Immunosuppressive Activity of 8-Gingerol on Immune Responses in Mice

Abstract
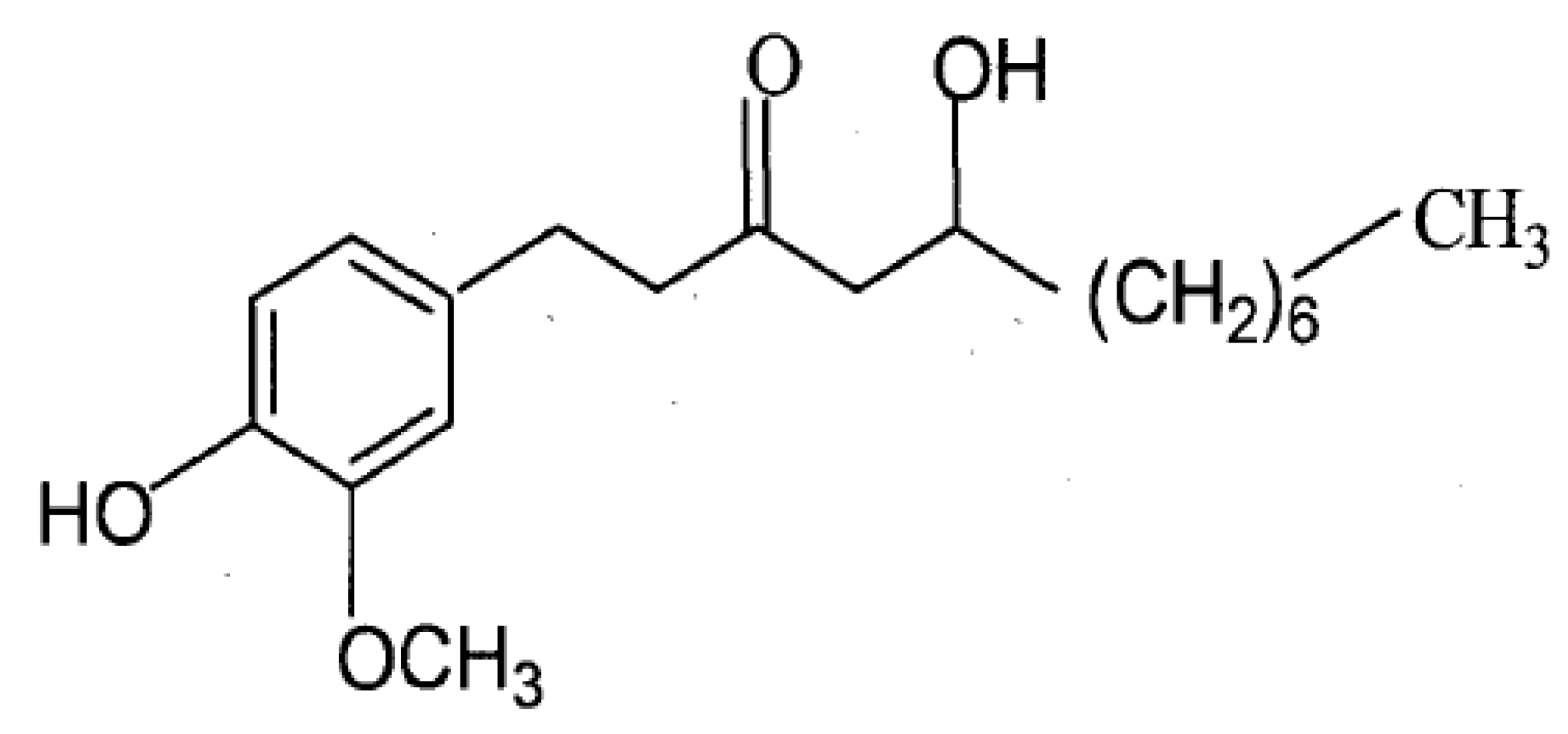
:1. Introduction

2. Results and Discussion
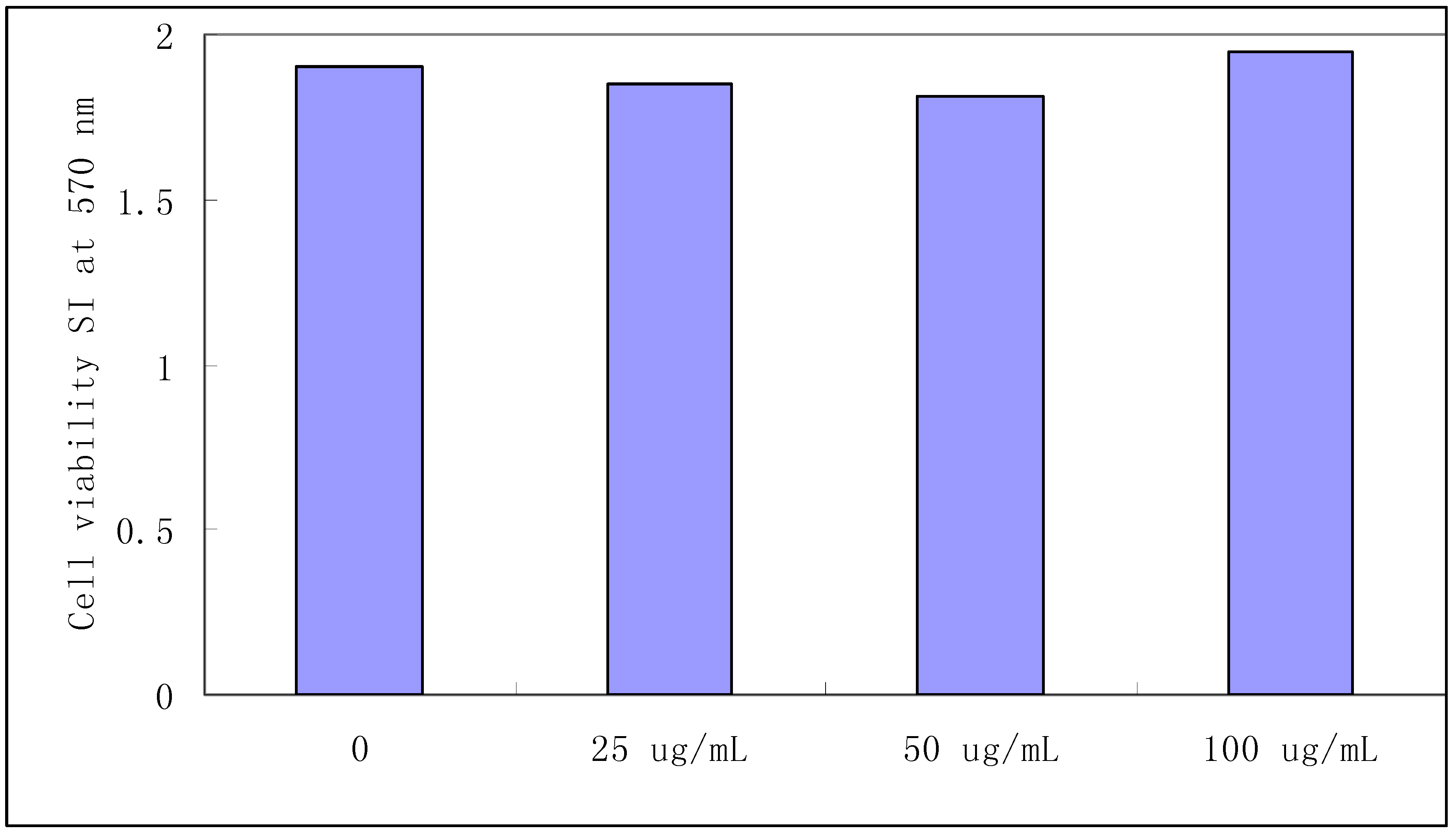
2.1. Splenocyte viability

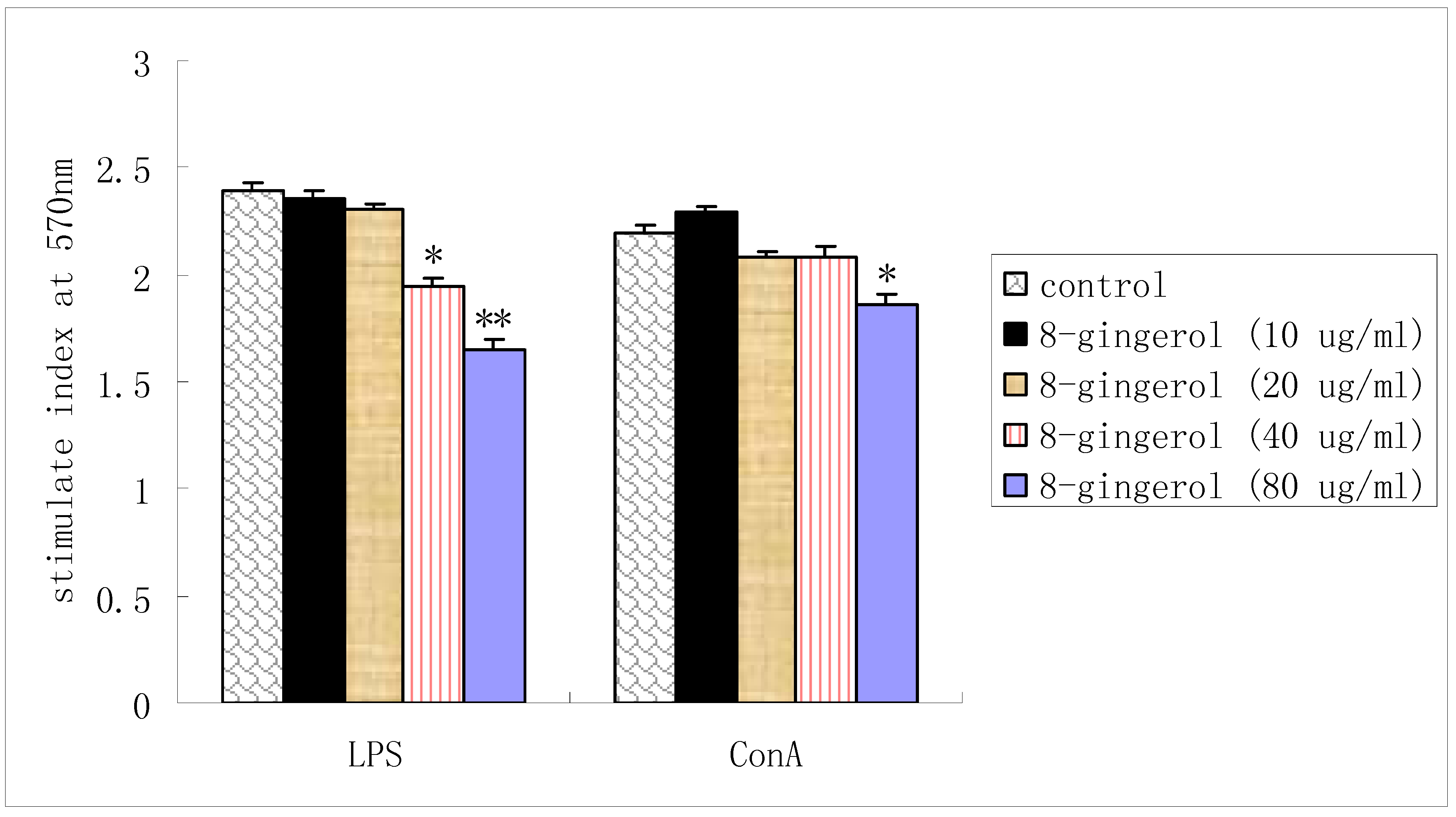
2.2. Proliferation of splenocytes in vitro

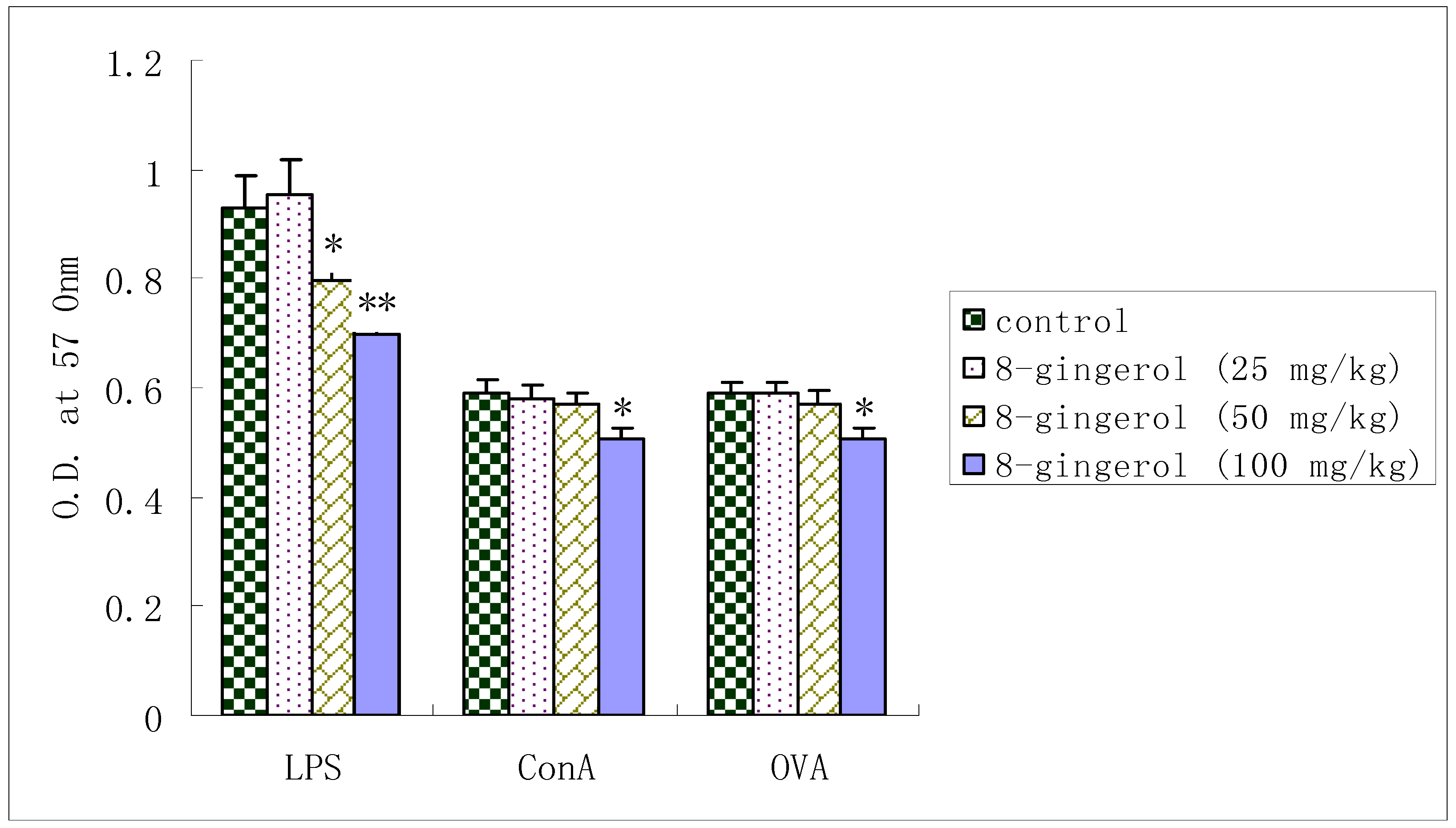
2.3. Proliferation of splenocytes from OVA-immunized mice
2.4. Effect of 8-gingerol on splenocyte surface markers in OVA-immunized mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | CD3+ CD19-% | CD3- CD19+% | CD3+CD19+% |
|---|---|---|---|
| Negative group | 26.17 ± 1.14 | 27.62 ± 1.25 | 1.68 ± 0.19 |
| OVA Control | 32.65 ± 1.22 | 34.11 ± 1.54 | 2.51 ± 1.29 |
| OVA+ 8-gingerol (25 mg/kg) | 33.54 ± 2.05 | 33.63 ± 1.87 | 2.85 ± 1.07 |
| OVA+ 8-gingerol (50 mg/kg) | 31.72 ± 2.06 | 30.32 ± 1.94* | 2.03 ± 0.76 |
| OVA+ 8-gingerol (100 mg/kg) | 28.65 ± 2.15* | 24.8 ± 2.13** | 2.66 ± 0.88 |
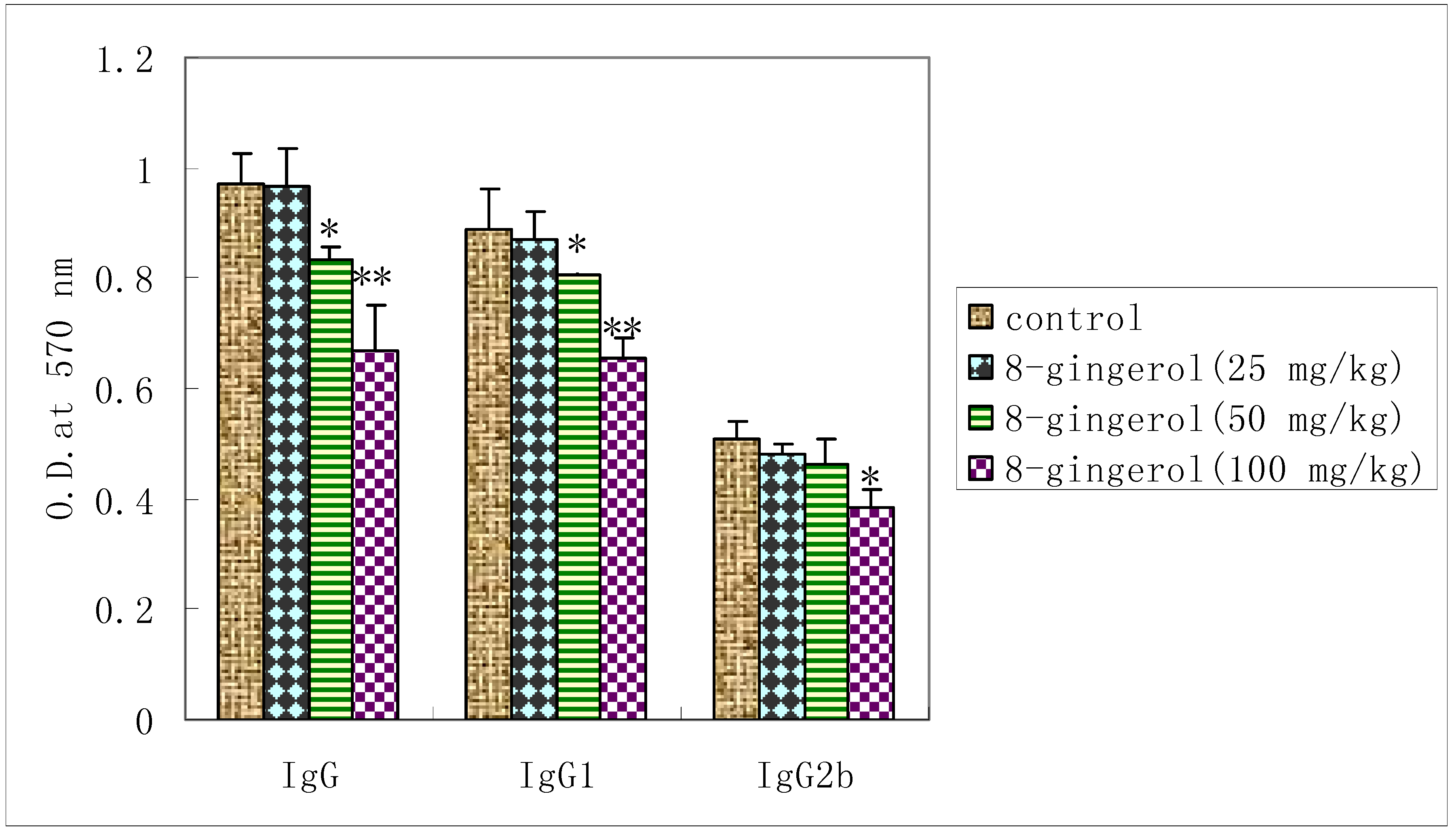
2.5. Effect of 8-gingerol on OVA-specific serum antibody response
2.6. Discussion


3. Experimental
3.1. Materials
3.2. Experimental animals
3.3. Cell viability assay
3.4. In vitro splenocyte proliferation assay
3.5. Administration and immunization
3.6. In vivo splenocyte proliferation
3.7. Analysis of cell surface markers by flow cytometry
3.8. Measurement of OVA-specific antibody levels by ELISA
3.9. Statistical analysis
4. Conclusions
Acknowledgements
References and Notes
- Wong, S.H.Y. Therapeutic drug monitoring for immunosuppressants. Clin. Chim. Acta 2001, 313, 241–253. [Google Scholar] [CrossRef]
- Serkova, N.; Brand, A.; Christians, U.; Leibfritz, D. Evaluation of the effects of immunosuppressants on neuronal and glial cells in vitro by multinuclear magnetic resonance spectroscopy. Biochim. Biophys. Acta 1996, 1314, 93–104. [Google Scholar]
- Zhang, D.H.; Marconi, A.; Xu, L.M.; Uzan, G. Tripterine inhibits the expression of adhesion molecules in activated endothelial cells. J. Leukoc. Biol. 2006, 80, 309–311. [Google Scholar] [CrossRef]
- Ye, M.; Xie, W.D.; Lei, F.; Meng, Z.; Zhao, Y.N.; Su, H.; Du, L.J. Brazilein, an important immunosuppressive component fromCaesalpinia sappanL. Int. Immunopharmacol. 2006, 6, 426–411. [Google Scholar] [CrossRef]
- Feng, H.; Yamaki, K.; Takano, H.; Inoue, K.; Yanagisawa, R.; Yoshino, S. Effect of sinomenine on collagen-induced arthritis in mice. Autoimmunity 2007, 40, 532–539. [Google Scholar] [CrossRef]
- Afzal, M.; Al-Hadidi, D.; Menon, M.; Pesek, J.; Dhami, M.S. Ginger: An ethnomedical, chemical and pharmacological review. Drug Metabol. Drug Interac. 2001, 18, 159–190. [Google Scholar]
- Wang, W.; Li, C.Y.; Wen, X.D.; Li, P.; Qi, L.W. Simultaneous determination of 6-gingerol, 8-gingerol, 10-gingerol and 6-shogaol in rat plasma by liquid chromatography-mass spectrometry: Application to pharmacokinetics. J. Chromatogr. B 2009, 877, 671–679. [Google Scholar] [CrossRef]
- Nie, H.; Meng, L.Z.; Zhang, H.; Zhang, J.Y.; Yin, Z.; Huang, X.S. Analysis of anti-platelet aggregation components of Rhizoma Zingiberis using chicken thrombocyte extract and high performance liquid chromatography. Chin. Med. J. (Engl.) 2008, 121, 1226–1229. [Google Scholar]
- Nurtjahja-Tjendraputra, E.; Ammit, A.J.; Roufogalis, B.D.; Tran, V.H.; Duke, C.C. Effective anti-platelet and COX-1 enzyme inhibitors from pungent constituents of ginger. Thromb. Res. 2003, 111, 259–265. [Google Scholar] [CrossRef]
- Ghayur, M.N.; Gilani, A.H.; Afridi, M.B.; Houghton, P.J. Cardiovascular effects of ginger aqueous extract and its phenolic constituents are mediated through multiple pathways. Vascul. Pharmacol. 2005, 43, 234–241. [Google Scholar] [CrossRef]
- Koh, E.M.; Kim, H.J.; Kim, S.; Choi, W.H.; Choi, Y.H.; Ryu, S.Y.; Kim, Y.S.; Koh, W.S.; Park, S.Y. Moduation of Macrophage Functions by Compounds Isolated from Zingiber officinale. Planta Med. 2009, 75, 148–151. [Google Scholar] [CrossRef]
- Lantz, R.C.; Chen, G.J.; Sarihan, M.; Solyom, A.M.; Jolad, S.D.; Timmermann, B.N. The effect of extracts from ginger rhizome on inflammatory mediator production. Phytomedicine 2007, 14, 123–128. [Google Scholar] [CrossRef]
- Abdel-Aziz, H.; Nahrstedt, A.; Petereit, F.; Windeck, T.; Ploch, M.; Verspohl, E.J. 5-HT3 receptor blocking activity of arylalkanes isolated from the rhizome of Zingiber officinale. Planta Med. 2005, 71, 609–616. [Google Scholar] [CrossRef]
- Cao, Y.G.; Song, Y.; An, N.; Zhou, C.F.; Deng, X.M. The effects of telocinobufagin isolated from Chan Su on the activation and cytokine secretion of immunocytes in vitro. Fund. Clin. Pharmacol. 2009, 23, 457–464. [Google Scholar] [CrossRef]
- Byun, J.A.; Ryu, M.H.; Lee, J.K. The immunomodulatory effects of 3-monochloro-1, 2-propanediol on murine splenocyte and peritoneal macrophage function in vitro. Toxicol. In Vitro 2006, 20, 272–278. [Google Scholar] [CrossRef]
- Caballero-Hernandez, D.; Weberb, R.J.; Hicks, M.E.; Tamez-Guerra, R.; Rodriguez-Padilla, C.; Tamez-Guerra, P.; Rice, K.C.; Ananthan, S.; Gomez-Flores, R. Potentiation of rat lymphocyte proliferation by novel non-peptidic synthetic opioids. Int. Immunopharmacol. 2005, 5, 1271–1278. [Google Scholar] [CrossRef]
- Dearman, R.J.; Caddick, H.; Basketter, D.A.; Kimber, I. Divergent antibody iso-type responses induced inmice by systemic exposure to proteins: A comparison of ovalbumin with bovine serum albumin. Food Chem. Toxicol. 2000, 38, 351–360. [Google Scholar] [CrossRef]
- Wey, S.P.; Wu, H.Y.; Chang, F.C.; Jan, T.R. Methamphetamine and diazepam suppress antigen-specific cytokine expression and antibody production in ovalbumin-sensitized BALB/c mice. Toxicol. Lett. 2008, 181, 157–162. [Google Scholar] [CrossRef]
- Shan, X.; Chen, L.; Cao, M.; Xu, L.; Zhang, S. Effects of human soluble BAFF synthesized in Escherichia coli on CD4+ and CD8+ T lymphocytes as well as NK cells in mice. Physiol. Res. Praha. 2006, 55, 301–307. [Google Scholar]
- Li, Y.; Li, X.F.; Ma, Y.Y.; Fang, N.; Russe, J.; Ma, D.; Sun, X.R.; Han, X.; Yang, H.S.; Kinuya, S. Changes in the levels of CD4+ and CD8+ T-lymphocytes after strontium-89 chloride therapy for painful bone metastases in patients correlate with treatment efficacy. Cancer Biother. Radiopharm. 2007, 22, 367–373. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the 8-gingerol are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lu, J.; Guan, S.; Shen, X.; Qian, W.; Huang, G.; Deng, X.; Xie, G. Immunosuppressive Activity of 8-Gingerol on Immune Responses in Mice. Molecules 2011, 16, 2636-2645. https://doi.org/10.3390/molecules16032636
Lu J, Guan S, Shen X, Qian W, Huang G, Deng X, Xie G. Immunosuppressive Activity of 8-Gingerol on Immune Responses in Mice. Molecules. 2011; 16(3):2636-2645. https://doi.org/10.3390/molecules16032636
Chicago/Turabian StyleLu, Jing, Shuang Guan, Xue Shen, Wenhui Qian, Guoren Huang, Xuming Deng, and Guanghong Xie. 2011. "Immunosuppressive Activity of 8-Gingerol on Immune Responses in Mice" Molecules 16, no. 3: 2636-2645. https://doi.org/10.3390/molecules16032636