Astragaloside IV Improves Metabolic Syndrome and Endothelium Dysfunction in Fructose-Fed Rats

Abstract
:1. Introduction

2. Results and Discussion
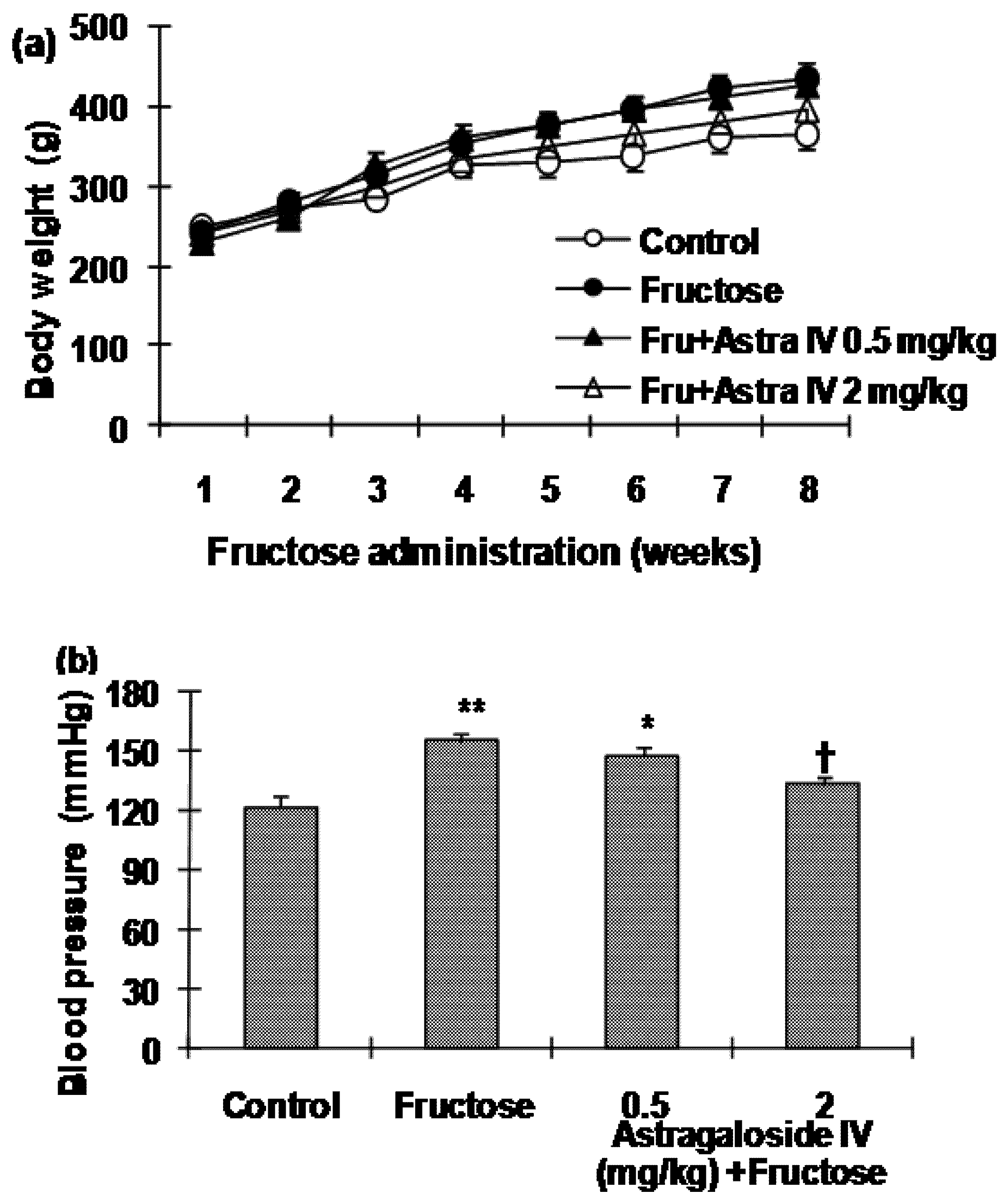
2.1. Bodyweight and Blood Pressure

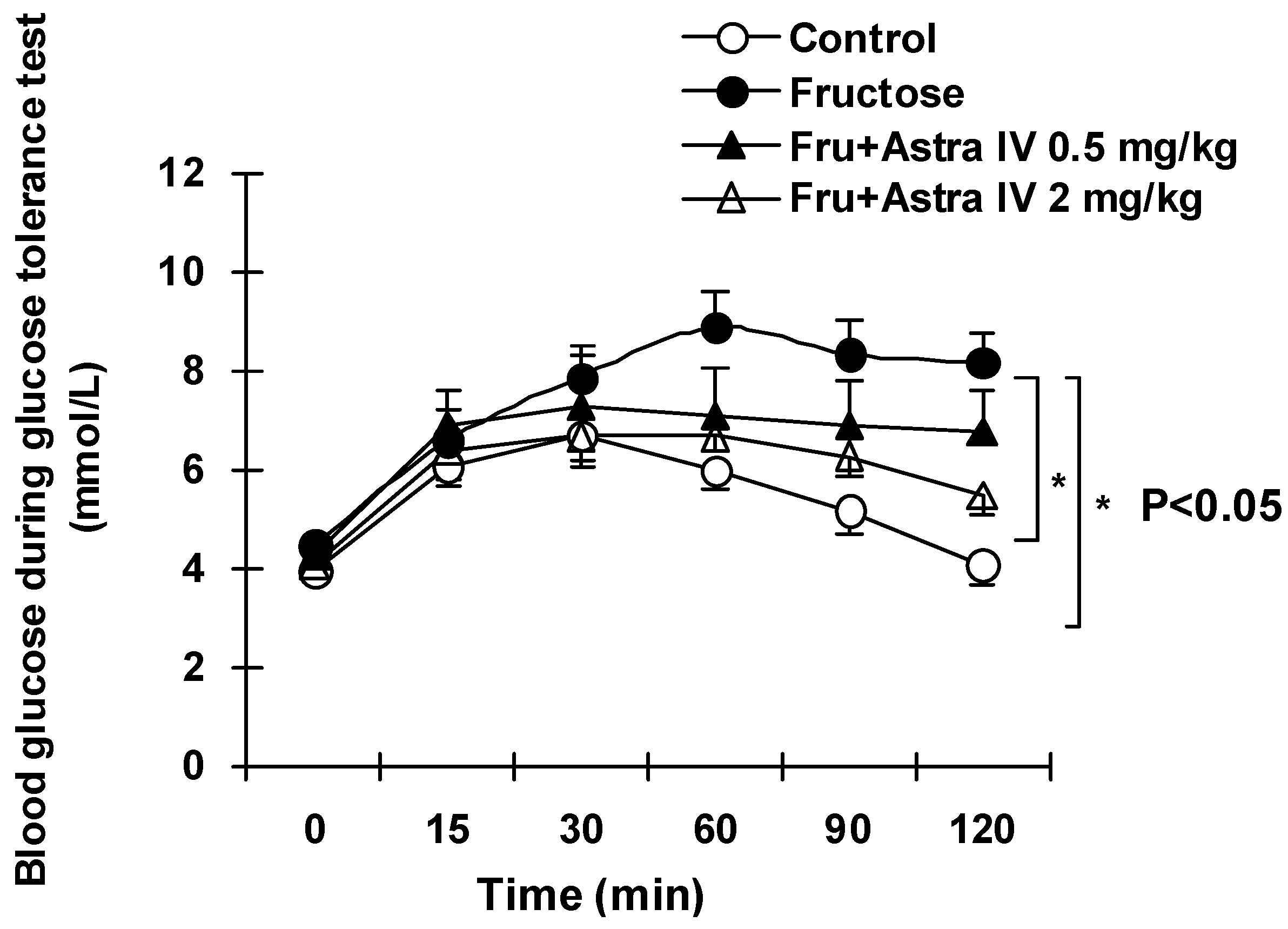
2.2. Blood Glucose, Lipids and Glucose Tolerance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Fructose | Fructose + 0.5 mg/kg astragaloside IV | Fructose + 2 mg/kg astragaloside IV | |
|---|---|---|---|---|
| Fasting glucose (mmol/L) | 4.0 ± 0.6 | 4.5 ± 0.8 | 4.3 ± 0.7 | 4.1 ± 0.9 |
| Serum insulin (ng/mL) | 1.22 ± 0.25 | 3.95 ± 0.32** | 2.77 ± 0.20† | 2.13 ± 0.20† |
| Triglyceride (mmol/L) | 1.18 ± 0.13 | 2.53 ± 0.18** | 1.93 ± 0.17† | 1.23 ± 0.12†† |
| Total cholesterol (mmol/L) | 1.91 ± 0.04 | 2.05 ± 0.06 | 1.99 ± 0.07 | 1.95 ± 0.10 |
| LDL-C (mmol/L) | 0.42 ± 0.03 | 0.44 ± 0.06 | 0.45 ± 0.04 | 0.41 ± 0.06 |
| HDL-C (mmol/L) | 1.17 ± 0.06 | 1.30 ± 0.11 | 1.24 ± 0.10 | 1.22 ± 0.13 |

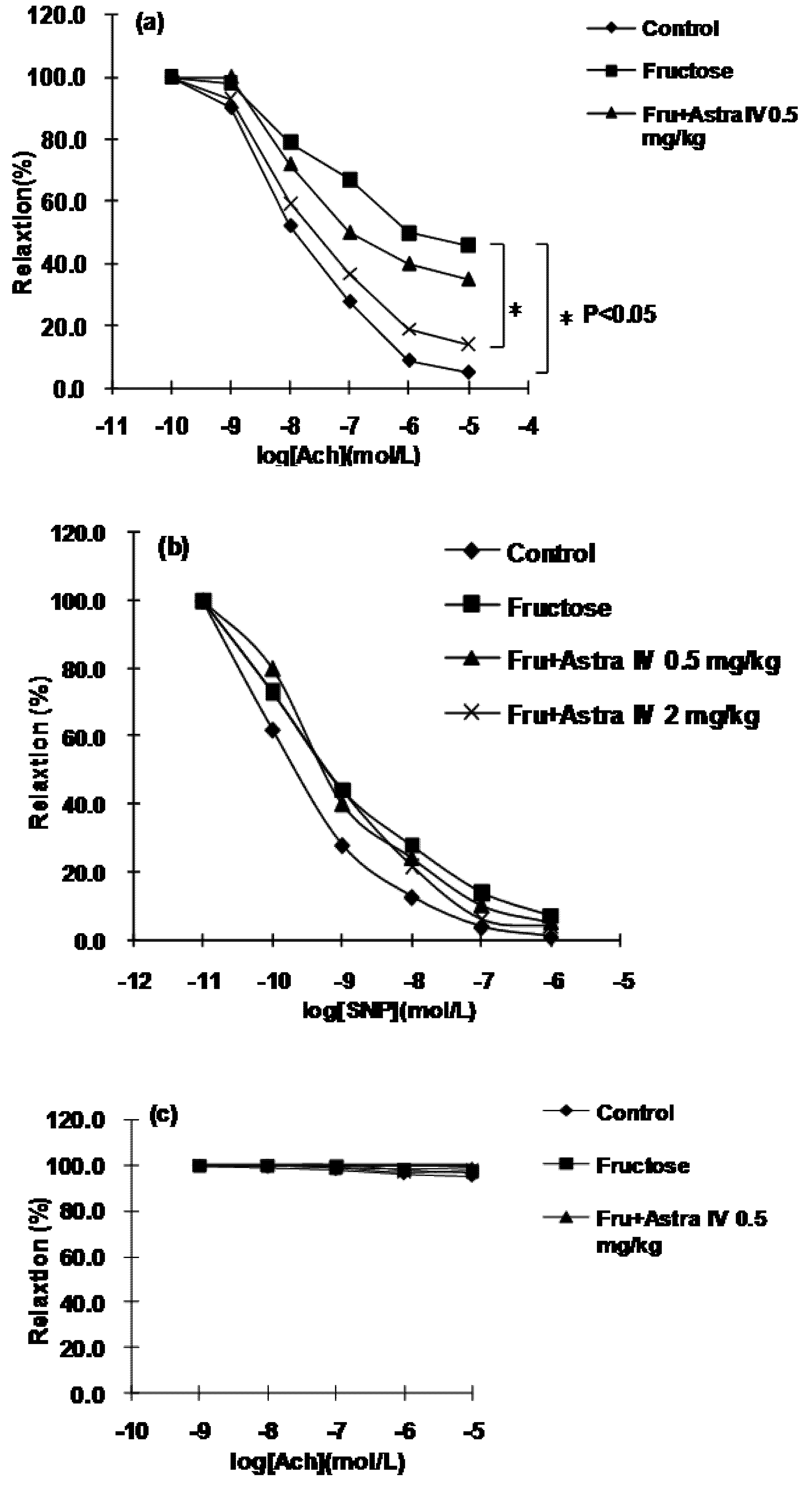
2.3. Vessel Relaxation

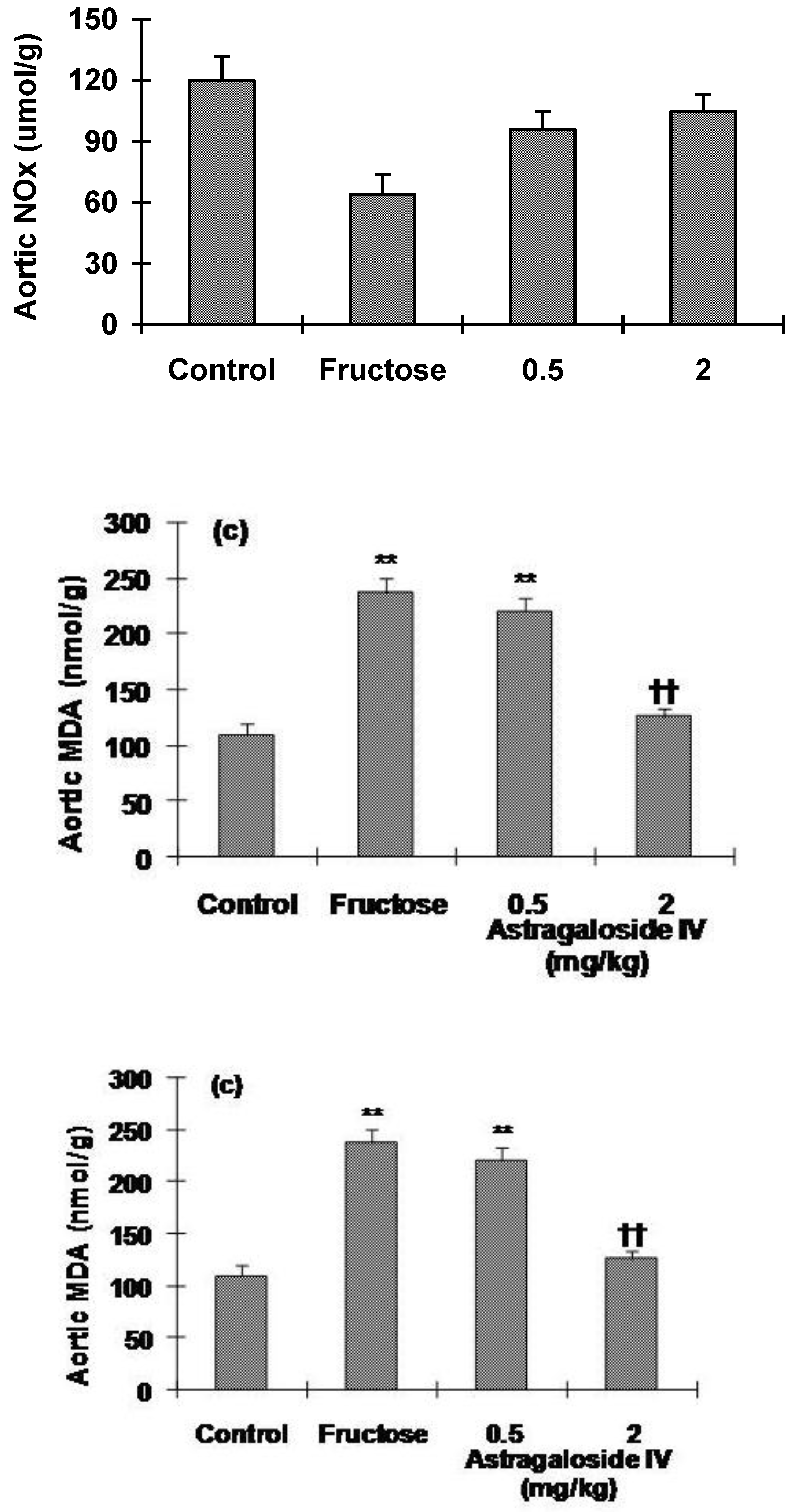
2.4. Aortic Levels of NOx and cGMP

3. Experimental
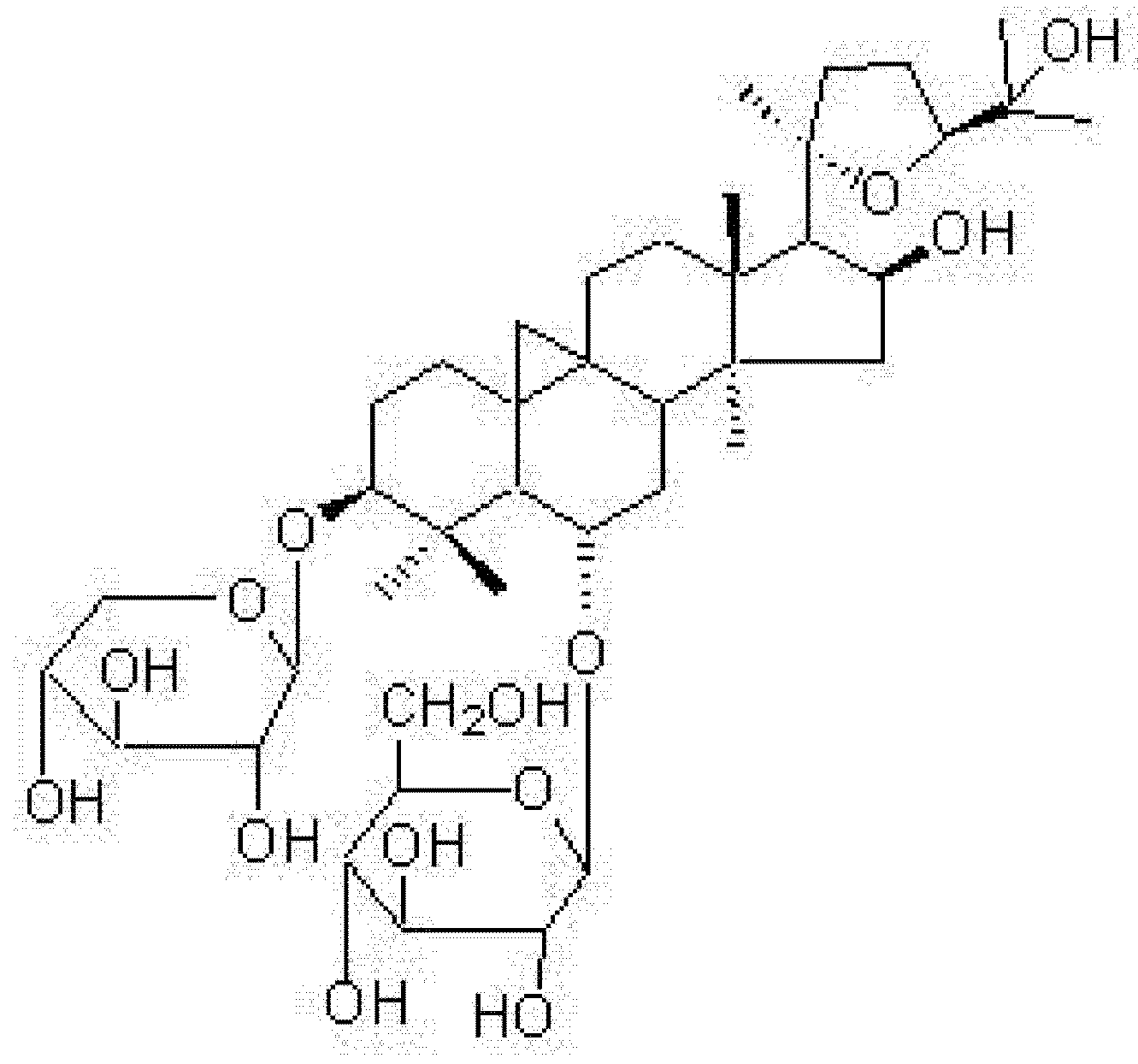
3.1. Source of Astragaloside IV
3.2. Animals and Fructose Feeding
3.3. Blood Pressure Measurement and GTT
3.4. Determination of Blood Lipids, Insulin, Tissue NO Metabolites and Malondialdehyde
3.5. Preparation of Thoracic Aortic Rings and Tension Recording
3.6. Determination of Tissue cGMP
3.7. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Zhang, W.D.; Chen, H.; Zhang, C.; Li, H.L.; Liu, R.H.; Chen, H.Z. Astragaloside IV from Astragalus membranaceus shows cardioprotection during myocardial ischemia in vivo and in vitro. Planta Med. 2006, 72, 4–8. [Google Scholar] [CrossRef]
- Luo, Y.; Qin, Z.; Hong, Z.; Zhang, X.M.; Ding, D.; Fu, J.H.; Zhang, W.D.; Chen, J. Astragaloside IV protects against ischemic brain injury in a murine model of transient focal ischemia. Neurosci. Lett. 2004, 363, 218–223. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Fan, Y.; Kong, J.L.; Wu, D.Z.; Hu, Z.B. Effects of components isolated from Astragalus membranaceus Bunge on cardiac function injured by myocardial ischemia reperfusion in rats. Chin. J. Chin. Mater. Med. 2000, 25, 300–302. [Google Scholar]
- Qu, Y.Z.; Li, M.; Zhao, Y.L.; Zhao, Z.W.; Wei, X.Y.; Liu, J.P.; Gao, L.; Gao, G.D. Astragaloside IV attenuates cerebral ischemia-reperfusion-induced increase in permeability of the blood-brain barrier in rats. Eur. J. Pharmacol. 2009, 606, 137–141. [Google Scholar] [CrossRef]
- Zhang, W.D.; Zhang, C.; Wang, X.H.; Gao, P.J.; Zhu, D.L.; Chen, H.; Liu, R.H.; Li, H.L. Astragaloside IV dilates aortic vessels from normal and spontaneously hypertensive ratsthrough endothelium-dependent and endothelium-independent ways. Planta Med. 2006, 72, 621–626. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.H.; Zhong, M.F.; Liu, R.H.; Li, H.L.; Zhang, W.D.; Chen, H. Mechanisms underlying vasorelaxant action of astragaloside IV in isolated rat aortic rings. Clin. Exp. Pharmacol. Physiol. 2007, 34, 387–392. [Google Scholar] [CrossRef]
- Xu, M.E.; Xiao, S.Z.; Sun, Y.H.; Ou-Yang, Y.; Zheng, X.X. Effects of astragaloside IV on pathogenesis of metabolic syndrome in vitro. Acta Pharmacol. Sin. 2006, 27, 229–236. [Google Scholar] [CrossRef]
- Liu, T.P.; Lee, C.S.; Liou, S.S.; Liu, I.M.; Cheng, J.T. Improvement of insulin resistance by Acanthopanax senticosus root in fructose-rich chow-fed rats. Clin. Exp. Pharmacol. Physiol. 2005, 32, 649–654. [Google Scholar] [CrossRef]
- Chang, J.C.; Wu, M.C.; Liu, I.M.; Cheng, J.T. Increase of insulin sensitivity by stevioside in fructose-rich chow-fed rats. Horm. Metab. Res. 2005, 37, 610–616. [Google Scholar] [CrossRef]
- Xi, L.; Qian, Z.; Xu, G.; Zheng, S.G.; Sun, S.; Wen, N.; Sheng, L.; Shi, Y.; Zhang, Y.B. Beneficial impact of crocetin, a carotenoid from saffron, on insulin sensitivity in fructose-fed rats. J. Nutr. Biochem. 2007, 18, 64–72. [Google Scholar] [CrossRef]
- Jin, X.; Amitani, K.; Zamami, Y.; Takatoria, S.; Hobarab, N.; Kawamura, N.; Hirata, T.; Wada, A.; Kitamurad, Y.; Kawasakia, H. Ameliorative effect of Eucommia ulmoides Oliv. leaves extract (ELE) on insulin resistance and abnormal perivascular innervation in fructose-drinking rats. J. Ethnopharmacol. 2010, 128, 672–678. [Google Scholar] [CrossRef]
- Dai, S.; Todd, M.E.; Lee, S.; McNeill, J.H. Fructose loading induces cardiovascular and metabolic changes in nondiabetic and diabetic rats. Can. J. Physiol. Pharmacol. 1994, 72, 771–778. [Google Scholar] [CrossRef]
- Wheatcroft, S.B.; Williams, I.L.; Shah, A.M.; Kearney, M.T. Pathophysiological implications of insulin resistance on vascular endothelial function. Diabet. Med. 2003, 20, 225–268. [Google Scholar] [CrossRef]
- Caballero, A.E. Endothelial dysfunction in obesity and insulin resistance: A road to diabetes and heart disease. Obes. Res. 2003, 11, 1278–1289. [Google Scholar] [CrossRef]
- Worku, A.; Mahmood, S.M. Vascular reactivity changes in glucose-intolerant rat. J. Cardiovasc. Pharmacol. 2007, 50, 590–597. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, S.G.; Wu, G.T. Effect of acarbose on FFA; IGF-1 and vascular disease in impaired glucose tolerance rats [Article in Chinese]. J. Tongji Univ. (Med. Sci.). 2009, 30, 32–35. [Google Scholar]
- Onuma, S.; Nakanishi, K. Superoxide dismustase mimetic tempol decreases blood pressure by increasing renal medullary blood flow in hyperinsulinemic-hypertensive rats. Metabolism 2004, 53, 1305–1308. [Google Scholar] [CrossRef]
- Shinozaki, K.; Kashiwagi, A.; Nishio, Y.; Okamura, T.; Yoshida, Y.; Masada, M.; Toda, N.; Kikkawa, R. Abnormal biopterin metabolism is a major cause of impaired endothelium-dependent relaxation through nitric oxide/O2- imbalance in insulin-resistant rat aorta. Diabetes 1999, 48, 2437–2445. [Google Scholar] [CrossRef]
- Katakam, P.V.; Ujhelyi, M.R.; Hoenig, M.E.; Miller, A.W. Endothelialdysfunction precedes hypertension in diet-induced insulin resistance. Am. J. Physiol. 1998, 275, R788–R792. [Google Scholar]
- Al-Awwadi, N.A.; Bornet, A.; Azay, J.; Araiz, C.; Delbosc, S.; Cristol, J.P.; Linck, N.; Cros, G.; Teissedre, P.L. Red wine polyphenols alone or in association with ethanol prevent hypertension; cardiac hypertrophy; and production of reactive oxygen species in the insulin-resistant fructose-fed rat. J. Agr. Food Chem. 2004, 52, 5593–5597. [Google Scholar] [CrossRef]
- Thirunavukkarasu, V.; Anuradha, C.V. Influence of alpha-lipoic acid on lipid peroxidation and antioxidant defence system in blood of insulin-resistant rats. Diabetes Obes. Metab. 2004, 6, 200–207. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, W.D.; Liu, R.H.; Wang, J.; Chen, H. Effect of astragaloside IV on gene expression profile of rat myocardium [Article in Chinese]. Chin. J. Chin. Mater. Med. 2008, 33, 36–40. [Google Scholar]
- Sample Availability: Samples of the compounds are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, N.; Wang, X.-H.; Mao, S.-L.; Zhao, F. Astragaloside IV Improves Metabolic Syndrome and Endothelium Dysfunction in Fructose-Fed Rats. Molecules 2011, 16, 3896-3907. https://doi.org/10.3390/molecules16053896
Zhang N, Wang X-H, Mao S-L, Zhao F. Astragaloside IV Improves Metabolic Syndrome and Endothelium Dysfunction in Fructose-Fed Rats. Molecules. 2011; 16(5):3896-3907. https://doi.org/10.3390/molecules16053896
Chicago/Turabian StyleZhang, Ning, Xu-Hui Wang, Shi-Long Mao, and Feng Zhao. 2011. "Astragaloside IV Improves Metabolic Syndrome and Endothelium Dysfunction in Fructose-Fed Rats" Molecules 16, no. 5: 3896-3907. https://doi.org/10.3390/molecules16053896