Tannins of Constant Structure in Medicinal and Food Plants—Hydrolyzable Tannins and Polyphenols Related to Tannins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Type A Tannins and Related Polyphenols of Constant Structures and Compositions
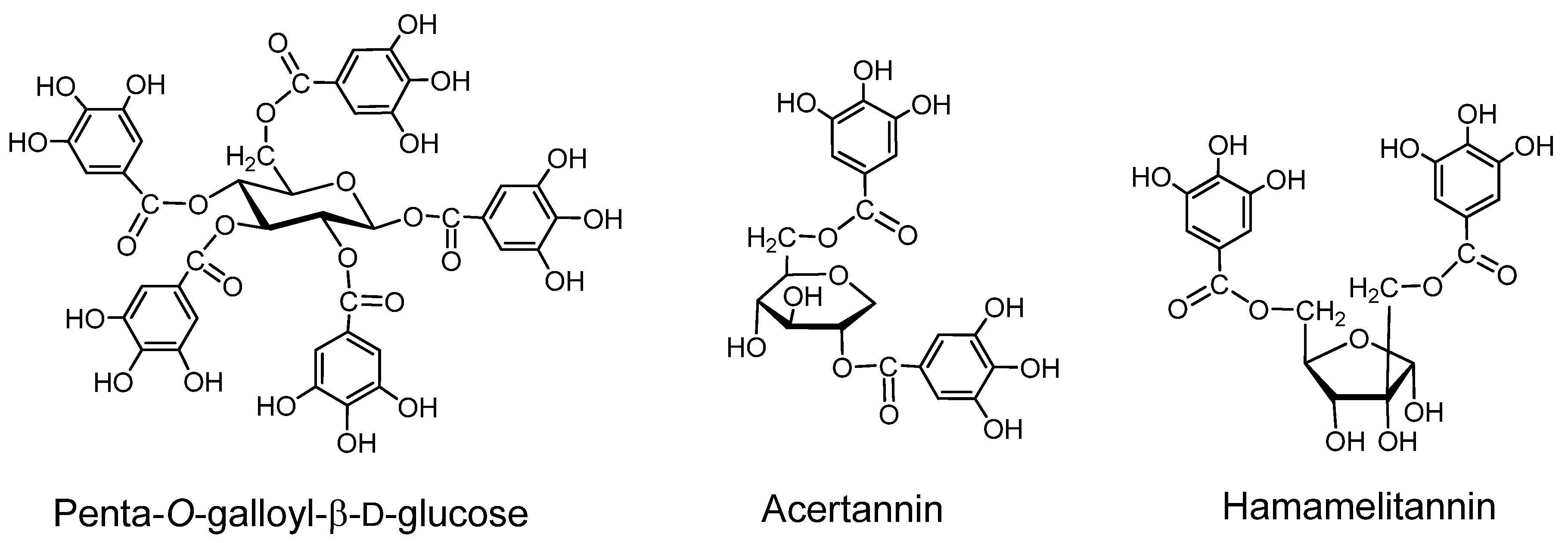
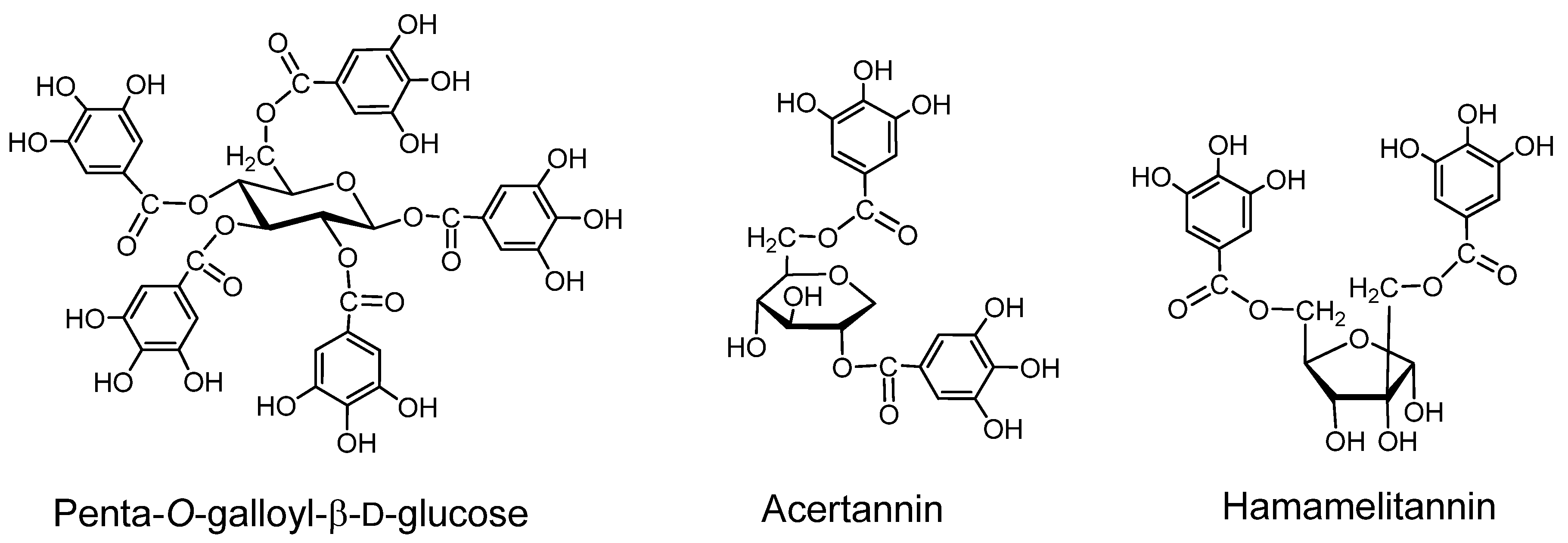
2.1. Gallotannins of Type A (Figure 1)
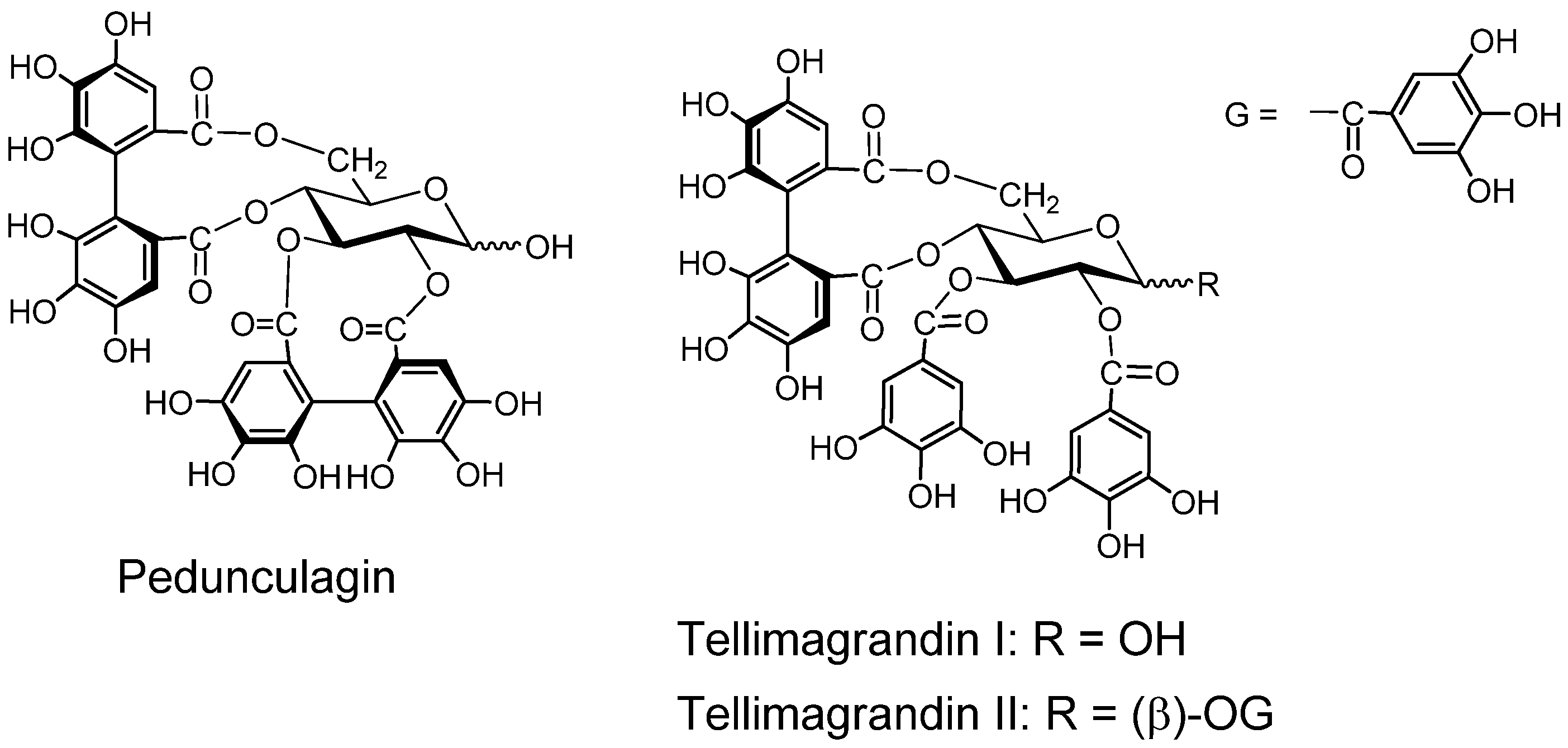
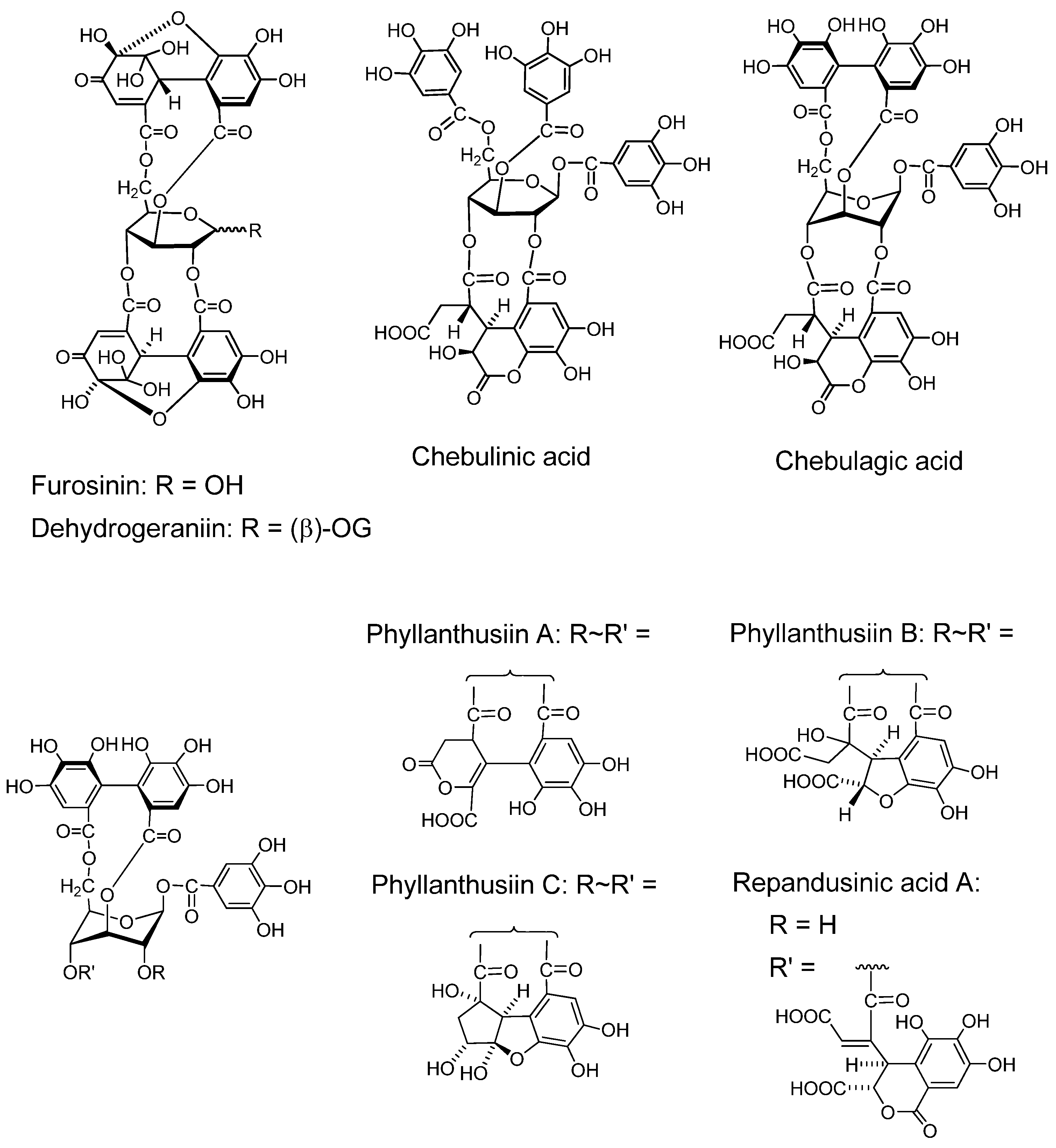
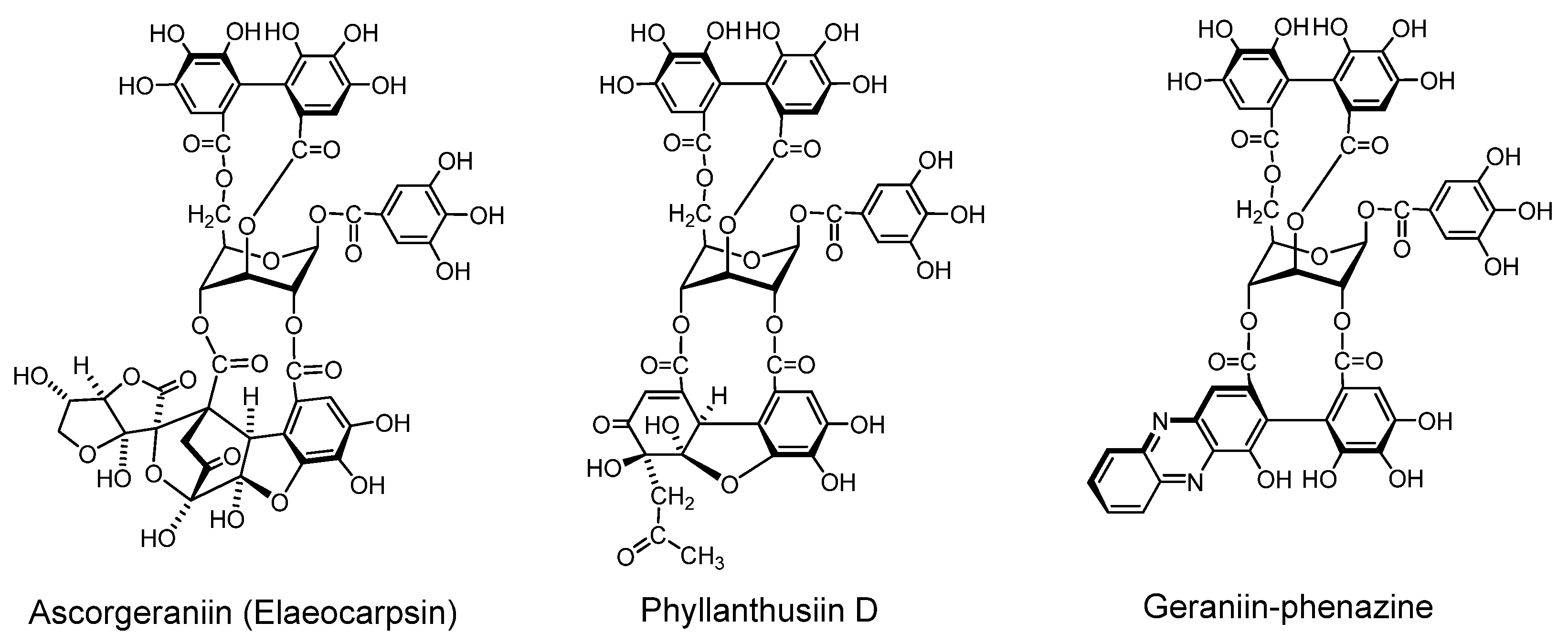
2.2. Ellagitannin, Dehydroellagitannins and Their Oxidatively Transformed Analogs
2.2.1. Ellagitannin and dehydroellagitannin monomers and their oxidized congeners


2.2.2. Condensation of dehydroellagitannin with coexisting compound under mild conditions (Figure 5)
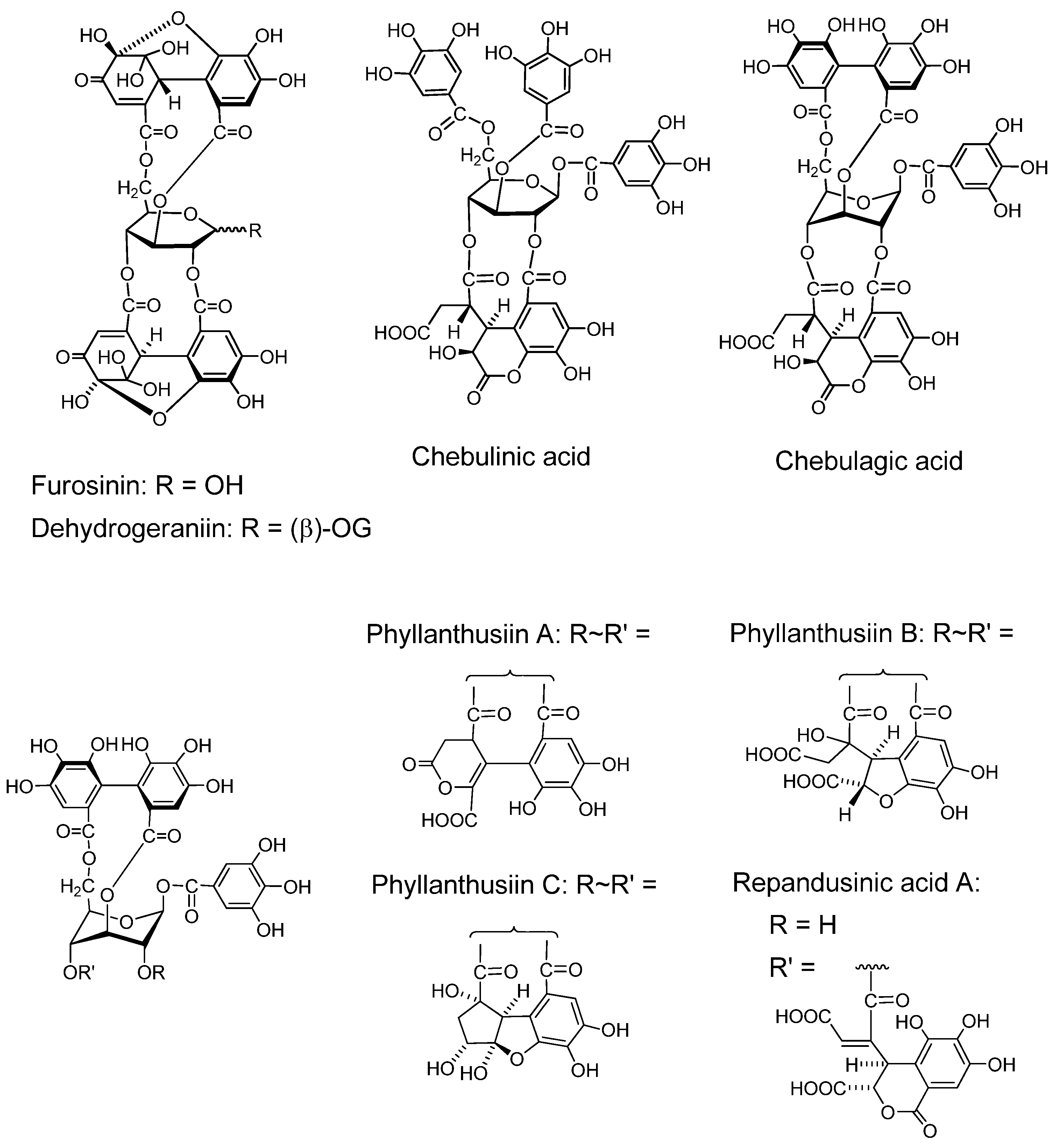

2.2.3. Ellagitannin oligomers with notable pharmacological activities
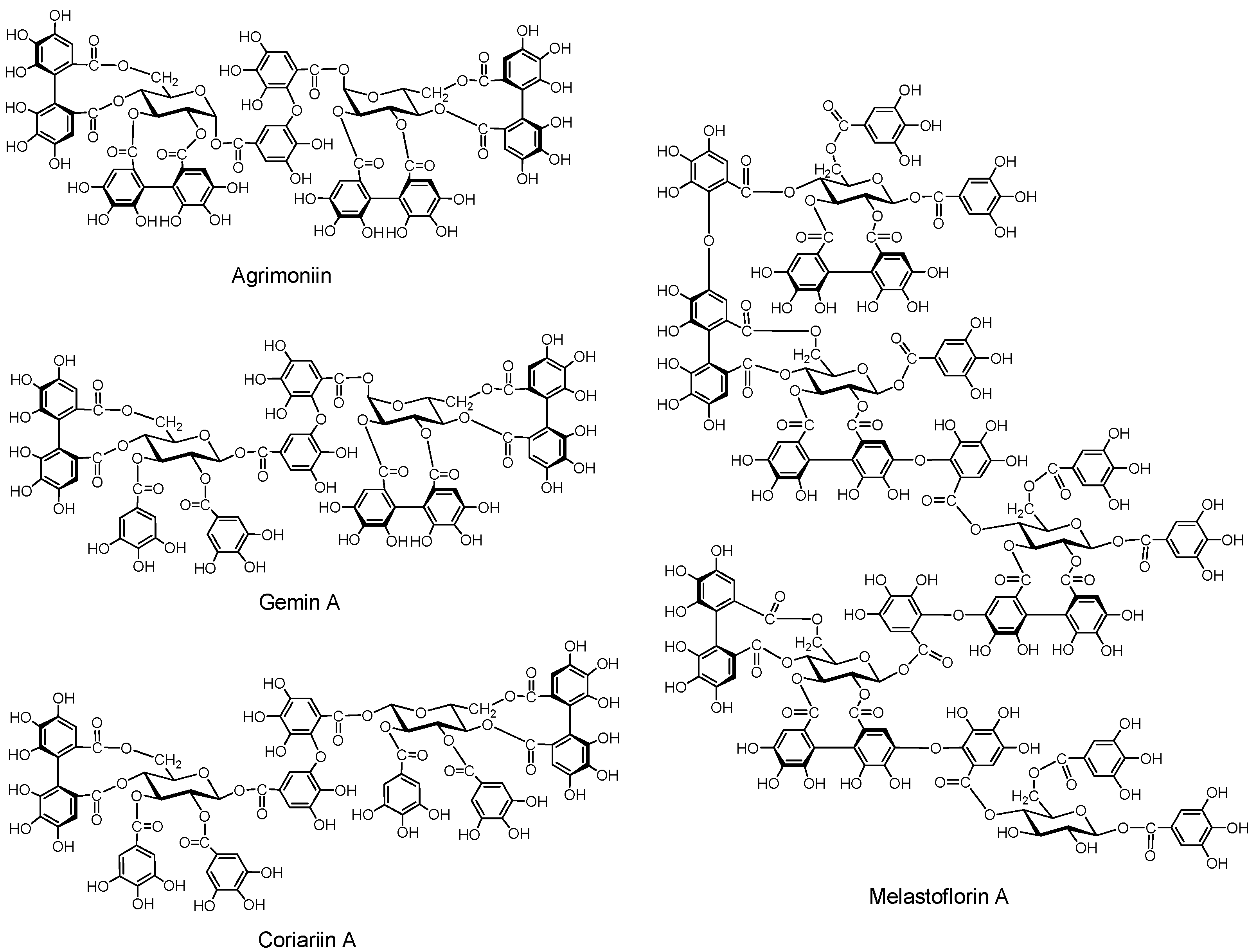
2.2.3.1. Ellagitannin and dehydroellagitannin oligomers (Figure 6)

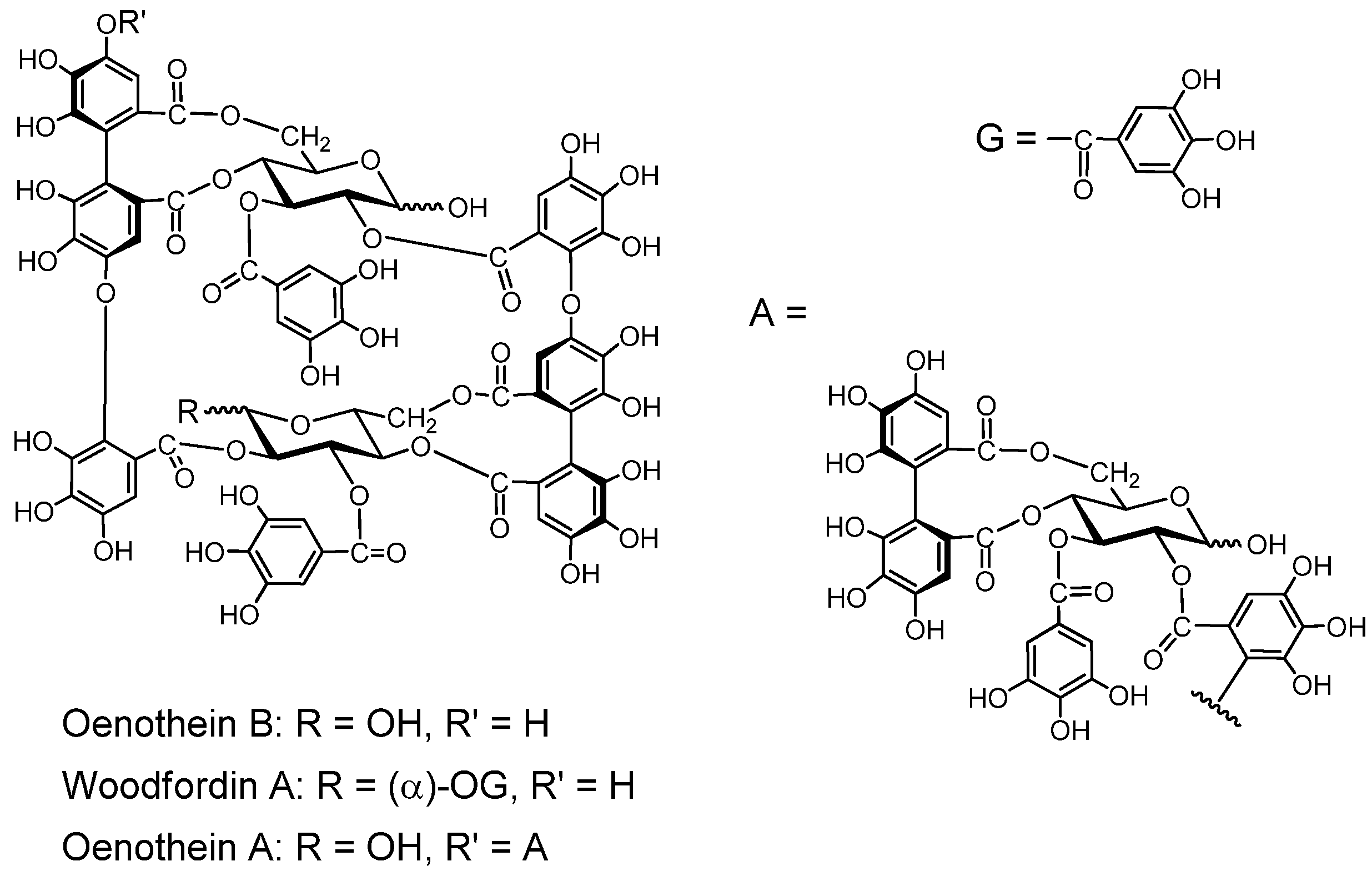
2.2.3.2. Macrocyclic Oligomers (Figure 7).

2.2.3.3. C-Glucosidic Ellagitannins and Complex Tannins
2.3. Transformations of the Type A Tannins in Plants, during Extraction and upon Ingestion of the Medicines and Foods Containing Them
2.3.1. Transformation of hydrolyzable tannin structures in young leaf of a woody plant
2.3.2. Transformation of ellagitannin structures during extraction
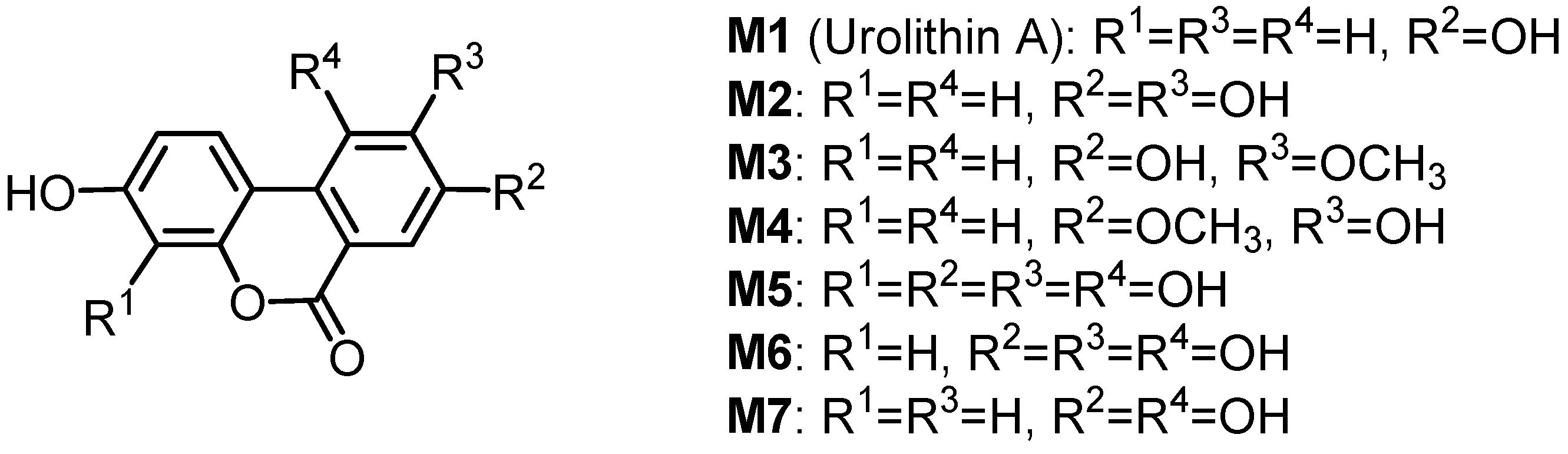
2.3.3. Transformation of ellagitannin structures upon ingestion

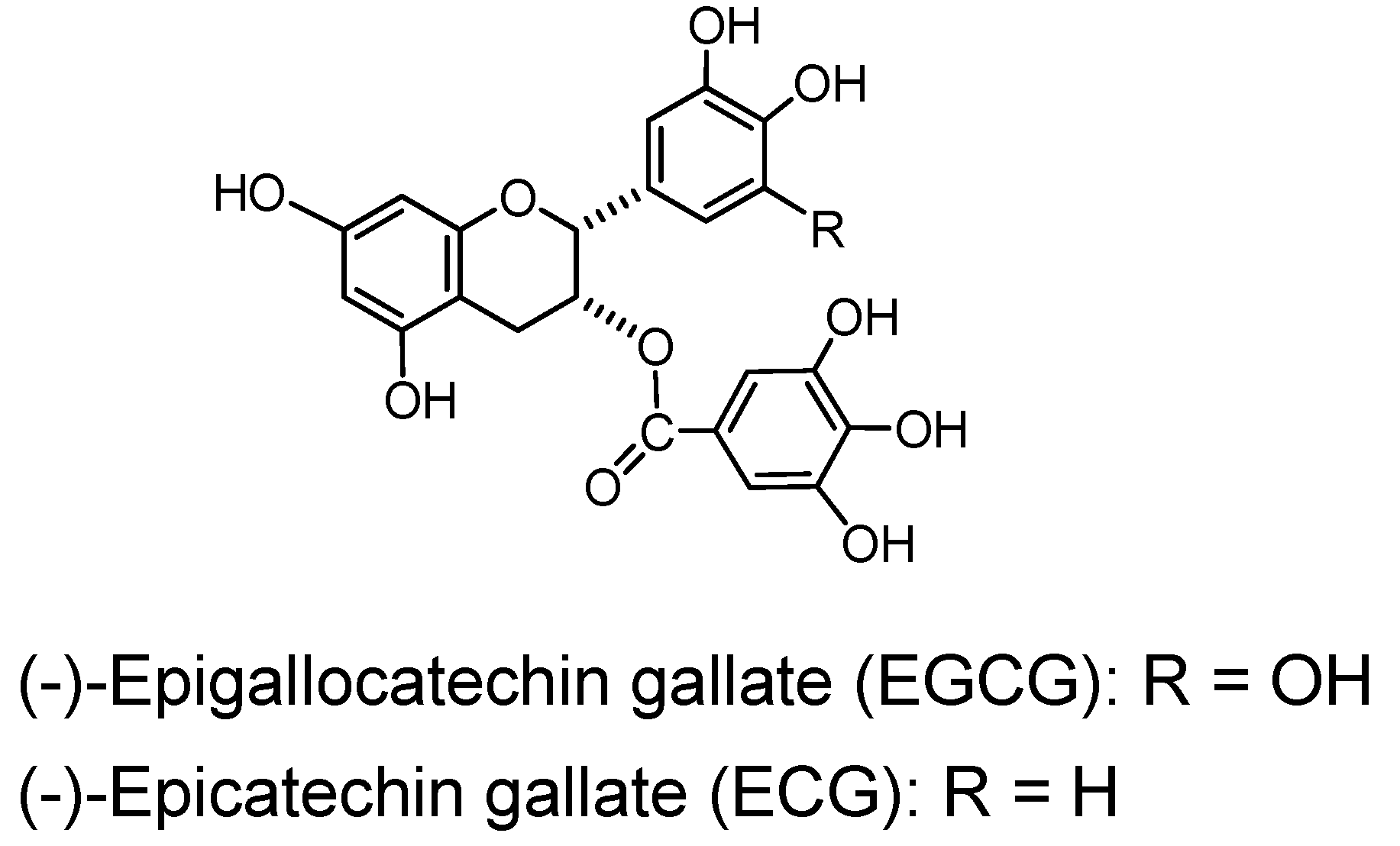
2.4. Polyhydroxyflavan Gallates

2.5. Pharmacological Activities of Ellagitannins and Monomeric Polyhydroxyflavan Gallates
2.5.1. Antimicrobial activities
2.5.2. Antitumor activities
2.5.2.1. Inhibition of the mutagenicity of carcinogens
2.5.2.2. Inhibition of tumor promotion
2.5.2.3. Host-mediated antitumor activity of ellagitannin oligomers
2.6. Caffeic Acid Esters (Caffetannins)
2.6.1. Caffeoyl esters of quinic acid (Figure 10).

2.6.2. Caffeic acid oligomers
2.7. Stilbenoids (Figure 11).
2.7.1. Stilbenoid monomers and their glucosides

2.7.2. Stilbenoid oligomers
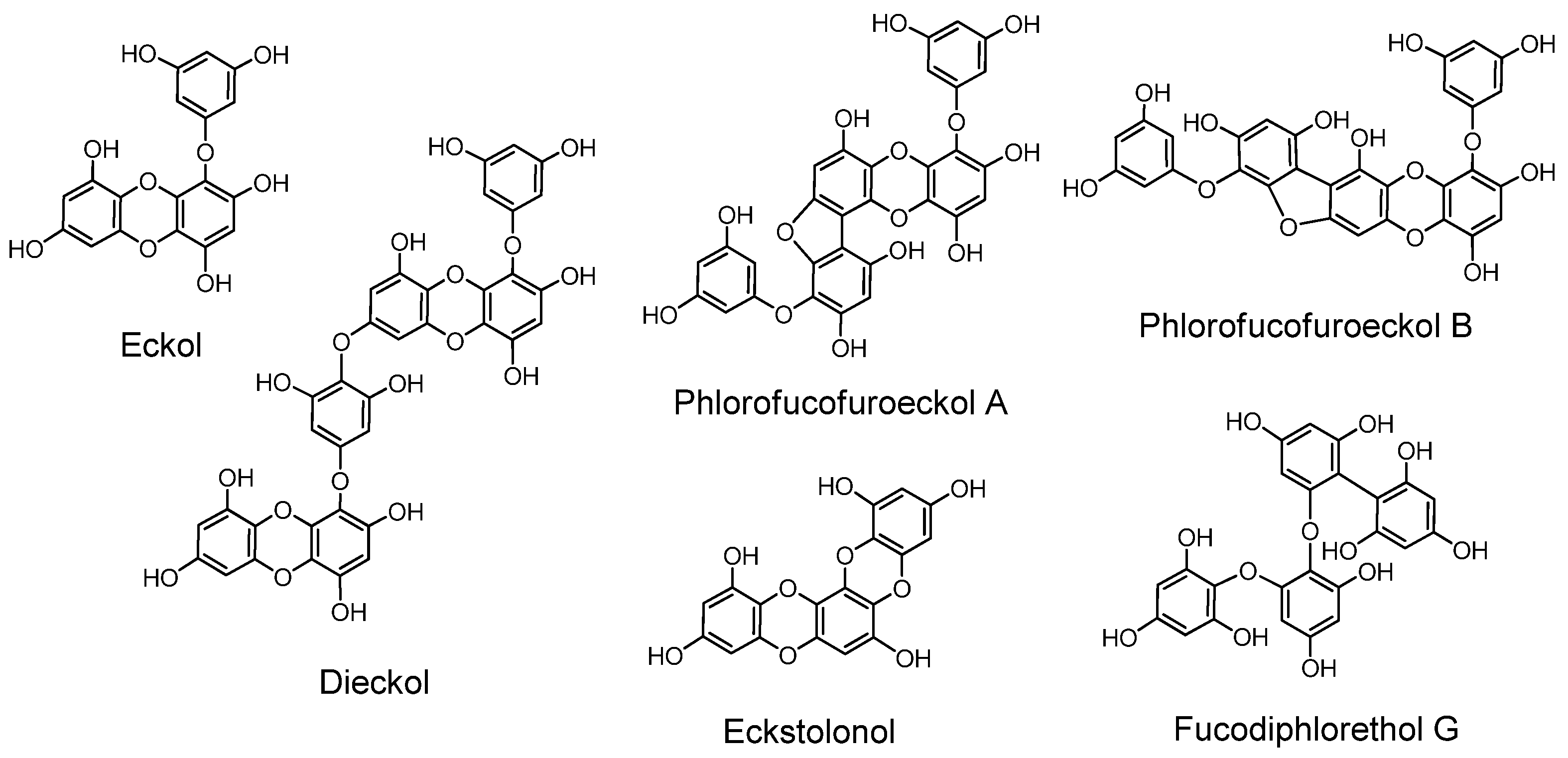
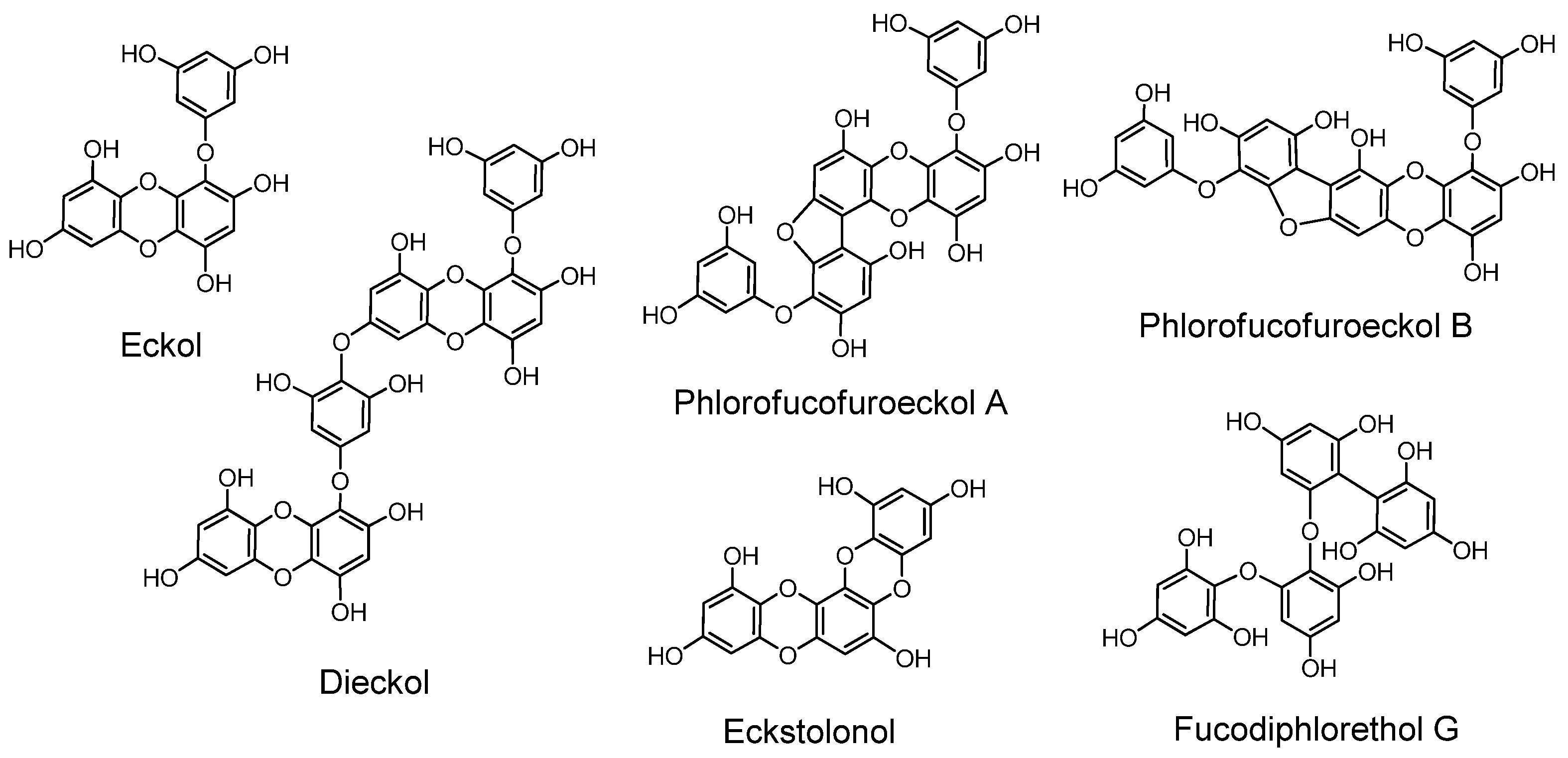
2.8. Phlorotannins (Figure 12)
3. Type B Tannins: Variable Mixtures of Analogous Polyphenols
3.1. Gallotannins of Type B
3.2. Polyhydroxyflavan Oligomers (Condensed Tannins)
3.2.1. Galloylated polyhydroxyflavan oligomers
3.2.2. Non-galloylated polyhydroxyflavans
4. Conclusions
References
- Haslam, E. Plant Polyphenols; Cambridge University Press: Cambridge, NY, USA, 1989. [Google Scholar]
- Okuda, T.; Mori, K; Hatano, T. Relationship of the structures of tannins to the binding activities with hemoglobin and methylene blue. Chem. Pharm. Bull. 1985, 33, 1424–1433. [Google Scholar] [CrossRef]
- Okuda, T. Systematics and health effects of chemically distinct tannins in medicinal plants. Phytochemistry 2005, 66, 2012–2031. [Google Scholar] [CrossRef]
- Okuda, T. Novel aspects of tannins—Renewed concept and structure-activity relationships. Curr. Org. Chem. 1999, 3, 609–622. [Google Scholar]
- Grassmann, W.; Endres, H.; Pauckner, W. Über die Gerbstoffe der Fichtenrinde, VI. Die Konstitution des Piceatannols. Chem. Ber. 1958, 91, 134–140. [Google Scholar] [CrossRef]
- Nakayama, Y.; Takahashi, M.; Fukuyama, Y.; Kinzyo, Z. An anti-plasmin inhibitor, eckol, isolated from the brown alga Echlonia kurome Okamura (biological chemistry). Agr. Biol. Chem. 1989, 53, 3025–3030. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Ito, H. Ellagitannins renewed the concept of tannins. In Chemistry and Biology of Ellagitannins; Quideau, S., Ed.; World Scientific: Singapore, 2009; pp. 1–54. [Google Scholar]
- Yoshida, T.; Hatano, T.; Ito, H.; Okuda, T. Structure diversity and antimicrobial activities of ellagitannins. In Chemistry and Biology of Ellagitannins; Quideau, S., Ed.; World Scientific: Singapore, 2009; pp. 55–93. [Google Scholar]
- Yoshida, T.; Mori, K.; Hatano, T.; Okumura, T.; Uehara, I.; Komagoe, K.; Fujita, Y.; Okuda, T. Radical-scavenging effects of tannins and related polyphenols on 1,1-diphenyl-2-picrylhydrazyl radical. Chem. Pharm. Bull. 1989, 37, 1919–1921. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Hydrolyzable tannins and related polyphenols. In Progress in the Chemistry of Organic Natural Products; Herz, G.W., Kirby, G.W., Moor, R.E., Steglich, W., Tamm, Ch., Eds.; Springer-Verlag: Vienna, Austria, 1995; Volume 66, pp. 1–117. [Google Scholar]
- Okuda, T. Structure-activity relationship of antioxidant and antitumor polyphenols. In Food Factors for Cancer Prevention; Ohigashi, H., Osawa, T., Terao, J., Watanabe, S., Yoshikawa, T., Eds.; Springer-Verlag: Vienna, Austria, 1997; pp. 280–285. [Google Scholar]
- Yoshizawa, S.; Horiuchi, T.; Fujiki, H.; Yoshida, T.; Okuda, T.; Sugimura, T. Antitumor promoting activity of (−)-Epigallocatechin gallate, the main constituent of “Tannin” in green tea. Phytother. Res. 1987, 1, 44. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Polyphenols from Asian plants, structural diversity and antitumor and antiviral activities. In Phenolic Compounds in Food and Their Effects on Health II, Antioxidants and Cancer Prevention; Huang, M.-T., Ho, C.-T., Lee, C.Y., Eds.; American Chemical Society: Washington DC, USA, 1992; pp. 160–183. [Google Scholar]
- Yoshizawa, S.; Horiuchi, T.; Suganuma, M.; Nishiwaki, S.; Yatsunami, J.; Okabe, S.; Okuda, T.; Muto, Y.; Frenkel, K.; Trol, W.; Fujiki, H. Penta-O-galloyl-β-D-glucose and (−)-Epigallocatechin gallate, cancer preventive agents. In ACS Symposium Series 507. Phenolic Compounds in Food and Their Effects on Health II: Antioxidants and Cancer Prevention; Huang, M.T., Ho, C.T., Lee, C.Y., Eds.; America Chemical Society: Washington DC, USA, 1992; p. 316. [Google Scholar]
- Quideau, S.; Feldman, K.S. Ellagitannin chemistry. Chem. Rev. 1996, 96, 475–503. [Google Scholar]
- Okuda, T.; Yoshida, T.; Hatano, T. Constituents of Geranium thunbergii Sieb. et Zucc. Part 12. Hydrated stereostructure and equibration of geraniin. J. Chem. Soc. Perkin Trans. 1 1982, 9–14. [Google Scholar]
- Okuda, T.; Yoshida, T.; Kuwahara, M.; Memon, M.U.; Shingu, T. Agrimoniin and potentillin, ellagitannin dimer and monomer having α-glucose core. J. Chem. Soc. Chem. Commun. 1982, 163–164. [Google Scholar]
- Takaoka, M. Resveratrol, a new phenolic compound from Veratrum grandiflorum. J. Chem. Soc. Jpn. 1939, 60, 1090–1100. [Google Scholar]
- Ferrigni, N.R.; McLaughlin, J.L.; Powell, R.G.; Smith, C.R., Jr. Use of potato disc and brine shrimp bioassays to detect activity and isolate piceatannol as the antileukemic principle from the seeds of Euphorbia lagascae. J. Nat. Prod. 1984, 47, 347–352. [Google Scholar] [CrossRef]
- Lee, S.H.; Park, M.H.; Heo, S.J.; Kang, S.M.; Ko, S.C.; Hans, J.S.; Jeon, Y.J. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef]
- Armitage, R.; Bayliss, G.S.; Gramshaw, J.W.; Haslam, E. Gallotannins part III: The constitution of Chinese, Turkish, Sumach, and Tara tannins. J. Chem. Soc. 1961, 1842–1853. [Google Scholar]
- Mayer, W.; Kunz, N.; Loebich, F. Die struktur hamamelitannins. Liebigs Ann. Chem. 1965, 688, 232–238. [Google Scholar] [CrossRef]
- Perkin, A.G.; Uyeda, Y. Occurrence of a crystalline tannin in the leaves of the Acer ginnala. J. Chem. Soc. 1922, 66–76. [Google Scholar]
- Haddock, E.A.; Gupta, R.K.; Al-Shafi, S.M.K.; Haslam, E.; Magnolato, D. The metabolism of gallic acid and hexahydroxydiphenic acid in plants. Part1: Introduction, naturally occurring galloyl esters. J. Chem. Soc. Perkin Trans. 1 1982, 2515–2524. [Google Scholar]
- Hatano, T.; Hattori, S.; Ikeda, Y.; Shingu, T.; Okuda, T. Gallotannins having 1,5-anhydro-D-glucitol core and some ellagitannins from Acer Species. Chem. Pharm. Bull. 1990, 38, 1902–1905. [Google Scholar] [CrossRef]
- Ozawa, T.; Kobayashi, S.; Seki, R.; Imagawa, H. A new gallotannin from bark of chestnut tree, Castanea crenata Sieb. et Zucc. Agric. Biol. Chem. 1984, 48, 1411–1416. [Google Scholar] [CrossRef]
- Nonaka, G.; Ishimaru, K.; Tanaka, T.; Nishioka, I. Galloylhamameloses from Castanea crenata and Sanguisorba officinalis. I. Chem. Pharm. Bull. 1984, 32, 483–489. [Google Scholar] [CrossRef]
- Yoshida, T.; Fujii, R.; Okuda, T. Revised structures of chebulinic acid and chebulagic acid. Chem. Pharm. Bull. 1980, 28, 3713–3715. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Correlation of oxidative transformations of hydrolyzable tannins and plant evolution. Phytochemistry 2000, 55, 513–529. [Google Scholar] [CrossRef]
- Yoshida, T.; Hatano, T.; Ito, H.; Okuda, T. Highly oxidized ellagitannins and their biological activity. In Plant Polyphenols 2—Chemistry, Biology, Pharmacology, Ecology; Gross, G.G., Hemingway, R.W., Yoshida, T., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 1999; pp. 127–144. [Google Scholar]
- Luger, P.; Weber, M.; Kashino, S.; Amakura, Y.; Yoshida, T.; Okuda, T.; Beurskens, G.; Dauter, Z. Structure of the Tannin Geraniin based on Conventional X-ray Data at 295K and on Synchrotron Data at 293 and 120K. Acta Cryst. 1998, B54, 687–694. [Google Scholar]
- Quideau, S.; Feldman, K.S. Ellagitannin chemistry. The first synthesis of dehydrohexahydroxydiphenoate esters from oxidative coupling of unetherified methyl gallate. J. Org. Chem. 1997, 62, 8809–8813. [Google Scholar] [CrossRef]
- Okuda, T.; Mori, K.; Hatano, T. The distribution of geraniin and mallotusinic acid in the order geraniales. Phytochemistry 1980, 19, 547–551. [Google Scholar] [CrossRef]
- Okuda, T.; Mori, K.; Hayatsu, H. Inhibitory effect of tannins on direct-acting mutagens. Chem. Pharm. Bull. 1984, 32, 3755–3758. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Ashida, M.; Yazaki, K. Structures of pedunculagin, casuarictin, strictinin, casuarinin, casuariin and stachyurin. J. Chem. Soc. Perkin Trans. 1 1983, 1765–1772. [Google Scholar]
- Nakanishi, Y.; Okuda, T.; Abe, H. Effects of geraniin on the liver in rats. III. Nat. Med. 1999, 53, 22–26. [Google Scholar]
- Kimura, Y.; Okuda, H.; Mori, K.; Okuda, T.; Arichi, S. Effects of various extracts of Geranii Herba and geraniin on liver injury and lipid metabolism in rats fed peroxidized oil. Chem. Pharm. Bull. 1984, 32, 1866–1871. [Google Scholar] [CrossRef]
- Kimura, Y.; Okuda, H.; Okuda, T.; Arichi, S. Effects of geraniin, corilagin and ellagic acid isolated from Geranii herba on arachidonate metabolism in leukocytes. Planta Med. 1986, 52, 337–338. [Google Scholar] [CrossRef]
- Yamada, H.; Nagao, K.; Dokei, K.; Kasai, Y.; Michihata, N. Total synthesis of (−)-Corilagin. J. Am. Chem. Soc. 2008, 130, 7566–7567. [Google Scholar]
- Okuda, T.; Yoshida, T.; Hatano, T.; Koga, T.; Toh, N.; Kuriyama, K. Circular dichroism of hydrolyzable tannins, I. Tetrahedron Lett. 1082, 23, 3937–3940. [Google Scholar]
- Wilkins, C.K.; Bohm, B.A. Ellagitannins from Tellima grandiflora. Phytochemistry 1976, 15, 211–214. [Google Scholar] [CrossRef]
- Yazaki, K.; Hatano, T.; Okuda, T. Structures of dehydrogeraniin, furosinin and furosin. J. Chem. Soc. Perkin Trans.1 1989, 2289–2296. [Google Scholar]
- Schmidt, O.T.; Mayer, W. Naturliche gerbstoffe. Angew. Chem. 1956, 68, 103–115. [Google Scholar] [CrossRef]
- Uddin, M.; Haslam, E. Some observations on the structure of chebulinic acid and its derivatives. J. Chem. Soc. (C) 1967, 2381–2384. [Google Scholar]
- Yoshida, T.; Itoh, H.; Matsunaga, S.; Tanaka, R.; Okuda, T. Hydrolyzable tannins with 1C4 glucose core from Phyllanthus flexuosus muell. Arg. Chem. Pharm. Bull. 1992, 40, 53–60. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Ikeda, Y.; Shingu, T.; Inoue, T. Isolation of water-soluble tannins by centrifugal partition chromatography, and biomimetic synthesis of elaeocarpusin. Chem. Pharm. Bull. 1986, 34, 4075–4082. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T. Biomimetic synthesis of elaeocarpusin. Heterocycles 1986, 24, 1841–1843. [Google Scholar] [CrossRef]
- Amakura, Y.; Yoshida, T. Tannins and related polyphenols of euphorbiaceous plants. XIV. Euphorbin I, a new dimeric hydrolyzable tannin from Euphorbia watanabei. Chem. Pharm. Bull. 1996, 44, 1293–1297. [Google Scholar] [CrossRef]
- Foo, L.Y.; Wong, H. Phyllanthusiin D, an unusual hydrolysable tannin from Phyllanthus amarus. Phytochemistry 1992, 31, 711–713. [Google Scholar]
- Yazaki, K.; Yoshida, T.; Okuda, T. Tannin production in cell suspension culture of Geranium thunbergii. Phytochemisrty 1991, 30, 501–503. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Nayeshiro, H. Constituents of Geranium thunbergii SIEB. et Zucc. IV. Ellagitannins. (2). Structure of Geraniin. Chem. Pharm. Bull. 1977, 25, 1862–1869. [Google Scholar] [CrossRef]
- Okuda, T.; Hatano, T.; Nitta, H.; Fujii, R. Hydrolyzable tannins having enantiomeric dehydrohexahydroxydiphenoyl group: Revised structure of terchebin and structure of granatin B. Tetrahedron Lett. 1980, 21, 4361–4364. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Nayeshiro, H. Structure of geraniin. Chem. Pharm. Bull. 1977, 25, 1862–1869. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Kuwahara, M.; Memon, M.U.; Shingu, T. Agrimoniin and potentillin, an ellagitannin dimer and monomer having an α-glucose core. J. Chem. Soc. Chem. Commun. 1982, 162–164. [Google Scholar]
- Miyamoto, K.; Kishi, N.; Koshiura, R.; Yoshida, T.; Hatano, T.; Okuda, T. Relationship between the structures and the antitumor activities of tannins. Chem. Pharm. Bull. 1987, 35, 814–822. [Google Scholar] [CrossRef]
- Okuda, T.; Yoshida, T.; Hatano, T.; Kubo, M.; Orime, T.; Yoshizaki, M.; Naruhashi, N. Hydrolysable tannins as chemotaxonomic markers in the rosaceae. Phytochemistry 1992, 31, 3091–3096. [Google Scholar] [CrossRef]
- Yoshida, T.; Maruyama, Y.; Memon, M.U.; Shingu, T.; Okuda, T. Gemins A, B and C, new dimeric ellagitannins from Geum japonicum. J. Chem. Soc. Perkin Trans. 1 1985, 315–321. [Google Scholar]
- Hatano, T.; Hattori, S.; Okuda, T. Tannins of Coriaria japonica A. Gray, I: Coriariin A and B, new dimeric and monomeric hydrolyzable tannins. Chem. Pharm. Bull. 1986, 34, 4092–4097. [Google Scholar] [CrossRef]
- Yoshida, T.; Ito, H.; Hippolito, I.J. Pentameric ellagitannin oligomers in melasmataceous plants—chemotaxonomic significance. Phytochemistry 2005, 66, 1972–1983. [Google Scholar] [CrossRef]
- Karonen, M.; Salminen, J.-P. First evidence of hexa- and heptameric ellagitannins in plants. In Polyphenol Communications, 2010 ed. Ageorges, A., Cheynier, V., Lefer, P., Sarni-Manchado, P., Eds.; XXVth International Conference on Polyphenols: Montpellier, France, 2010; pp. 540–541. [Google Scholar]
- Hatano, T.; Yasuhara, T.; Matsuda, M.; Yazaki, K.; Yoshida, T.; Okuda, T. Oenothein B, a dimereric hydrolysable tannin with macrocyclic structure and accompanying tannins from Oenothera erythrosepara. J. Chem. Soc. Perkin Trans. 1 1990, 2735–2743. [Google Scholar]
- Okuda, T.; Yoshida, T.; Hatano, T.; Yazaki, K.; Kira, R.; Ikeda, Y. Preparative fractination of hydrolyzable tannins by centrifugal partition chromatography. J. Chromatogr. 1986, 362, 375–381. [Google Scholar] [CrossRef]
- Yoshida, T.; Chou, T.; Nitta, A.; Miyamoto, K.; Koshiura, R.; Okuda, T. Woodfordin C, a macro-ring hydrolyzable tannin dimer with antitumor activity and accompanying dimers from Woodfordia fruticosa flowers. Chem. Pharm. Bull. 1990, 38, 1211–1217. [Google Scholar] [CrossRef]
- Miyamoto, K.; Sasakura, M.; Matsui, E.; Koshiura, R.; Murayama, T.; Hatano, T.; Yoshida, T.; Okuda, T. Antitumor activity of oenothein B, a unique macrocyclic ellagitannin. Jpn. J. Cancer Res. 1993, 84, 99–103. [Google Scholar] [CrossRef]
- Yoshida, T.; Chou, T.; Matsuda, M.; Yasuhara, T.; Yazaki, K.; Hatano, T.; Nitta, A.; Okuda, T. Woodfordin D and oenothein A, trimeric hydrolyzable tannins of macro-ring structure with antitumor activity. Chem. Pharm. Bull. 1991, 39, 1157–1162, (Japan). [Google Scholar] [CrossRef]
- Hervé du Penhoat, C.L.M.; Michon, V.M.F.; Ohassan, A.; Peng, S.; Scalbert, A.; Gage, D. Structural elucidation of new dimeric ellagitannins from Quercus robur. Phytochemistry 1991, 30, 329–332. [Google Scholar]
- Vivas, N.; Glories, Y.; Bourgeois, G.; Vitry, C. The heartwood ellagitannins of different oak (Quercus sp.) and chestnut species (Castanea sativa Mill.). Quantity analysis of red wines aging in Barels. J. Coop. Sci. Tech. 1996, 2, 51–75. [Google Scholar]
- Jourdes, M.; Saucier, C.; Quideau, S.; Teissedre, P.-L. Identification, amount and kinetics of extraction of C-Glycosidic ellagitannins during wine aging in Oak Barrels or in stainless steel vats with Oak Chips. In Polyphenol Comminications, 2010 edition; Ageorges, A., Cheynier, V., Lefer, P., Sarni-Manchado, P., Eds.; XXVth International Conference on Polyphenols: Montpellier, France, 2010; pp. 134–135. [Google Scholar]
- Hatano, T.; Shida, S.; Han, L.; surname, T. Two new complex tannins from Camellia japonica L. Chem. Pharm. Bull. 1991, 39, 876–880. [Google Scholar] [CrossRef]
- Yoshida, T.; Nakata, F.; Hosotani, K.; Nitta, A.; Okuda, T. Three new complex tannins from Melastoma malabathricum L. Chem. Pharm. Bull. 1992, 40, 1727–1732. [Google Scholar] [CrossRef]
- Hatano, T.; Kira, R.; Yoshizaki, M.; Okuda, T. Seasonal changes in the tannins of Liquidambar formosana reflectin their biogenesis. Phytochemisrty 1986, 25, 2787–2789. [Google Scholar] [CrossRef]
- Okuda, T.; Mori, K.; Ishino, M. Transformations of geraniin upon decoction. Yakugaku Zasshi 1979, 99, 505–509. [Google Scholar]
- Das, M.; Bickers, D.R.; Mukhtar, H. Effect of ellagic acid on hepatic and pulmonary xenobiotic metabolism in mice: Studies on mechanism of its anticarcinogenic action. Carcinogenesis 1985, 6, 1409–1413. [Google Scholar] [CrossRef]
- Lesca, P. Protective effects of ellagic acid and other plant phenols on benzo[a]pyrene-induced neoplasia in mice. Carcinogenesis 1983, 4, 1651–1653. [Google Scholar] [CrossRef]
- Ito, H.; Iguchi, A.; Hatano, T. Identification of urinary and intestinal bacterial metabolites of ellagitannin geraniin in rats. J. Agric. Food Chem. 2008, 56, 393–400. [Google Scholar] [CrossRef]
- Ishimoto, H.; Ito, H.; Kimura, Y.; Yoshimura, M.; Amakura, Y.; Yoshida, T.; Tai, A.; Hatano, T. Antioxidant properties of tannin metabolites in biofluids. In Pacifichem, 2010 ed.; Honolulu: Hawaii, USA, 2010. [Google Scholar]
- Fujita, Y.; Komagoe, K.; Niwa, Y.; Uehara, I.; Hara, R.; Mori, H.; Okuda, T.; Yoshida, T. Inhibition mechanism of tannins isolated from medicinal plants and related compounds on autoxidation of methyl linoleate. Yakugaku Zasshi 1988, 108, 528–537. [Google Scholar]
- Yoshizawa, S.; Horiuchi, T.; Fujiki, H.; Yoshida, T.; Okuda, T.; Sugimura, T. Antumor promoting activity of (−)-epigallocatechin gallate, the main constituent of “tannin” in green tea. Phytother. Res. 1987, 1, 44–47. [Google Scholar]
- Kuzuhara, T.; Suganuma, M.; Fujiki, H. Green tea catechin as a chemical chaperone in cancer prevention. Cancer Lett. 2008, 261, 12–20. [Google Scholar] [CrossRef]
- Shimizu, M.; Fukutomi, Y.; Ninomiya, M.; Nagura, K.; Kato, T.; Araki, H.; Suganuma, M.; Fujiki, H.; Moriwaki, H. Green tea extracts for the prevention of metachronous colorectal adenomas: A pilot study. Cancer Epidem. Biomarker. Prev. 2008, 17, 3020–3025. [Google Scholar] [CrossRef]
- Hatano, T.; Tsugawa, M.; Ohyabu, T.; Kusuda, M.; Shiota, S.; Tsuchiya, T.; Yoshida, T. Effects of polyphenols in tea and foods on methicillin-resistant Staphylococcus aureus and the sustainability of the antibacterial effects in the presence of food additives. J. Jpn. Soc. Med. Use Funct. Foods 2006, 4, 43–48. [Google Scholar]
- Shiota, S.; Shimizu, M.; Sugiyama, J.; Morita, Y.; Mizushima, T.; Tsuchiya, T. Mechanisms of action of corilagin and tellimagrandin i that remarkably potentiate the activity of β-lactams against methicillin-resistant Staphilococcus aureus. Microbiol. Immunol. 2004, 48, 67–73. [Google Scholar]
- Shiota, S.; Shimizu, M.; Mizushima, T.; Ito, H.; Hatano, T.; Yoshida, T.; Tsuchiya, T. Restoration of effectiveness of beta-lactams on methicillin-resistant Staphylococcus aureus by tellimagrandin I from rose red. FEMS Microbiol. Lett. 2000, 185, 135–138. [Google Scholar]
- Asanaka, M.; Kurimura, T.; Koshiura, R.; Okuda, T.; Mori, M.; Yokoi, H. Inhibitory effect of ellagitannins on the in vitro replication of human immuno-deficiency virus (HIV). AIDS Res. Newslett. 1987, 1, 72. [Google Scholar]
- Ito, H.; Miyake, M.; Nishitani, E.; Mori, K.; Hatano, T.; Okuda, T.; Konoshima, T.; Takasaki, M.; Kozuka, M.; Mukainaka, T.; Tokuda, H.; Nishino, H.; Yoshida, T. Anti-tumor promoting activity of polyphenols from Cowania mexicana and Coleogyne ramosissima. Cancer Lett. 1999, 143, 5–12. [Google Scholar] [CrossRef]
- Miyamoto, K.; Murayama, T.; Yoshida, T.; Hatano, T.; Okuda, T. Anticarcinogenic activities of polyphenols in foods and herbs. In Antinutrients and Phytochemicals in Food; Shahidi, F., Ed.; ACS: Washington, DC, USA, 1997; pp. 245–259. [Google Scholar]
- Miyamoto, K.; Murayama, T.; Nomura, M.; Hatano, T.; Furuya, T.; Koshiura, R.; Okuda, T. Antitumor activity and interleukin-1 induction by tannins. Anticancer Res. 1993, 31, 37–42. [Google Scholar]
- Hegnauer, R. Chemotaxonomie der Pflanzen, 4th ed; Birkhaeuser: Basel-Stuttgart, Switzerland, 1966; p. 327. [Google Scholar]
- Okuda, T.; Hatano, T.; Agata, I.; Nishibe, S.; Kimura, K. Tannins in Artemisia Montana, A. princeps and related species of plant. Yakugaku Zasshi 1986, 106, 894–899. [Google Scholar]
- Okuda, T.; Hatano, T.; Agata, I.; Nishibe, S. The comnponents of tannic activities in labiatae plants, I: rosmarinic acid from labiatae plants in Japan. Yakugaku Zasshi 1986, 106, 1108–1111. [Google Scholar]
- Kelly, C.J.; Harruff, R.C.; Carmack, M. The polyphenolic acids of Lithospermum ruderale, II. carbon-13 nuclear magnetic resonance of lithospermic and rosmarinic acids. J. Org. Chem. 1976, 41, 449–455. [Google Scholar] [CrossRef]
- Agata, I.; Kusakabe, H.; Hatano, T.; Nishibe, S.; Okuda, T. Melitric acids A and B, new trimeric caffeic acid derivatives from Melissa officinalis. Chem. Pharm. Bull. 1993, 41, 1608–1611. [Google Scholar] [CrossRef]
- Agata, I.; Hatano, T.; Nishibe, S.; Okuda, T. A tetrameric derivative of caffeic acid from Rabdosia japonica. Phytochemistry 1989, 28, 2447–2450. [Google Scholar] [CrossRef]
- Ito, T.; Abe, N.; Ali, Z.; Oyama, M.; Tanaka, T.; Sawa, R.; Takahashi, Y.; Murata, J.; Darnaedi, D.; Iinuma, M. Two new resveratrol tetramers from Upuna borneensis. Chem. Pharm. Bull. 2009, 57, 516–519. [Google Scholar] [CrossRef]
- Ito, T.; Abe, N.; Ali, Z.; Oyama, M.; Tanaka, T.; Murata, J.; Danaedi, D.; Iinuma, M. Resveratrol tetramer with a C6-C3 or a C1 unit from Upuna borneensis. Chem. Pharm. Bull. 2007, 55, 1535–1539. [Google Scholar] [CrossRef]
- Abe, N.; Ito, T.; Oyama, M.; Iinuma, M. Oligomers from Vatica chinensis. In The 5th JSP-CCTNM-KSP Symposium on Pharmacognosy, Tokushima, Japan, September 2010; p. 350, Abstract Papers.
- Xiao, K.; Zhang, H.-J.; Xuan, L.-J.; Zhang, J.; Xu, Y.-M.; Bai, D.-L. Stilbenoids: Chemistry and bioactivities. Stud. Nat. Prod. Chem. 2008, 34, 453–646. [Google Scholar] [CrossRef]
- Gorham, J.; Tori, M.; Asakawa, Y. The Biochemistry of the Stilbenoids; Chapman & Hall: London, UK, 1995. [Google Scholar]
- Shu, N.; Hong, Z.; Changgi, H. Simultaneous dtermination of the contents of three stilbene oligomers in Caragana sinica collected in different seasons using an improved HPLC method. Biol. Pharm. Bull. 2006, 29, 608–612. [Google Scholar] [CrossRef]
- Siemann, E.H.; Creasey, L.L. Concentration of the phytoalexin resveratrol in wine. Am. J. Enol. Viticilt. 1992, 43, 49–52. [Google Scholar]
- Rimando, A.M.; Kalt, W.; Magee, J.B.; Dewey, J.; Ballington, J.R. Resveratrol, pterostilbene, and piceatannol in vaccinium berries. J. Agric. Food Chem. 2004, 52, 4713–4719. [Google Scholar] [CrossRef]
- Woo, A.; Min, B.; Ryoo, S. Piceatannol-3’-O-β-D-glucopyranoside as an active component of rhubarb activates endothelial nitric oxide synthase through inhibition of arginase activity. Exp. Mol. Med. 2010, 42, 524–532. [Google Scholar] [CrossRef]
- Miura, T.; Muraoka, S.; Ikeda, N.; Watanabe, M.; Fujimoto, Y. Antioxidative and prooxidative action of stilbene derivatives. Basic Clin. Pharmacol. Toxicol. 2000, 86, 203–208. [Google Scholar]
- Cunningham, J.; Haslam, E.; Haworth, R.D. The constitution of piceatannol. J. Chem. Soc. 1963, 2875–2883. [Google Scholar]
- Duarte, N.; Kayser, O.; Abreu, P.; Ferreira, M. J. Antileishmanial activity of piceatannol isolated from Euphorbia lagascae seeds. Phytother. Res. 2008, 22, 455–457. [Google Scholar] [CrossRef]
- Yokozawa, T.; Kim, Y.J. Piceatannol inhibits melanogemesis by its antioxidative actions (Biochemistry). Chem. Pharm. Bull. 2007, 30, 2007–2011. [Google Scholar] [CrossRef]
- Larrosa, M.; Thomás-Barberán, F.A.; Espín, J.C. The grape and wine polyphenol piceatannol is a potent inducer of apoptosis in human SK-Mel-28 melanoma cells. Eur. J. Nutr. 2004, 43, 275–284. [Google Scholar] [CrossRef]
- Geahlen, R.L.; McLaughlin, J.L. Piceatannol (3,4,3’,5’-tetrahydroxy-trans-stilbene) is a naturally ocurring protein-tyrosine kinase inhibitor. Biochem. Biophys. Res. Comm. 1989, 165, 241–245. [Google Scholar] [CrossRef]
- Swanson-Mungerson, M.; Ikeda, M.; Lev, L.; Longneckuy, R.; Portis, T. Identification of latent membrane protein 2A (LMP2A) specific targets for treatment and eradication of epstein-barr virus (EBV)-associated diseases. J. Antimicrob. Chemother. 2003, 52, 152–154. [Google Scholar] [CrossRef]
- Chen, J.; Ma, M.; Lu, Y.; Wang, L.; Wu, C.; Duan, H. Rhaponticin from rhubarb rhizomes alleviates liver steatosis and improves blood glucose and lipid profiles in KK/Ay diabetic mice. Planta Med. 2009, 75, 472–477. [Google Scholar] [CrossRef]
- Iinuma, M.; Ohyama, M.; Tanaka, T.; Lang, F.A. Flavonostilbene and two stilbene oligomers in roots of Sophora leachiana. Phytochemistry 1994, 37, 1157–1159. [Google Scholar] [CrossRef]
- Wada, S.; Yasui, Y.; Hitomi, T.; Tanaka, R. Flavonostilbenes jezonocinols A, B and C with DPPH radical scavenging activity were isolated from Picea jezoensis var. jezoensis. J. Nat. Prod. 2007, 70, 1605–1610. [Google Scholar] [CrossRef]
- Ito, T.; Abe, N.; Oyama, M.; Iinuma, M. Oligostilbenoids from dipterocarpaceaeous plants. A new resveratrol tetramer from Vatica indica and the revised structure of isohopeaphenol. Helv. Chim. Acta 2008, 91, 1989–1998. [Google Scholar] [CrossRef]
- Xiang, T.; Uno, T.; Ogino, F.; Ai, C.; Duo, J.; Sankawa, U. Antioxidant constituents of Caragana tibetica. Chem. Pharm. Bull. 2005, 53, 1204–1206. [Google Scholar] [CrossRef]
- Privat, C.; Telo, J.P.; Bernardes-Genisson, V.; Vieira, A.; Souchard, J.-P.; Nepveu, F. Antioxidant properties of trans-ε-viniferin as compared to stilbene derivatives in aqueous and nonaqueous media. J. Agric. Food Chem. 2002, 50, 1213–1217. [Google Scholar]
- Kitanaka, S.; Ikezawa, T.; Yasukawa, K.; Yamanouchi, S.; Takido, M.; Sung, H.K.; Kim, I.H. (+)-α-Viniferin, an anti-inflammatory compound from Caragana chamlagu root. Chem. Pharm. Bull. 1990, 38, 432–435. [Google Scholar] [CrossRef]
- 117 Abe, N.; Ito, T.; Ohguchi, K.; Nasu, M.; Masuda, Y.; Oyama, M.; Nozawa, Y.; Ito, M.; Iinuma, M. Resveratrol oligomers from Vatica albiramis. J. Nat. Prod. 2010, 73, 1499–1506. [Google Scholar]
- Tanaka, T.; Ito, T.; Nakaya, K.; Iinuma, M.; Takahashi, Y.; Naganawa, H.; Matsuura, N.; Ubukata, M. Vaticanol D a novel resveratrol hexamer isolated from Vatica rassak. Tetrahedron Lett. 2000, 41, 7929–7932. [Google Scholar] [CrossRef]
- Ito, T.; Tanaka, T.; Ali, Z.; Akao, Y.; Takahashi, Y.; Sawa, R.; Nakaya, K.-I.; Murata, J.; Darnaedi, D.; Iinuma, M. A new resveratrol hexamer from Upuna borneensis. Heterocycles 2004, 63, 129–136. [Google Scholar] [CrossRef]
- Ito, T.; Tanaka, T.; Iinuma, M.; Nakaya, K.-I.; Takahashi, Y.; Sawa, R.; Murata, J.; Darnaedi, D. Two new resveratrol hexamer (5-[1E]-2-(4-Hydroxyphenyl) ethenyl]-benzene-1,3-diol) tetramers with a tetraheydrofuran ring from Dipterocarpus grandiflorus. Helv. Chim. Acta 2004, 87, 479–495. [Google Scholar]
- Ito, T.; Tanaka, T.; Nakaya, K.; Iinuma, M.; Takahashi, Y.; Bastow, K.F.; Lee, K.-H. A new resveratrol octamer, vateriaphenol A, in Vatica indica. Tetrahedron Lett. 2001, 42, 5909–5912. [Google Scholar] [CrossRef]
- Sakagami, Y.; Sawabe, A.; Komemushi, S.; Ali, Z.; Tanaka, T.; Iliya, I.; Iinuma, M. Antibacterial activity of stilbene oligomers against vancomycin-resistant Enterococci (VRE) and methicillin-resistant Staphylococcus aureus (MRSA) and their synergism with antibiotics. Biocontrol Sci. 2007, 12, 7–14. [Google Scholar] [CrossRef]
- Yamada, M.; Hayashi, K.; Ikeda, S.; Tsutsui, Ke.; Tsutsui, Ki.; Ito, T.; Iinuma, M.; Nazaki, H. Inhibitory activity of plant stilbene oligomers against DNA topoisomerase II (pharmacogosy). Biol. Pharm. Bull. 2006, 29, 1504–1507, (Japan). [Google Scholar]
- Ito, T.; Akao, Y.; Yi, H.; Ohguchi, K.; Matsumoto, K.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Antitumor effect of resveratrol oligomers against human cancer cell lines and the molecular mechanism of apoptosis induced by vitamin C. Carcinogenesis 2003, 24, 1489–1497. [Google Scholar]
- Iliya, I.; Akao, Y.; Matsumoto, K.; Nakagawa, Y.; Zulfigar, A.; Ito, T.; Oyama, M.; Murata, H.; Tanaka, T.; Nozawa, Y.; Iinimu, M. Growth inhibition of stilbenoids in welwitsiaceae and gnetaceae through induction of apoptosis in human leukemia HL60 Cells. Biol. Pharm. Bull. 2006, 29, 1490–1492. [Google Scholar] [CrossRef]
- Morikawa, T.; Chaipech, S.; Matsuda, H.; Asao, Y.; Hamao, M.; Ninomiya, K.; Muraoka, O.; Pongpiriyadacha, Y.; Hayakawa, T.; Yoshikawa, M. Oligostilbenoids with anti-hyperlipidemic activity from thai natural medicine phayom, the bark of Shorea roxburghii. In The 5th JSP-CCTNM-KSP Joint Symposium on Pharmacognosy, Tokushima, Japan, 24 September 2010; p. 55, Abstract Papers.
- Grosse-Damhues, J.; Glombitza, K.-W.; Schulten, H.-R. An eight-ring phlorotannin from the brown alga Himanthalia elongate. Phytochemistry 1983, 22, 2043–2046. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Kodama, M.; Miura, I.; Kinjo, Z. Mori, H.; Nakayama, Y.; Takahashi, M. Anti-plasmin inhibitor VI: Structure of phlorofucofuroeckol A, a novel phlorotannin with both dibenzo-1,4-dioxin and dibenzofuran elements, from Ecklonia kurome Okamura. Chem. Pharm. Bull. 1990, 38, 133–135. [Google Scholar] [CrossRef]
- Fukuyama, Y.; Miura, I.; Kinzyo, Z.; Mori, H.; Kido, M.; Nakayama, Y.; Takahashi, M.; Ochi, M. The phlorotannin with a dibenzo-p-dioxin skeleton, isolated from Ecklonia kurome, a brown alga, inhibited plasma α2” macroglobulin. Chem. Lett. 1985, 739–742. [Google Scholar]
- Sook, K.H.; Young, C.H.; Hyung, J.J.; Wha, S.B. A new phlorotannin, eckstolonol (5,8,13,14-tetraoxapentaphene-1,3.6,9.11-pentaol) from the brown alga Ecklonia stolonifera. Chem. Pharm. Bull. 2003, 51, 1012–1014. [Google Scholar] [CrossRef]
- Sugiura, Y.; Matsuda, K.; Yamada, Y.; Nishikawa, M.; Shioya, K.; Katsuzaki, H.; Imai, K.; Amano, H. Isolation of a new anti-allergic phlorotannin, phlorofucofuroeckol-B from an edible brown alga, Eisenia arborea. Biosci. Biotechnol. Biochem. 2006, 70, 2807–2811. [Google Scholar] [CrossRef]
- Young, M.H.; Jong, S.B.; Jim, W.H.; Nam, H.L. Isolation of a new phlorotannin, fucodiphlorethol G, from a brown alga Ecklonia cava. Bull. Korean Chem. Soc. 2007, 28, 1595–1597. [Google Scholar] [CrossRef]
- Moon, C.; Kim, S.-H.; Kim, J-C.; Hyun, J.W.; Lee, N.H.; Park, J.W.; Shin, T. Protective effect of phlorotannin components phloroglucinol and eckol on radication-induced intestinal injury in mice. Phytother. Res. 2008, 22, 238–242. [Google Scholar] [CrossRef]
- Glombitza, K.-W.; Hauperich, S. Acetylation or methylation of phlorotannins masks their biological activities. Phytochemistry 1997, 46, 735–740. [Google Scholar] [CrossRef]
- Shibata, T.; Kawaguchi, S.; Hama, Y.; Inagaki, M.; Yamaguchi, K.; Nakamura, T. Local and chemical distribution of phlorotannins in brown algae. J. Appl. Phycol. 2004, 16, 291–296. [Google Scholar] [CrossRef]
- Meinnes, A.G.; Ragan, M.A.; Smith, D.G.; Walter, J.A. High-molecular-weight phloroglucinol-based tannins from brown algae: Structural variants. Hydrobiologia 1984, 116-117, 597–602. [Google Scholar] [CrossRef]
- Amsler, C.D.; Fairhead, V.A. Defensive and sensory chemical ecology of brown algae. Advan. Botan. Res. 2005, 43, 1–91. [Google Scholar] [CrossRef]
- Niemetz, R.; Gross, G.G. Enzymology of gallotannin and ellagitannin biosynthesis. Phytochemistry 2005, 66, 2001–2011. [Google Scholar] [CrossRef]
- Li, Y.; Shibahara, A.; Matsuo, Y.; Tanaka, T.; Kouno, I. Reaction of the black tea pigments theaflavin during enzymatic oxidation of tea catechins. J. Nat. Prod. 2010, 73, 33–39. [Google Scholar] [CrossRef]
- Matsuo, T.; Itoo, S. The chemical structure of kaki-tannin from immature fruit of the persimmon (Diospyros kaki L.). Agric. Biol. Chem. 1978, 42, 1637–1643. [Google Scholar] [CrossRef]
- Ferreira, D.; Marais Jannie, P.J.; Desmond, S. Heterogeneity of the Interflavanyl bond in proanthocyanidins from natural sources lacking C-4(C-ring) deoxyflavonoid nucleophiles. Phytochemistry 2005, 66, 2216–2237. [Google Scholar] [CrossRef]
- Ferreira, D. Proanthocyanidin/polyphenol research: Trial and thrills. Planta Medica 2010, 76, 1164. [Google Scholar]
- Hatano, T.; Edamatsu, R.; Hiramatsu, M.; Mori, A.; Fujita, Y.; Yasuhara, T.; Yoshida, T.; Okuda, T. Effects of tannins and related polyphenols on superoxide anion radical, and on 1,1-diphenyl-2-picrylhydrazyl Radical. Chem. Pharm. Bull. 1989, 37, 2016–2021. [Google Scholar] [CrossRef]
- Hatano, T.; Urita, K.; Okuda, T. Tannins in Saxifraga stolonifera. J. Med. Pharm. Soc. WAKAN-YAKU 1986, 3, 434–435. [Google Scholar]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Okuda, T.; Ito, H. Tannins of Constant Structure in Medicinal and Food Plants—Hydrolyzable Tannins and Polyphenols Related to Tannins. Molecules 2011, 16, 2191-2217. https://doi.org/10.3390/molecules16032191
Okuda T, Ito H. Tannins of Constant Structure in Medicinal and Food Plants—Hydrolyzable Tannins and Polyphenols Related to Tannins. Molecules. 2011; 16(3):2191-2217. https://doi.org/10.3390/molecules16032191
Chicago/Turabian StyleOkuda, Takuo, and Hideyuki Ito. 2011. "Tannins of Constant Structure in Medicinal and Food Plants—Hydrolyzable Tannins and Polyphenols Related to Tannins" Molecules 16, no. 3: 2191-2217. https://doi.org/10.3390/molecules16032191