less reactive, but the reactivity of ON-7 was least affected, and reactivities of ON-1, ON-3, and ON-5 were equivalent in the presence of PSD (Table 3, Figure S3).2.2.2. Reactivity on Anti-Parallel Single-Stranded DNA and RNA
3. Discussion
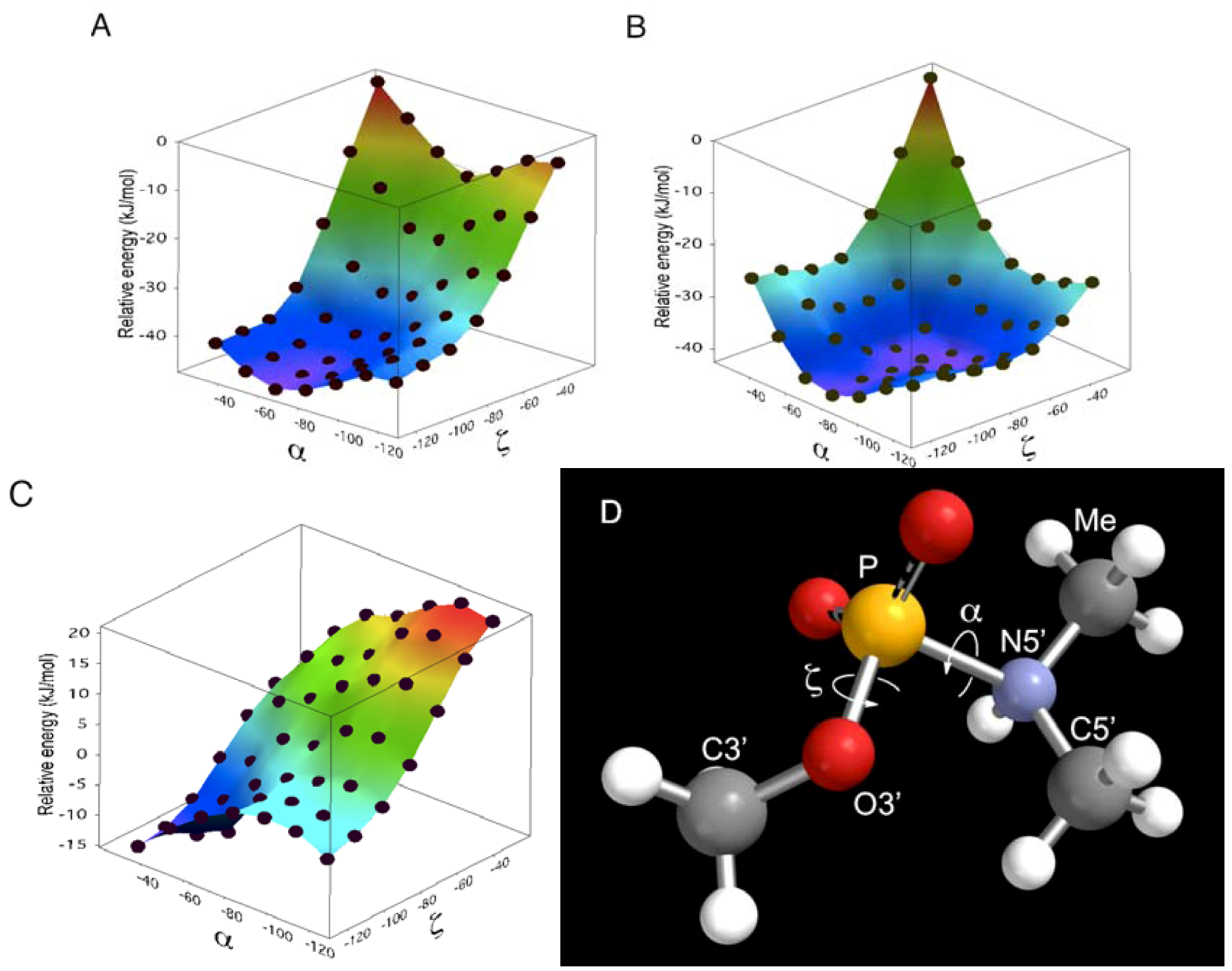
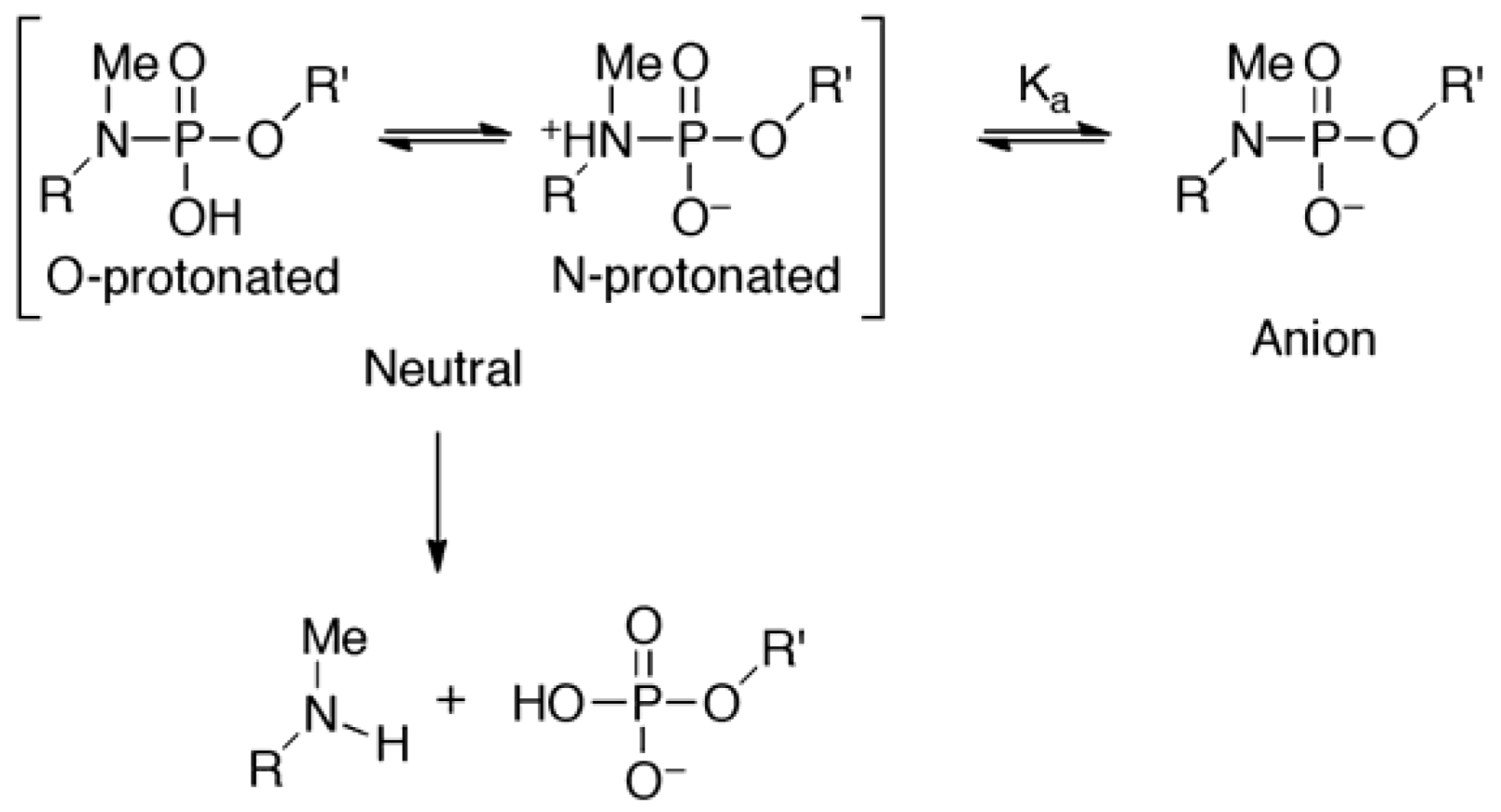
 did not prohibit α and ζ dihedral angles from adopting −sc orientations (Figure 5, Figure S5). However, the high reactivity in triplexes and parallel Hoogsteen duplexes may have been due to a change in the preferred dihedral angles, which would make the phosphoramidates more basic. The quantum chemical calculations using one model compound (N,N,O-trimethylphosphoramidate, R = R’ = Me in Scheme 1) revealed the relative stability between the neutral form (N-protonated) and the anionic form of the phosphoramidate (Scheme 1) as functions of α and ζ.
did not prohibit α and ζ dihedral angles from adopting −sc orientations (Figure 5, Figure S5). However, the high reactivity in triplexes and parallel Hoogsteen duplexes may have been due to a change in the preferred dihedral angles, which would make the phosphoramidates more basic. The quantum chemical calculations using one model compound (N,N,O-trimethylphosphoramidate, R = R’ = Me in Scheme 1) revealed the relative stability between the neutral form (N-protonated) and the anionic form of the phosphoramidate (Scheme 1) as functions of α and ζ. 
 is known to affect the sugar conformation of 3’-adjacent nucleotide inducing C3’-endo conformation [50,51]. Although such an effect is not the case in ON-2 and ON-4 because is pre-locked to C3’-endo conformation due to the 2’,4’-bridge moiety [26], at 5’-adjacent of the phosphoramidate will have structurally affected the preferred conformation of the phosphoramidate, which resulted in inactivation of ON-2 and ON-4 in the presence of templates.
is known to affect the sugar conformation of 3’-adjacent nucleotide inducing C3’-endo conformation [50,51]. Although such an effect is not the case in ON-2 and ON-4 because is pre-locked to C3’-endo conformation due to the 2’,4’-bridge moiety [26], at 5’-adjacent of the phosphoramidate will have structurally affected the preferred conformation of the phosphoramidate, which resulted in inactivation of ON-2 and ON-4 in the presence of templates.4. Experimental Section
4.1. Preparation of Oligonucleotides
2.2. UV Melting Experiments
4.3. Hydrolysis Experiments
4.4. Molecular Modeling and Computation
. The structures generated were exported to MacroModel 9.1TM (Schrödinger, LLC, New York, NY, USA). An energy minimization calculation was performed for each structure using 1) AMBER* as a force field, 2) the GB/SA solvation model of water, and 3) the PRCG method to obtain structures optimized to within a gradient of 0.05 kJ/molÅ. Finally, the 5’-oxygen of 2’,4’-BNA/LNA in ON-1 of the optimized structures was replaced by nitrogen attached to a methyl group to obtain the molecular models (Figure 5, Figure S5).5. Conclusions
Supplementary Materials
Acknowledgments
Conflict of Interest
References
- Xu, Y.; Karalkar, N.B.; Kool, E.T. Nonenzymatic autoligation in direct three-color detection of RNA and DNA point mutations. Nat. Biotechnol. 2001, 19, 148–152. [Google Scholar] [CrossRef]
- Sando, S.; Kool, E.T. Quencher as leaving group: Efficient detection of DNA-joining reactions. J. Am. Chem. Soc. 2002, 124, 2096–2097. [Google Scholar] [CrossRef]
- Abe, H.; Kool, E.T. Destabilizing universal linkers for signal amplification in self-ligating probes for RNA. J. Am. Chem. Soc. 2004, 126, 13980–13986. [Google Scholar] [CrossRef]
- Ficht, S.; Dose, C.; Seitz, O. As fast and selective as enzymatic ligations: Unpaired nucleobases increase the selectivity of DNA-controlled native chemical PNA ligation. ChemBioChem 2005, 6, 2098–2103. [Google Scholar] [CrossRef]
- Dose, C.; Ficht, S.; Seitz, O. Reducing product inhibition in DNA-template-controlled ligation reactions. Angew. Chem. Int. Ed. 2006, 45, 5369–5373. [Google Scholar] [CrossRef]
- Abe, H.; Kool, E.T. Flow cytometric detection of specific RNAs in native human cells with quenched autoligating FRET probes. Proc. Natl. Acad. Sci. USA 2006, 103, 263–268. [Google Scholar] [CrossRef]
- Ogasawara, S.; Fujimoto, K. SNP genotyping by using photochemical ligation. Angew. Chem. Int. Ed. 2006, 45, 4512–4515. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Noguchi, Y.; Sato, H.; Fujimoto, K. Template-directed DNA photoligation in rapid and selective detection of RNA point mutations. ChemBioChem 2006, 7, 598–601. [Google Scholar] [CrossRef]
- Peng, X.; Greenberg, M.M. Facile SNP detection using bifunctional, cross-linking oligonucleotide probes. Nucleic Acids Res. 2008, 26, e31. [Google Scholar]
- Abe, H.; Kondo, Y.; Jinmei, H.; Abe, N.; Furukawa, K.; Uchiyama, A.; Tsuneda, S.; Aikawa, K.; Matsumoto, I.; Ito, Y. Rapid DNA chemical ligation for amplification of RNA and DNA signal. Bioconjug. Chem. 2008, 19, 327–333. [Google Scholar] [CrossRef]
- Dose, C.; Seitz, O. Single nucleotide specific detection of DNA by native chemical ligation of fluorescence labeled PNA-probes. Bioorg. Med. Chem. 2008, 16, 65–77. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Seitz, O. DNA-catalyzed transfer of a reporter group. J. Am. Chem. Soc. 2006, 128, 15596–15597. [Google Scholar] [CrossRef]
- Grossmann, T.N.; Röglin, L.; Seitz, O. Angew. Chem. Int. Ed. 2008, 47, 7119–7123. [CrossRef]
- Grossmann, T.N.; Seitz, O. Nucleic acid templated reactions: Consequences of probe reactivity and readout strategy for amplified signaling and sequence selectivity. Chem. Eur. J. 2009, 15, 6723–6730. [Google Scholar] [CrossRef]
- Brunner, J.; Mokhir, A.; Kraemer, R. DNA-templated metal catalysis. J. Am. Chem. Soc. 2003, 125, 12410–12411. [Google Scholar] [CrossRef]
- Boll, I.; Krämer, R.; Brunner, J.; Mokhir, A. Templated metal catalysis for single nucleotide specific DNA sequence detection. J. Am. Chem. Soc. 2005, 127, 7849–7856. [Google Scholar] [CrossRef]
- Franzini, R.M.; Kool, E.T. Efficient nucleic acid detection by templated reductive quencher release. J. Am. Chem. Soc. 2009, 131, 16021–16023. [Google Scholar] [CrossRef]
- Cai, J.; Li, X.; Yue, X.; Taylor, J.S. Nucleic acid-triggered fluorescent probe activation by the Staudinger reaction. J. Am. Chem. Soc. 2004, 126, 16324–16325. [Google Scholar]
- Cai, J.; Li, X.; Taylor, J.S. Improved nucleic acid triggered probe activation through the use of a 5-thiomethyluracil peptide nucleic acid building block. Org. Lett. 2005, 7, 751–754. [Google Scholar] [CrossRef]
- Pianowski, Z.L.; Winssinger, N. Fluorescence-based detection of single nucleotide permutation in DNA via catalytically templated reaction. Chem. Commun. 2007, 3820–3822. [Google Scholar] [CrossRef]
- Franzini, R.M.; Kool, E.T. 7-Azidomethoxy-coumarins as profluorophores for templated nucleic acid detection. ChemBioChem 2008, 9, 2981–2988. [Google Scholar] [CrossRef]
- Franzini, R.M.; Kool, E.T. Organometallic activation of a fluorogen for templated nucleic acid detection. Org. Lett. 2008, 10, 2935–2938. [Google Scholar] [CrossRef]
- Furukawa, K.; Abe, H.; Wang, J.; Uda, M.; Koshino, H.; Tsuneda, S.; Ito, Y. Reduction-triggered red fluorescent probes for dual-color detection of oligonucleotide sequences. Org. Biomol. Chem. 2009, 7, 671–677. [Google Scholar] [CrossRef]
- Prusty, D.K.; Herrmann, A. A fluorogenic reaction based on heavy-atom removal for ultrasensitive DNA detection. J. Am. Chem. Soc. 2010, 132, 12197–12199. [Google Scholar] [CrossRef]
- Li, X.; Liu, D.R. DNA-templated organic synthesis: Nature’s strategy for controlling chemical reactivity applied to synthetic molecules. Angew. Chem. Int. Ed. 2004, 43, 4848–4870. [Google Scholar] [CrossRef]
- Obika, S.; Nakagawa, O.; Hiroto, A.; Hari, Y.; Imanishi, T. Synthesis and properties of a novel bridged nucleic acid with a P3’→N5’ phosphoramidate linkage, 5’-amino-2’,4’-BNA. Chem. Commun. 2003, 2202–2203. [Google Scholar]
- Obika, S.; Tomizu, M.; Negoro, Y.; Osakai, T.; Orita, A.; Ueyama, Y.; Nakagawa, O.; Imanishi, T. Acid-mediated cleavage of oligonucleotide P3’→N5’ phosphoramidates triggered by sequence-specific triplex formation. Nucleos. Nucleot. Nucleic Acids 2007, 26, 893–896. [Google Scholar] [CrossRef]
- Obika, S.; Tomizu, M.; Negoro, Y.; Orita, A.; Nakagawa, O.; Imanishi, T. Double-stranded DNA-templated digestion triggered by triplex formation. ChemBioChem 2007, 8, 1924–1928. [Google Scholar] [CrossRef]
- Ito, K.R.; Kodama, T.; Tomizu, M.; Negoro, Y.; Orita, A.; Osaki, T.; Hosoki, N.; Tanaka, T.; Imanishi, T.; Obika, S. Double-stranded DNA-templated cleavage of oligonucleotides containing a P3’→N5’ linkage triggered by triplex formation: The effects of chemical modifications and remarkable enhancement in reactivity. Nucleic Acids Res. 2010, 38, 7332–7342. [Google Scholar]
- Obika, S.; Nanbu, D.; Hari, Y.; Andoh, J.; Morio, K.; Doi, T.; Imanishi, T. Stability and structural features of the duplexes containing nucleoside analogues with a fixed N-type conformation, 2’-O,4’-C-methyleneribonucleosides. Tetrahedron Lett. 1998, 39, 5401–5404. [Google Scholar] [CrossRef]
- Singh, S.K.; Nielsen, P.; Koshkin, A.A.; Wengel, J. LNA (locked nucleic acids): Synthesis and high-affinity nucleic acid recognition. Chem. Commun. 1998, 455–456. [Google Scholar]
- Bhattacharyya, J.; Maiti, S.; Muhuri, S.; Nakano, S.; Miyoshi, D.; Sugimoto, N. Effect of locked nucleic acid modifications on the thermal stability of noncanonical DNA structure. Biochemistry 2011, 50, 7414–7425. [Google Scholar] [CrossRef]
- Rizzo, C.J.; Dougherty, J.P.; Breslow, R. 3’-Deoxy-2’-phosphoramidites of adenosine and 5-methyluridine used for the solid phase synthesis of unnatural 3’-deoxy-2’-5’’-oligonucleotides. Tetrahedron Lett. 1992, 33, 4129–4132. [Google Scholar]
- Dougherty, J.P.; Rizzo, C.J.; Breslow, R. Oligodeoxynucleotides that contain 2’,5’’ linkages: Synthesis and hybridization properties. J. Am. Chem. Soc. 1992, 114, 6254–6255. [Google Scholar] [CrossRef]
- Giannaris, P.A.; Damha, M.J. Oligoribonucleotides containing 2’,5’-phosphodiester linkages exhibit binding selectivity for 3’,5’-RNA over 3’,5’-ssDNA. Nucleic Acids Res. 1993, 21, 4742–4749. [Google Scholar] [CrossRef]
- Prakash, T.P.; Jung, K.; Switzer, C. RNA recognition by the 2’-structural isomer of DNA. Chem. Commun. 1996, 1793–1794. [Google Scholar]
- Sheppard, T.L.; Breslow, R.C. Selective binding of RNA, not DNA, by complementary 2’,5’-linked DNA. J. Am. Chem. Soc. 1996, 118, 9810–9811. [Google Scholar] [CrossRef]
- Raghunathan, G.; Miles, H.T.; Sasisekharan, V. Parallel nucleic acid helices with Hoogsteen base pairing: Symmetry and structure. Biopolymers 1994, 34, 1573–1581. [Google Scholar] [CrossRef]
- Singleton, S.F.; Dervan, P.B. Influence of pH on the equilibrium association constants for oligodeoxyribonucleotide-directed triple helix formation at single DNAsites. Biochemistry 1992, 31, 10995–11003. [Google Scholar]
- Hashem, G.M.; Wen, J.; Do, Q.; Gray, D.M. Evidence from CD spectra and melting temperatures for stable Hoogsteen-paired oligomer duplexes derived from DNA and hybrid triplexes. Nucleic Acids Res. 1999, 3371–3379. [Google Scholar]
- Sugimoto, N.; Wu, P.; Hara, H.; Kawamoto, Y. pH and cation effects on the properties of parallel pyrimidine motif DNA triplexes. Biochemistry 2001, 40, 9396–9405. [Google Scholar] [CrossRef]
- Roberts, R.W.; Crothers, D.M. Stability and properties of double and triple helices: Dramatic effects of RNA or DNA backbone composition. Science 1992, 258, 1463–1466. [Google Scholar]
- Han, H.; Dervan, P.B. Sequence-specific recognition of double helical RNA and RNA•DNA by triple helix formation. Proc. Natl. Acad. Sci. USA 1993, 90, 3806–3810. [Google Scholar] [CrossRef]
- Escudé, C.; François, J.; Sun, J.; Ott, G.; Sprinzl, M.; Garestier, T.; Hélène, C. Stability of triple helices containing RNA and DNA strands: experimental and molecular modeling studies. Nucleic Acids Res. 1993, 21, 5547–5553. [Google Scholar] [CrossRef]
- Han, H.; Dervan, P.B. Different conformational families of pyrimidine•purine•pyrimidine triple helices depending on backbone composition. Nucleic Acids Res. 1994, 22, 2837–2844. [Google Scholar] [CrossRef]
- Xiong, Y.; Sundaralingam, M. Crystal structure and conformation of a DNA-RNA hybrid duplex with a polypurine RNA strand: d(TTCTTBr5CTTC)-r(GAAGAAGAA). Structure 1998, 6, 1493–1501. [Google Scholar] [CrossRef]
- MacKerell, A.D., Jr. Contribution of the intrinsic mechanical energy of the phosphodiester linkage to the relative stability of the A, BI, and BII forms of duplex DNA. J. Phys. Chem. B 2009, 113, 3235–3244. [Google Scholar] [CrossRef]
- Bhaumik, S.R.; Chary, K.V.R.; Govil, G.; Liu, K.; Miles, H.T. A novel palindromic triple-stranded structure fromed by homopyrimidine dodecamer d-CTTCTCCTCTTC and homopurine hexamer d-GAAGAG. Nucleic Acids Res. 1998, 26, 2981–2988. [Google Scholar] [CrossRef]
- Rhee, S.; Han, Z.; Liu, K.; Miles, H.T.; Davies, D.R. Structure of a triple helical DNA with a triplex-duplex junction. Biochemistry 1999, 38, 16810–16815. [Google Scholar] [CrossRef]
- Petersen, M.; Nielsen, C.B.; Nielsen, K.E.; Jensen, G.A.; Bondensgaard, K.; Singh, S.K.; Rajwanshi, V.K.; Koshkin, A.A.; Dahl, B.M.; Wengel, J.; et al. The conformations of locked nucleic acids (LNA). J. Mol. Recognit. 2000, 13, 44–53. [Google Scholar] [CrossRef]
- Jensen, G.A.; Singh, S.K.; Kumar, R.; Wengel, J.; Jacobsen, J.P. A comparison of the solution structures of an LNA:DNA duplex and the unmodified DNA:DNA duplex. J. Chem. Soc. Perkin Trans. 2 2001, 1224–1232. [Google Scholar]
- Sample Availability: Not available.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ito, K.R.; Kodama, T.; Makimura, F.; Hosoki, N.; Osaki, T.; Orita, A.; Imanishi, T.; Obika, S. Cleavage of Oligonucleotides Containing a P3’→N5’ Phosphoramidate Linkage Mediated by Single-Stranded Oligonucleotide Templates. Molecules 2011, 16, 10695-10708. https://doi.org/10.3390/molecules161210695
Ito KR, Kodama T, Makimura F, Hosoki N, Osaki T, Orita A, Imanishi T, Obika S. Cleavage of Oligonucleotides Containing a P3’→N5’ Phosphoramidate Linkage Mediated by Single-Stranded Oligonucleotide Templates. Molecules. 2011; 16(12):10695-10708. https://doi.org/10.3390/molecules161210695
Chicago/Turabian StyleIto, Kosuke Ramon, Tetsuya Kodama, Futaba Makimura, Noritsugu Hosoki, Tomohisa Osaki, Ayako Orita, Takeshi Imanishi, and Satoshi Obika. 2011. "Cleavage of Oligonucleotides Containing a P3’→N5’ Phosphoramidate Linkage Mediated by Single-Stranded Oligonucleotide Templates" Molecules 16, no. 12: 10695-10708. https://doi.org/10.3390/molecules161210695