Studies on the Genetic Variation of the Green Unicellular Alga Haematococcus pluvialis (Chlorophyceae) Obtained from Different Geographical Locations Using ISSR and RAPD Molecular Marker

Abstract
:1. Introduction
2. Results and Discussion



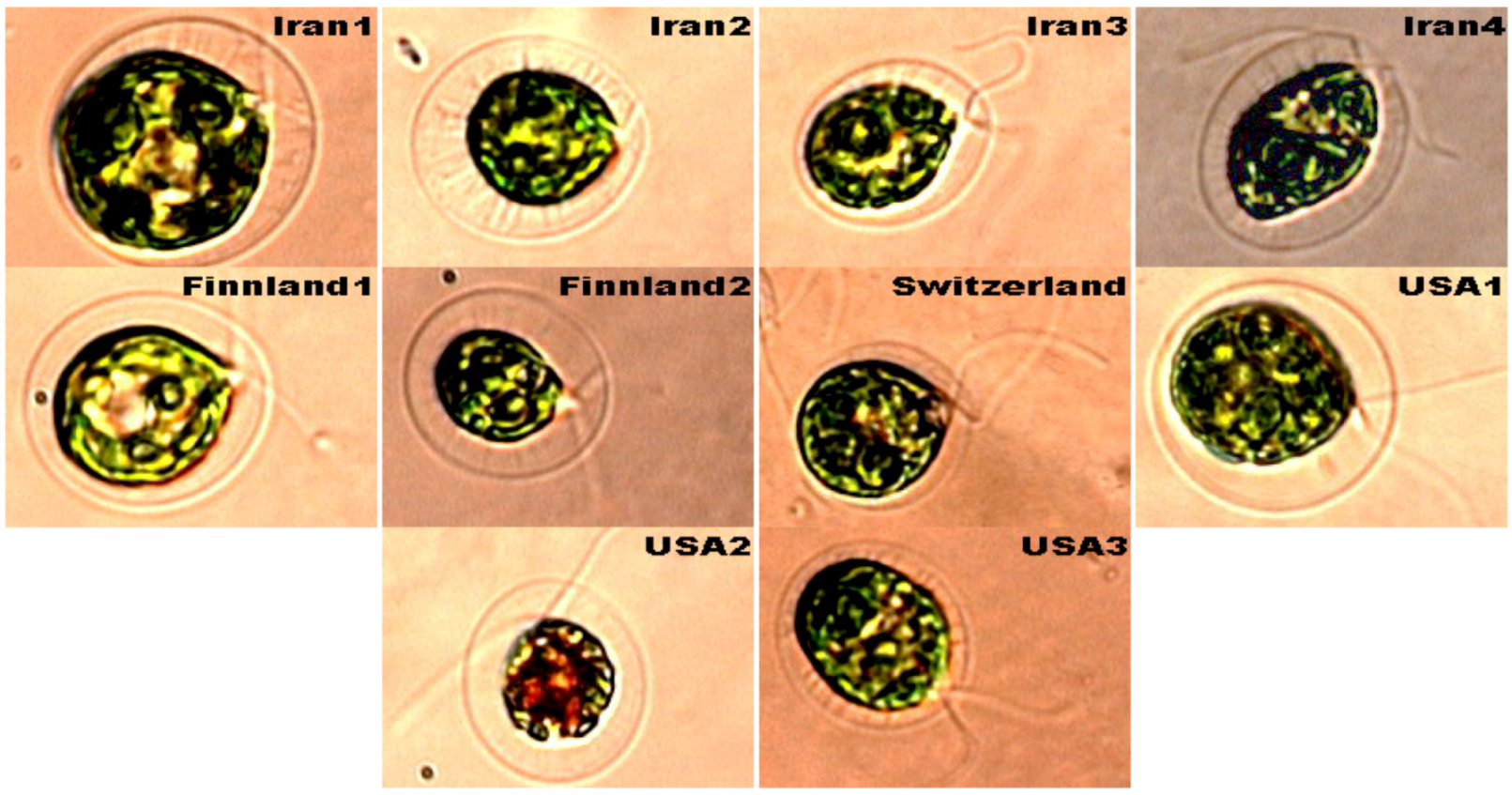{kind=link}
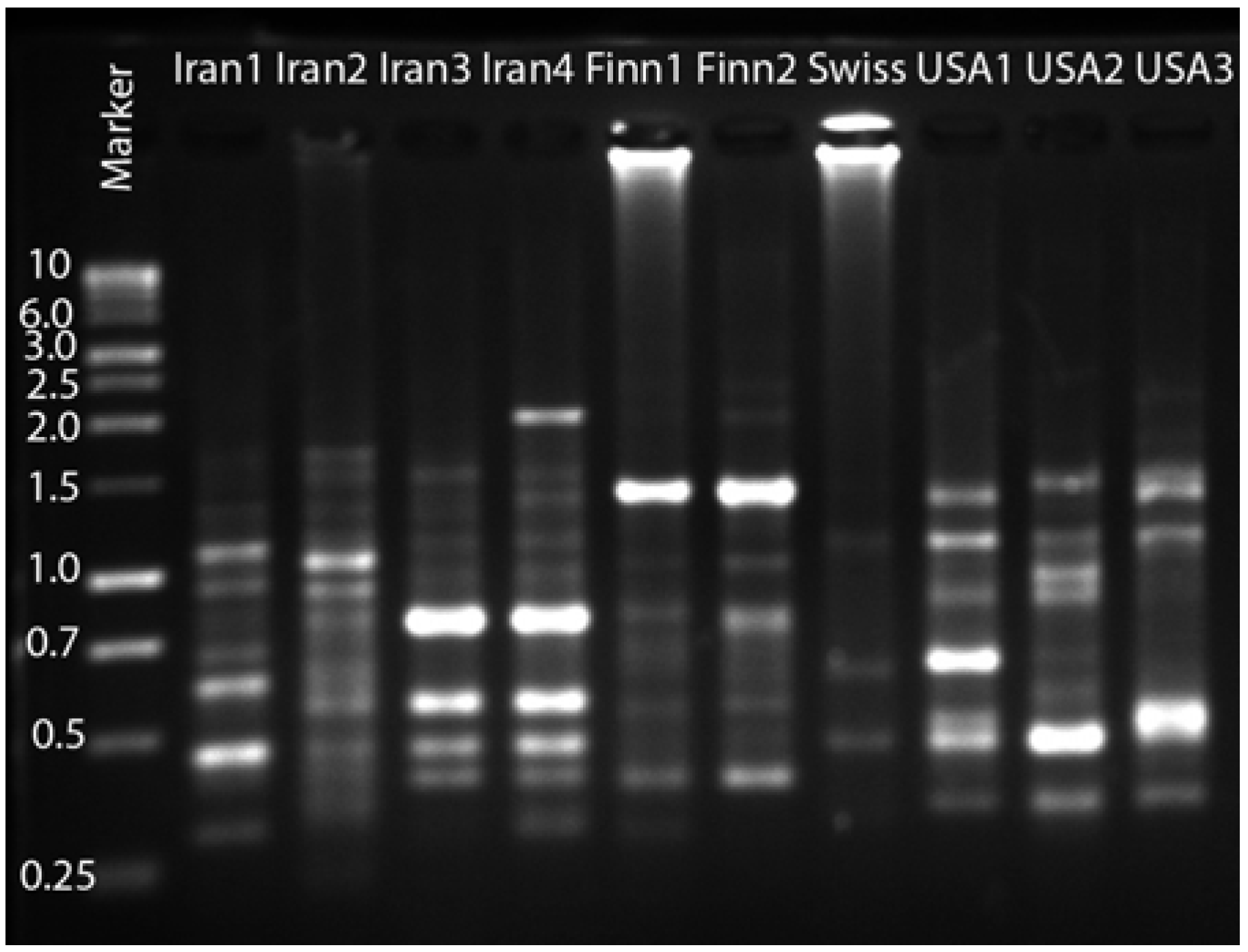{kind=link}
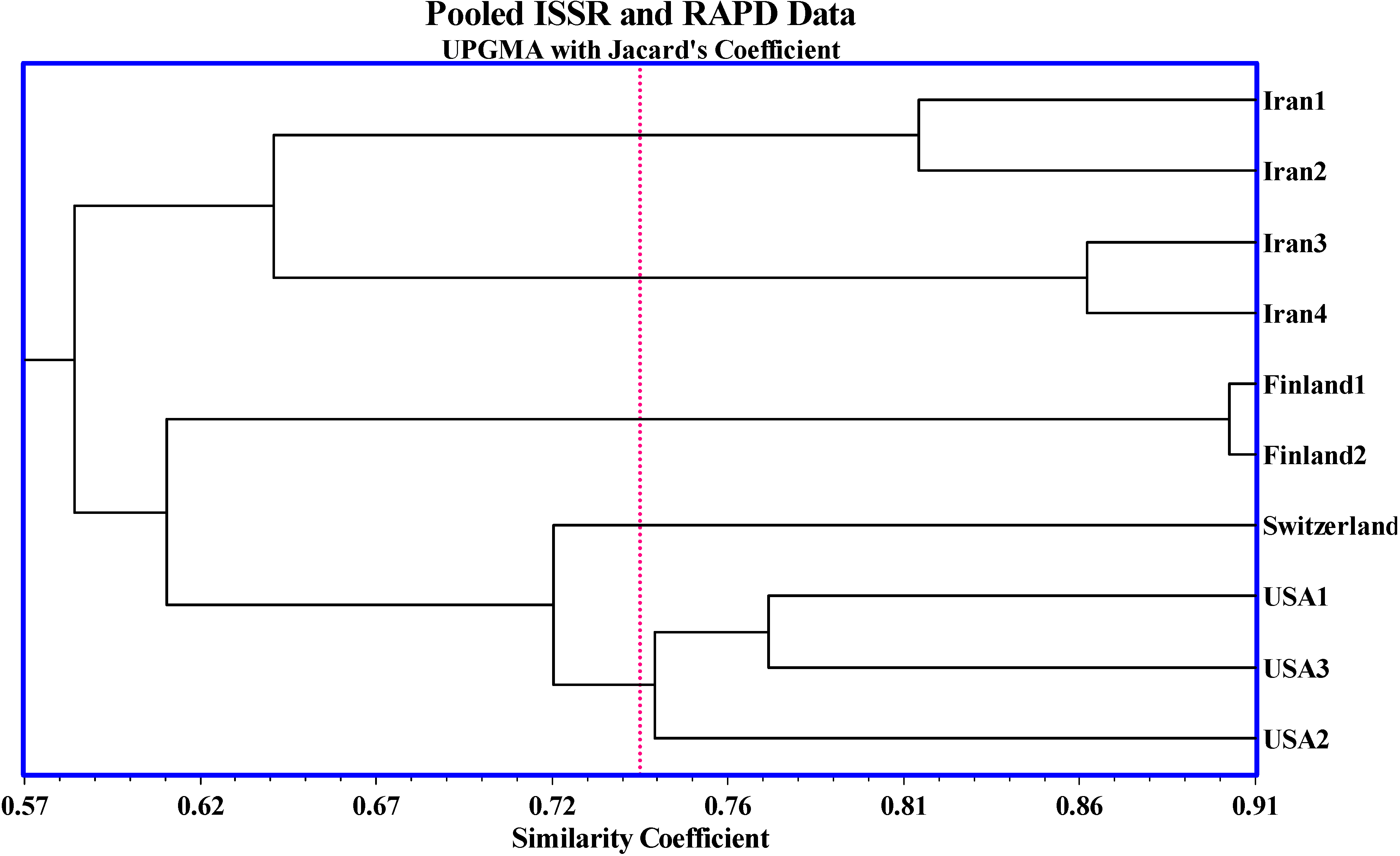{kind=link}
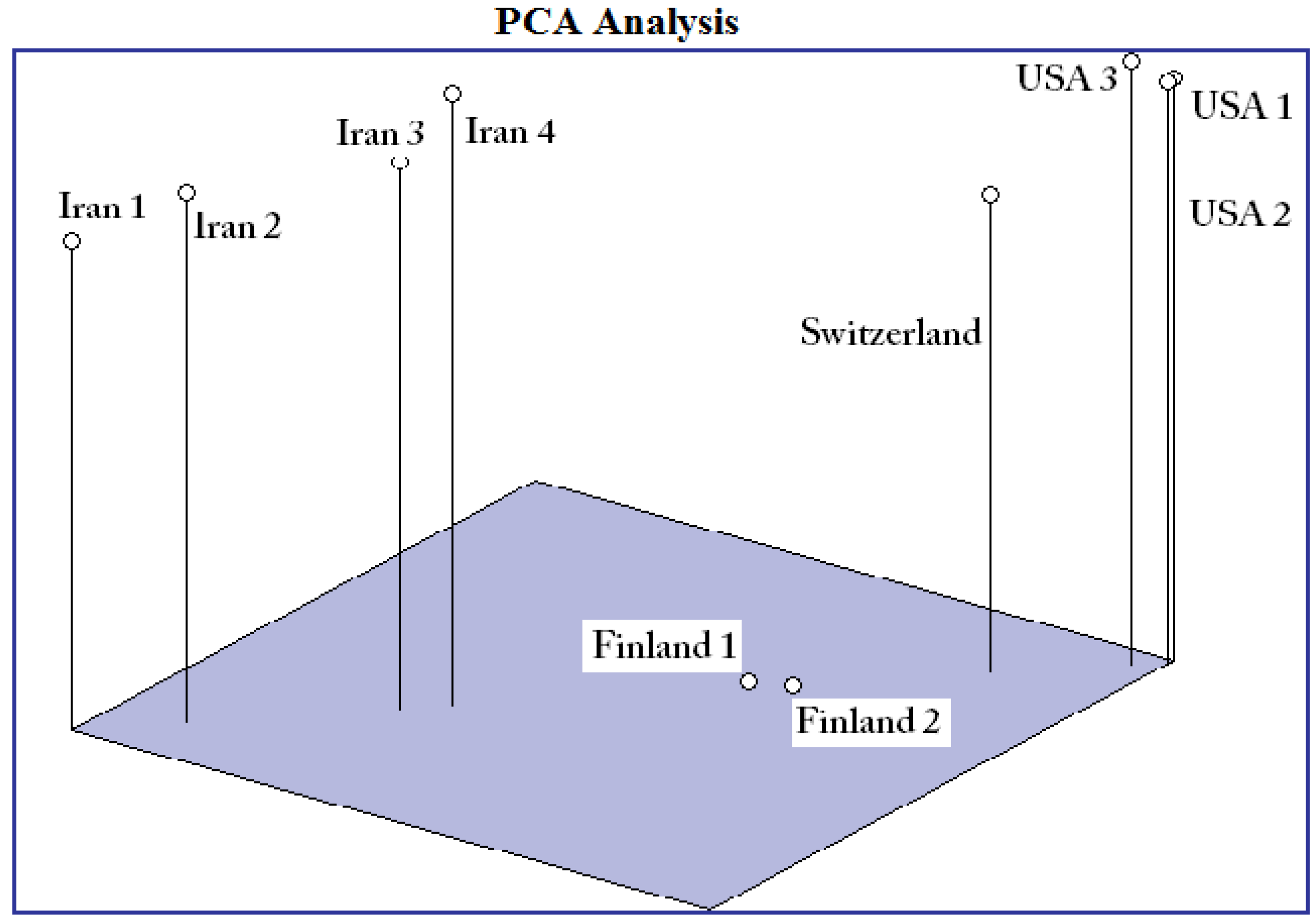{kind=link}
| Strains | Jacard’s similarity Index | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Iran1 | Iran2 | Iran3 | Iran4 | Finland1 | Finland2 | Switzer-land | USA1 | USA2 | USA3 | |
| Iran1 | 1.00 | |||||||||
| Iran2 | 0.47 | 1.00 | ||||||||
| Iran3 | 0.23 | 0.24 | 1.00 | |||||||
| Iran4 | 0.23 | 0.22 | 0.57 | 1.00 | ||||||
| Finland1 | 0.11 | 0.11 | 0.14 | 0.19 | 1.00 | |||||
| Finland2 | 0.10 | 0.11 | 0.14 | 0.16 | 0.64* | 1.00 | ||||
| Switzerland | 0.13 | 0.10 | 0.14 | 0.14 | 0.16 | 0.17 | 1.00 | |||
| USA1 | 0.14 | 0.08* | 0.16 | 0.12 | 0.14 | 0.16 | 0.32 | 1.00 | ||
| USA2 | 0.16 | 0.13 | 0.15 | 0.12 | 0.14 | 0.17 | 0.25 | 0.35 | 1.00 | |
| USA3 | 0.20 | 0.14 | 0.19 | 0.14 | 0.16 | 0.16 | 0.22 | 0.36 | 0.35 | 1.00 |
| Primer Name | Primer Sequence | Scored Bands | Ratio of Polymorphic Bands | Fragment Size range (Kb) | Annealing Temperature | |
|---|---|---|---|---|---|---|
| 1 | ISSR 1 | (CCA)5 | 25 | 24/25(96%) | 0.26–2.53 | 60 |
| 2 | ISSR 4 | (ATG)5 | 13 | 13/13(100%) | 0.45–1.19 | 50 |
| 3 | ISSR 5 | GTC(CT)8 | 19 | 19/19(100%) | 0.46–2.79 | 60 |
| 4 | ISSR 7 | GGGC(GA)8 | 29 | 29/29(100%) | 0.29–2.57 | 67 |
| 5 | ISSR 10 | (CAC)4RC | 35 | 35/35(100%)* | 0.44–2.68 | 57 |
| 6 | ISSR 11 | (GTG)3GC | 26 | 26/26(100%) | 0.36–2.54 | 60 |
| 7 | ISSR 12 | (GAG)3GC | 22 | 22/22(100%) | 0.47–2.1 | 58 |
| 8 | ISSR 13 | (GT)6GG | 15 | 15/15(100%) | 0.48–2.06 | 55 |
| 9 | ISSR 14 | (GA)6CC | 24 | 22/24(91.6%) | 0.14–2.11 | 55 |
| 10 | ISSR 15 | (GT)6CC | 16 | 16/16(100%) | 0.24–0.92 | 59 |
| 11 | ISSR 16 | (AC)8C | 21 | 21/21(100%) | 0.49–3.42 | 59 |
| 12 | ISSR 17 | (GA)8C | 24 | 24/24(100%) | 0.33–2.05 | 59 |
| 13 | ISSR 18 | (GA)8T | 26 | 26/26(100%) | 0.14–.052 | 57 |
| 14 | ISSR 19 | (GACA)4 | 22 | 22/22(100%) | 0.15–2.59 | 58 |
| 15 | RAPD 1 | ACA.ACT.GCT.C | 7 | 7/7(100%) | 0.42–1.44 | 37 |
| 16 | RAPD 2 | TGA.CTG.ACG.C | 6 | 5/6(83%) | o.35–2.1 | 42 |
| 17 | RAPD 3 | GCG.ATC.CCC.A | 7 | 9/9(100%) | 0.48–1.7 | 44 |
| 18 | RAPD 4 | ATC.GGG.TCC.G | 8 | 9/10(90%) | 0.62–2.16 | 45 |
| 19 | RAPD 5 | CAG.GCC.CTT.C | 11 | 17/17(100%) | 0.22–1.86 | 43 |
| 20 | RAPD 6 | AAT.CGG.GCT.G | 20 | 20/20 (100%)* | 0.43–1.8 | 42 |
| 21 | RAPD 7 | GGG.TAA.CGC.C | 10 | 6/6(100%) | 0.72–2.4 | 38 |
| 22 | RAPD 8 | CAA.TCG.CCG.T | 7 | 13/13(100%) | 0.59–2.28 | 42 |
| 23 | RAPD 9 | CAG.CAC.CCA.C | 6 | 9/9(100%) | 0.62–1.51 | 54 |
| 24 | RAPD 10 | AGG.TGA.CCG.T | 17 | 7/7(100%) | 0.17–1.2 | 40 |
| 25 | RAPD 11 | AGC.GCC.ATT.G | 17 | 11/11(100%) | 0.22–0.92 | 42 |
| 26 | RAPD 12 | GAG.AGC.CAA.C | 11 | 19/19(100%) | 0.51–2.1 | 32 |


3. Experimental
3.1. General
| Station | Golshahr Park (Karaj) | Kuye Karmandane shomali (Karaj) | Kuye Modares (Karaj) | Shemiranat (Tehran) |
| E (Longitude) | 50˚59′13″ | 50˚58′44″ | 50˚59′15″ | 51˚23′51″ |
| N (Latitude) | 35˚49′17″ | 35˚49′ 56″ | 35˚50′33″ | 35˚47′45″ |
3.2. DNA Isolation and PCR
3.3. Data Analysis
4. Conclusions
Acknowledgements
Supplementary Materials
Supplementary File 1References
- Hughes, D.A. Effects of dietary antioxidants on the immune function of middle-aged adults. Proc. Nutr. Soc. 1999, 58, 79–84. [Google Scholar] [CrossRef]
- Grossa, G.J.; Lockwood, S.F. Cardioprotection and myocardial salvage by a disodium disuccinate astaxanthin derivative (CardaxTM). Life Sci. 2004, 75, 215–224. [Google Scholar]
- Samuel, F.L.; Marc, S.P.; Stanley, L.; Hazen, Z.B.; Ferenc, Z. The effects of oral Cardax™ (disodium disuccinate astaxanthin) on multiple independent oxidative stress markers in a mouse peritoneal inflammation model: influence on 5-lipoxygenase in vitro and in vivo. Life Sci. 2006, 79, 162–174. [Google Scholar] [CrossRef]
- Kim, J.; Nam, S.; Kim, B.; Choi, W.; Lee, J.; Kim, W.; Choi, Y. Astaxanthin Improves Stem Cell Potency via an Increase in the Proliferation of Neural Progenitor Cells. Int. J. Mol. Sci. 2010, 11, 5109–5119. [Google Scholar] [CrossRef]
- Cianciola, E.; Popolizio, T.; Schneider, C.; Lane, C. Using Molecular-Assisted Alpha Taxonomy to Better Understand Red Algal Biodiversity in Bermuda. Diversity 2010, 2, 946–958. [Google Scholar] [CrossRef]
- Park, Y.; Lee, J.K.; Kim, N. Simple Sequence Repeat Polymorphisms (SSRPs) for Evaluation of Molecular Diversity and Germplasm Classification of Minor Crops. Molecules 2009, 14, 4546–4569. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.C.; Massola, J.N.S.; Hernandez-Marine, M.; Romo, S.; Moura, A.N. Taxonomic investigation using DNA fingerprinting in Geitlerinema species (Oscillatoriales, Cyanobacteria). Phycol. Res. 2007, 55, 214–221. [Google Scholar] [CrossRef]
- Arif, I.; Bakir, M.; Khan, H.; Al Farhan, A.; Al Homaidan, A.; Bahkali, A.; Al Sadoon, M.; Shobrak, M. A Brief Review of Molecular Techniques to Assess Plant Diversity. Int. J. Mol. Sci. 2010, 11, 2079–2096. [Google Scholar] [CrossRef]
- Hall, M.M.; Vis, M.L. Genetic variation in Batrachospermum helminthosum (Batrachospermales, Rhodophyta) among and within stream reaches using intersimple sequence repeat molecular markers. Phycol. Res. 2002, 50, 155–162. [Google Scholar] [CrossRef]
- Bornet, B.; Antoine, E.; Francoise, S.; Marcaillou, Le.; Baut, C. Development of sequence characterized amplified region markers from inter simple sequence repeat fingerprints for the molecular detection of toxic phytoplankton Alexanndrium catenella (Dinophyceae) and pseudo -Nitzschia Pseudodelicatissiam (Bacillariophyceae) from French coastline waters. J. Phycol. 2005, 41, 704–711. [Google Scholar] [CrossRef]
- Dayananda, C.; Sarada, R.; Kumar, V.; Aswathanarayana, R.G. Isolation and characterization of hydrocarbon producing green alga Botryococcus braunii from Indian freshwater bodies. Electron. J. Biotechnol. 2007, 10, 78–91. [Google Scholar]
- House, D.L.; Sherwood, R.A.; Vis, L.M. Comparison of three organelle markers for phylogeographic inference in Batrachospermum helminthosum (Batrachospermales, Rhodophyta) from North America. Phycol. Res. 2008, 56, 69–75. [Google Scholar] [CrossRef]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple sequence repeat (SSR) anchored polymerase chain reaction amplification. Genetics 1994, 20, 176–183. [Google Scholar]
- Bornet, B.; Branchard, M. Nonanchored Inter Simple Sequence Repeat (ISSR) Markers: Reproducible and Specific Tools for Genome Fingerprinting. Plant Mol. Biol. Rep. 2001, 19, 209–215. [Google Scholar] [CrossRef]
- Bardakci, F. Random Amplified Polymorphic DNA (RAPD) Markers. Turk. J. Biol. 2001, 25, 185–196. [Google Scholar]
- Martı´nez, R.; An˜ı´barro, C.; Ferna´ndez, S. Genetic variability among Alexandrium tamarense and Alexandrium minutum strains studied by RAPD banding pattern analysis. Harmful Algae 2006, 5, 599–607. [Google Scholar] [CrossRef]
- Prevost, A.; Wilkinson, M.J. A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 1999, 98, 107–112. [Google Scholar] [CrossRef]
- Singh, R.; Zaki, N.M.; Ting, N.; Rosli, R.; Soon-Guan, T.; Leslie, L.E.; Ithnin, M.; Cheah, S. Exploiting an oil palm EST database for the development of gene-derived SSR markers and their exploitation for assessment of genetic diversity. Biologia 2008, 63, 227–235. [Google Scholar] [CrossRef]
- Bhau, B.S.; Medhi, K.; Sarkar, T.; Saikia, S.P. PCR based molecular characterization of Nepenthes khasiana Hook Pitcher. Plant Genet. Resour. Crop. Evol. 2009, 56, 1183–1193. [Google Scholar] [CrossRef]
- Ferna´ndez, M.; Figueiras, A.; Benito, C. The use of ISSR and RAPD markers for detecting DNA polymorphism, genotype identification and genetic diversity among barley cultivars with known origin. Theor. Appl. Genet. 2002, 104, 845–851. [Google Scholar] [CrossRef]
- Zhao, K.G.; Zhou, M.Q.; Chen, L.Q. Genetic diversity and discrimination of Chimonanthus praecox (L.) Link germplasm using ISSR and RAPD markers. HortScience 2007, 42, 1144–1148. [Google Scholar]
- Rahimmalek, M.; Bahreininejad, B.; Khorrami, M.; Tabatabaei, B.E.S. Genetic Variability and Geographic Differentiation in Thymus daenensis subsp. daenensis, an Endangered Medicinal Plant, as Revealed by Inter Simple Sequence Repeat (ISSR) Markers. Biochem. Genet. 2009, 47, 831–842. [Google Scholar] [CrossRef]
- Dellaporta, S.L.; Wood, J.; Hicks, J.B. A plant DNA mini-preparation Version II. Plant Mol. Biol. Reptr. 1983, 1, 19–21. [Google Scholar] [CrossRef]
- Bornet, B.; Antoine, E.; Bardouil, M.; Marcaillou-Le, Baut, C. ISSR as new markers for genetic characterization and evaluation of relationships among phytoplankton. J. Appl. Phycol. 2004, 16, 285–290. [Google Scholar] [CrossRef]
- Rohlf, F. NTSYS-pc Numerical Taxonomy and Multivariate Analysis System; Exeter Software: New York, NY, USA, 2001. [Google Scholar]
- Huangfu, C.; Song, X.; Qiang, S. ISSR variation within and among wild Brassica juncea populations: implication for herbicide resistance evolution. Genet. Resour. Crop. Evol. 2009, 56, 913–924. [Google Scholar] [CrossRef]
- Sokal, R.R.; Sneath, P.H.A. Principles of Numerical Taxonomy; W. H. Freeman: San Francisco, CA, USA, 1963. [Google Scholar]
- Sample Availability: Samples of the ten of H. pluvialis strains are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mostafa, N.; Omar, H.; Tan, S.G.; Napis, S. Studies on the Genetic Variation of the Green Unicellular Alga Haematococcus pluvialis (Chlorophyceae) Obtained from Different Geographical Locations Using ISSR and RAPD Molecular Marker. Molecules 2011, 16, 2599-2608. https://doi.org/10.3390/molecules16032599
Mostafa N, Omar H, Tan SG, Napis S. Studies on the Genetic Variation of the Green Unicellular Alga Haematococcus pluvialis (Chlorophyceae) Obtained from Different Geographical Locations Using ISSR and RAPD Molecular Marker. Molecules. 2011; 16(3):2599-2608. https://doi.org/10.3390/molecules16032599
Chicago/Turabian StyleMostafa, Noroozi, Hishamuddin Omar, Soon Guan Tan, and Suhaimi Napis. 2011. "Studies on the Genetic Variation of the Green Unicellular Alga Haematococcus pluvialis (Chlorophyceae) Obtained from Different Geographical Locations Using ISSR and RAPD Molecular Marker" Molecules 16, no. 3: 2599-2608. https://doi.org/10.3390/molecules16032599