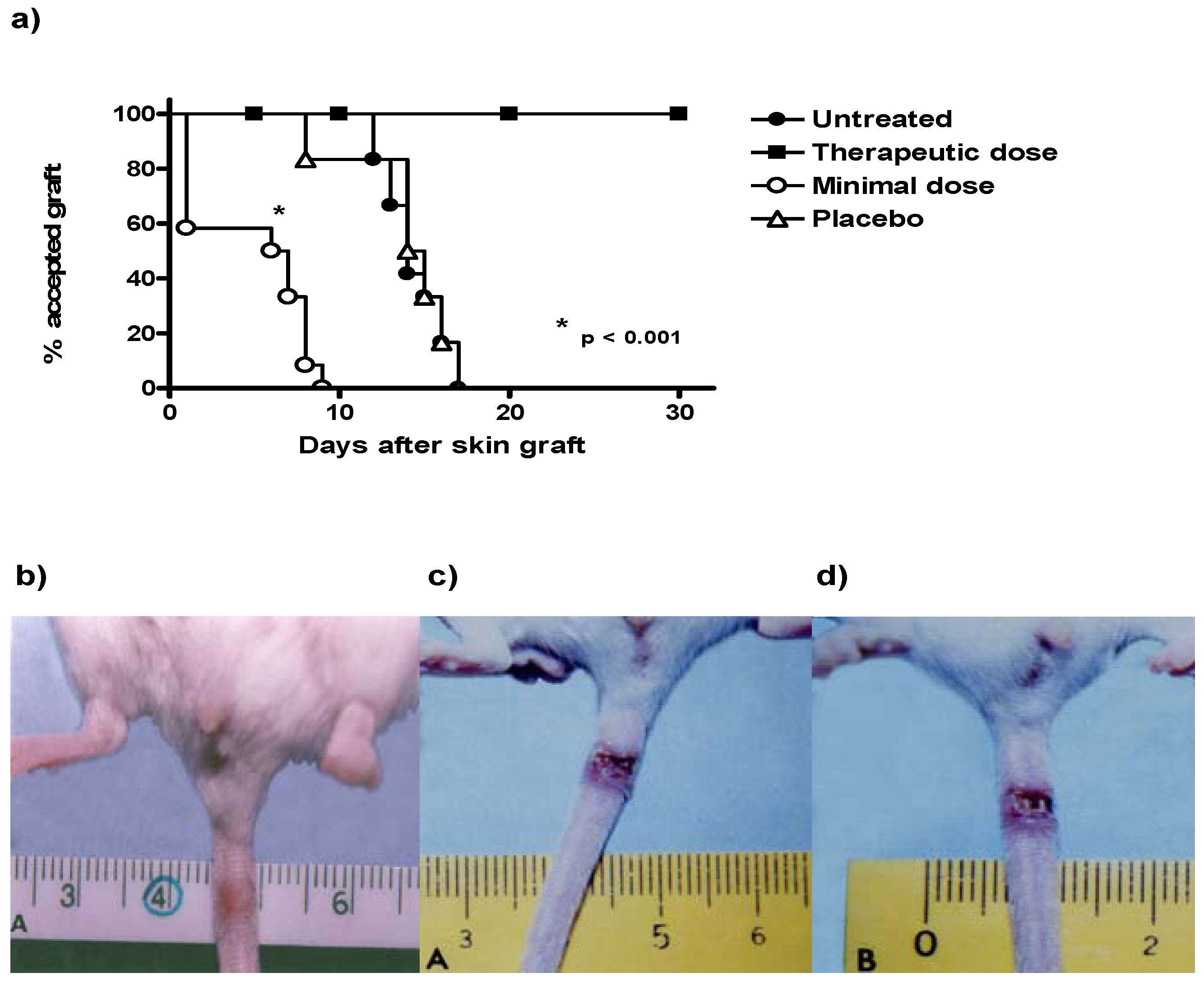
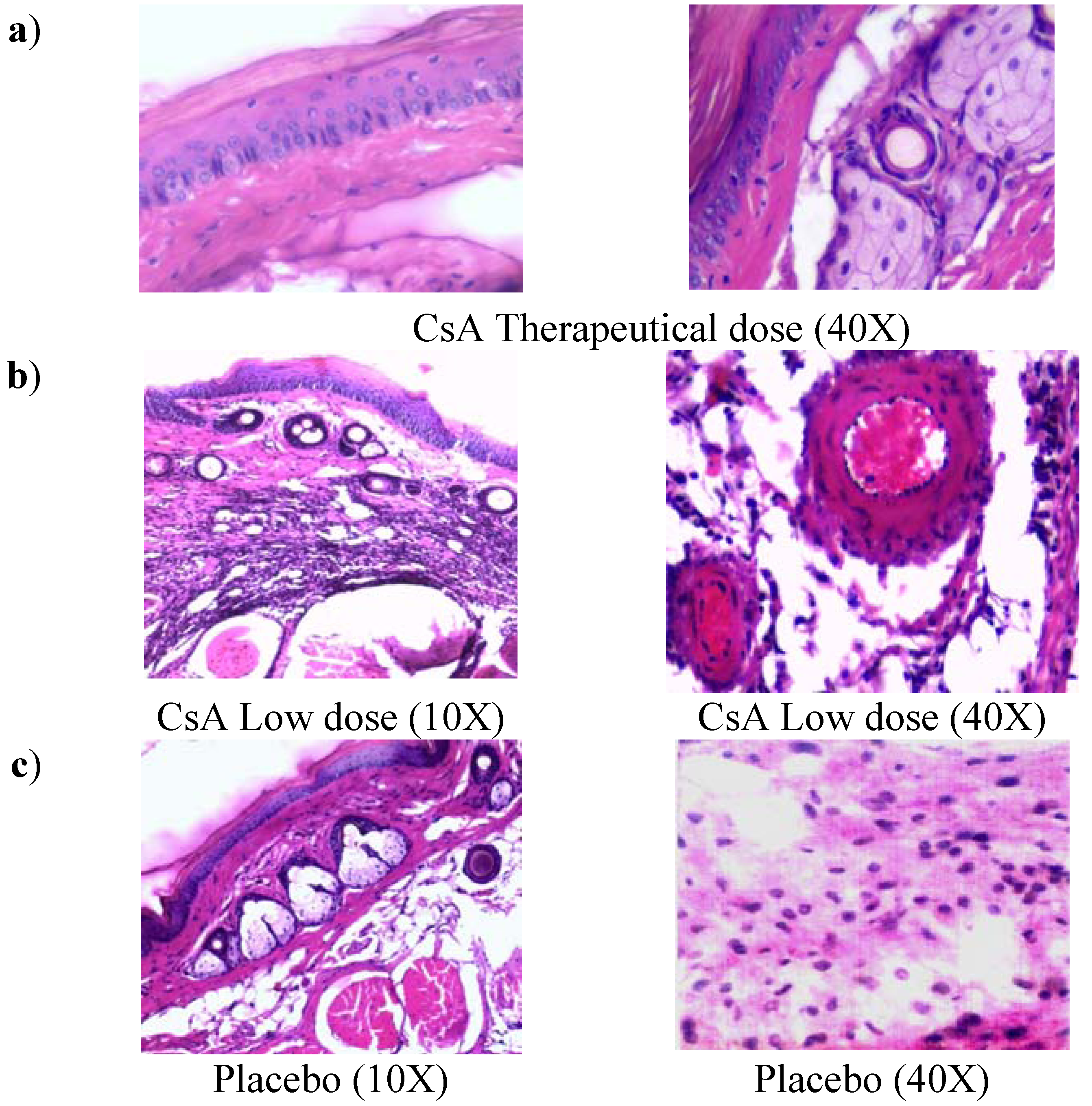
2.2. Histopathology Analysis of Skin Allograft Shows Acute Rejection in Mice Given Low Doses of CsA
Control mice with transplants but no treatment showed some inflammatory necrosis and little infiltration of macrophages and neutrophils at day 14 after the graft (
Figure 2a). In contrast, treatment with low doses of CsA induced a strong rejection reaction on the first days of treatment against the implanted dermis and epidermis, in which a severe neutrophil infiltration can be observed, with some macrophages. There were also some necrotic areas around a couple of arteries (
Figure 2b, 40X) and in some muscle fibers. In contrast with all other groups, the graft-receiving host dermis showed intense macrophage and neutrophil phagocytic activity against muscular and nerve tissue, which may stimulate degenerative processes in the epidermis and glands. In contrast, allografts in mice that received a therapeutic treatment of CsA did not show the above-mentioned reactions (data not shown). In mice given therapeutic doses of CsA, images of a graft that was incorporated into host tissues showed a region of skin alongside the graft with intact sebaceous gland (data not shown and
Figure 2b). The mice treated with the placebo formulation of therapeutic and minimal low doses of CsA solvents induced transplant rejection with characteristics similar to the control group with transplant and no treatment (placebo).
Figure 2.
Histology of grafted skin. a) Skin histology of therapeutic CsA-treated group, where graft was incorporated in the host tissues, the dermis presents abundant phagocytic cells but not necrosis, some intact sebaceous glands can be observed. b) Skin histology of grafted mice under consecutive low dose of CsA-treatment, where epidermis and dermis were found necrotic. Cell infiltrate with abundant neutrophils together with atrophic muscular beams were found. c) Image of grafted skin from mice treated with placebo. All photos were taken 7 days after skin allograft.
Figure 2.
Histology of grafted skin. a) Skin histology of therapeutic CsA-treated group, where graft was incorporated in the host tissues, the dermis presents abundant phagocytic cells but not necrosis, some intact sebaceous glands can be observed. b) Skin histology of grafted mice under consecutive low dose of CsA-treatment, where epidermis and dermis were found necrotic. Cell infiltrate with abundant neutrophils together with atrophic muscular beams were found. c) Image of grafted skin from mice treated with placebo. All photos were taken 7 days after skin allograft.
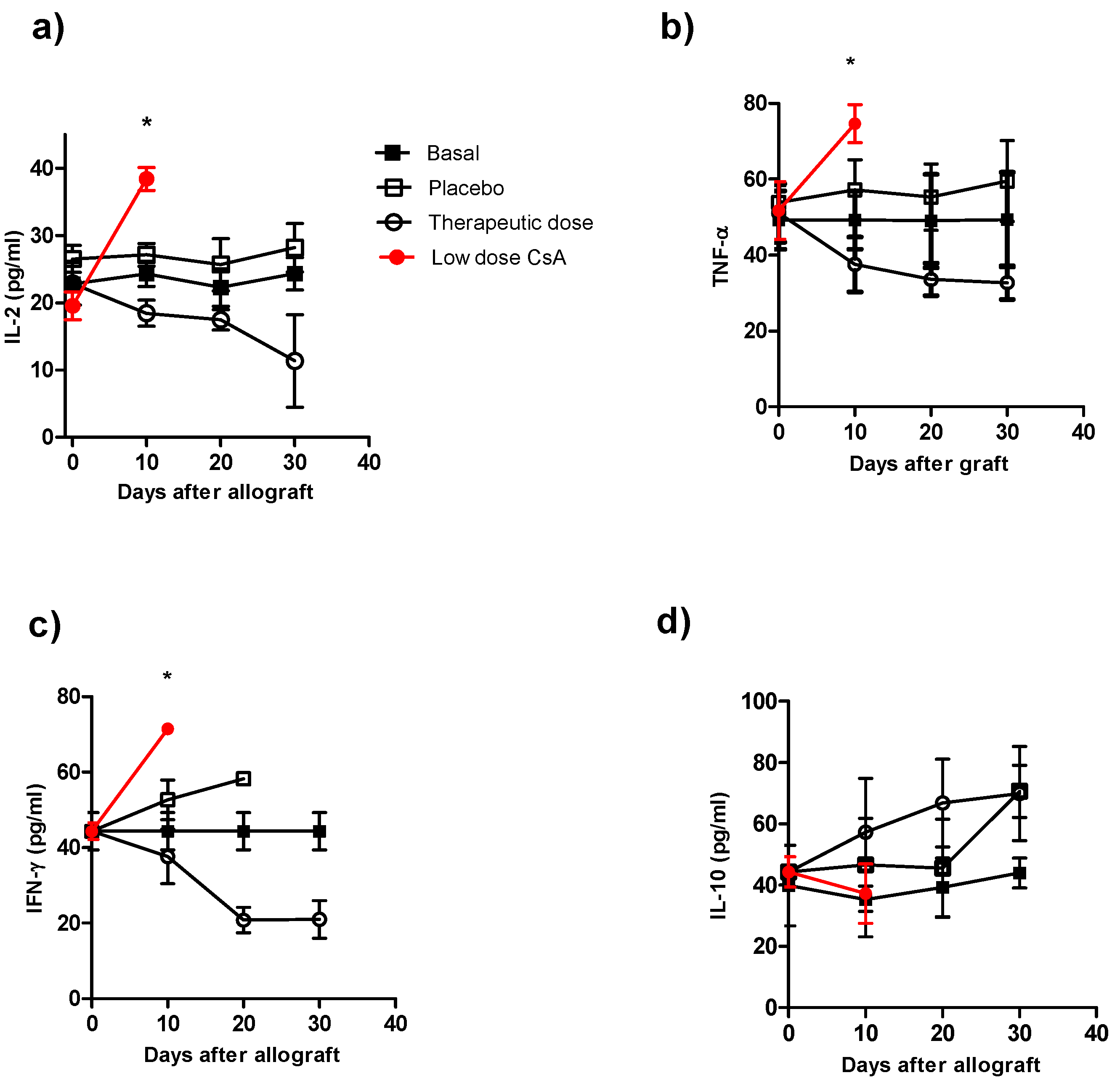
2.3. Mice Receiving Consecutive Minimal Doses of CsA Display a Pro-inflammatory Cytokine Profile
To correlate our findings on graft rejection with the cytokine profile in the different groups, we analyzed the serum levels of IL-2, IL-12, IFN-γ, TNF-α and IL-10. The control groups (animals with transplants without treatment and the control group plus placebo), which displayed normal rejection conditions, produced higher levels of IL-2 (
Figure 3a), TNF-α (
Figure 3b) and IFN-γ (
Figure 3c) early after skin allograft than the naïve group; however, these levels were lower than those in mice that received the minimal dose of CsA. This latter group produced the highest levels of pro-inflammatory cytokines (
Figure 3a–c). In contrast, the production of IL-2, TNF-α and IFN-γ was lower in mice given the therapeutic CsA treatment (
Figure 3a–c). Interestingly, no differences in IL-10 production were detected among the groups (
Figure 3d). The animals that received the therapeutic doses showed the lowest levels of IL-2, IFN-γ and TNF-α compared with all of the other groups but similar levels of IL-10 (
Figure 3a–d). The cytokine levels in mice receiving low doses of CsA were not followed after 10 days because by that time 100% of these mice displayed graft-rejection.
Figure 3.
Kinetics of cytokine production during course of allograft in ICR mice receiving different treatment of CsA. Individual mice were bleeding at different times after skin allograft, and sera were processed for cytokine detection by ELISA a) IL-2 detection, b) TNF-α detection, c) IFN-γ detection and d) IL-10 detection in sera of from animals receiving allografts and CsA treatment. The data are the mean of two independent experiments at each time point. An asterisk indicates statistically significant (*P < 0.05) differences between the groups receiving CsA with respect to basal and placebo.
Figure 3.
Kinetics of cytokine production during course of allograft in ICR mice receiving different treatment of CsA. Individual mice were bleeding at different times after skin allograft, and sera were processed for cytokine detection by ELISA a) IL-2 detection, b) TNF-α detection, c) IFN-γ detection and d) IL-10 detection in sera of from animals receiving allografts and CsA treatment. The data are the mean of two independent experiments at each time point. An asterisk indicates statistically significant (*P < 0.05) differences between the groups receiving CsA with respect to basal and placebo.
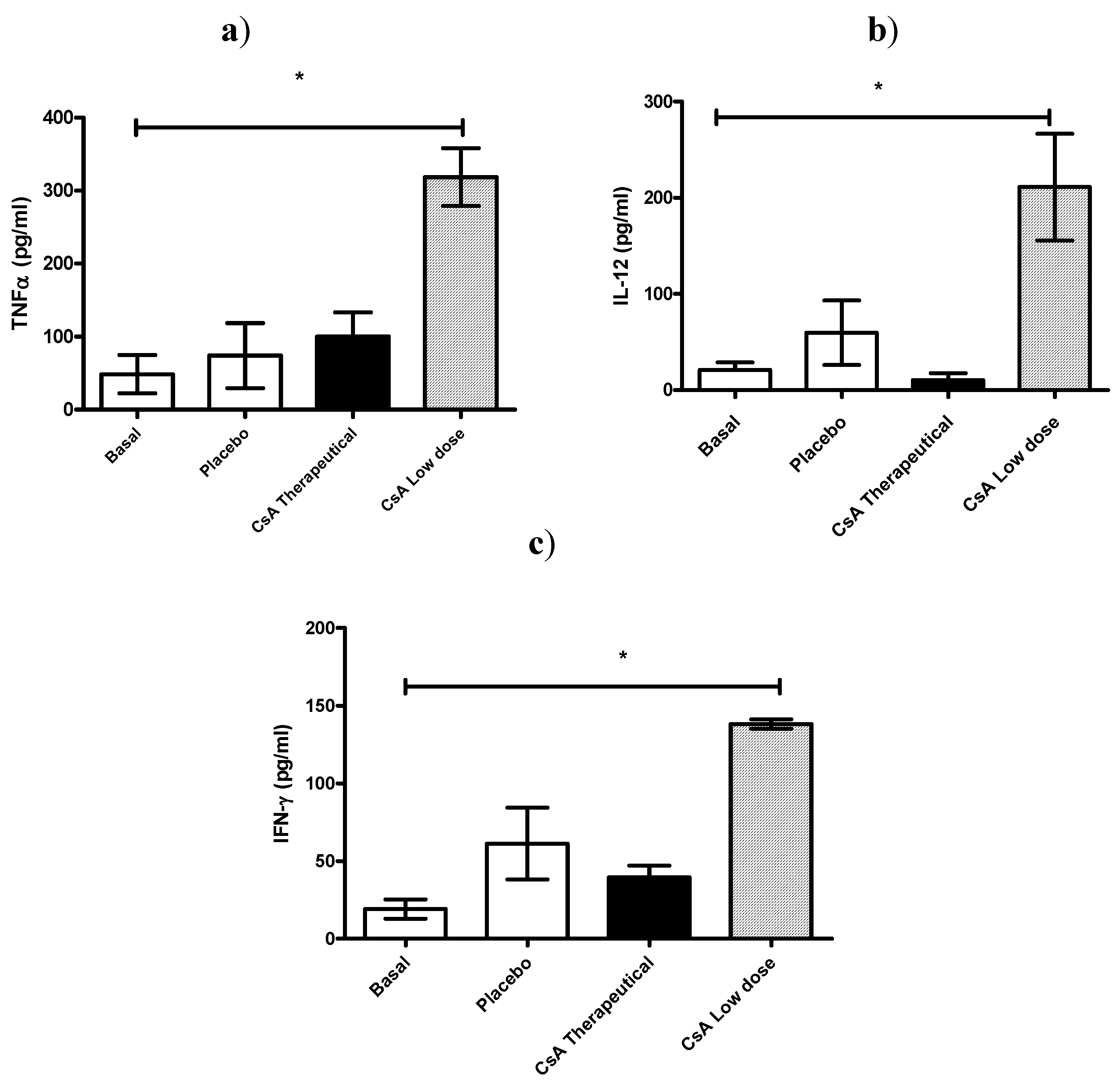
In another set of experiments, all mice were killed 7 days after skin allograft, and sera was analyzed for IL-12, TNF-α and IFN-γ production. As can be observed in
Figure 4 all these pro-inflammatory cytokines were significantly elevated in mice receiving daily low dose of CsA and which already had rejected the graft (
Figure 4 a–c).
Figure 4.
Effect of CsA on pro-inflammatory cytokine levels on sera early after rejection. (a–c) ICR mice, 6 mice per group, received placebo or 15 mg/kg of CsA (therapeutic dose) or 5 × 10−55 mg/kg of CsA (low dose) daily since the time of skin transplantation. Seven days later cytokines (TNF-α, IFN-γ and IL-12) were analyzed by ELISA.
Figure 4.
Effect of CsA on pro-inflammatory cytokine levels on sera early after rejection. (a–c) ICR mice, 6 mice per group, received placebo or 15 mg/kg of CsA (therapeutic dose) or 5 × 10−55 mg/kg of CsA (low dose) daily since the time of skin transplantation. Seven days later cytokines (TNF-α, IFN-γ and IL-12) were analyzed by ELISA.
The contradictory effects of very low and consecutive doses of CsA can be explained in part by the early observations of Bretscher and Havele [
10], who found that incremental doses of CsA to mice reconstituted with 5 × 10
6 spleen cells progressively inhibited DTH responses. The cell-mediated response occurred first, but was subsequently overtaken by the humoral response, as demonstrated when chicken erythrocytes were used as the antigen [
10]. However, when incremental CsA doses were given to mice reconstituted with 35 × 10
6 spleen cells, there was an initial antibody reaction that was gradually inhibited while the DTH reaction simultaneously increased, which was also shown using chicken erythrocytes as the antigen [
10].
These findings [
10] may suggest different immunological targets depending of the dose of CsA used. In fact, a recent report has indicated that CsA reduced IL-2 production and DTH reactions in BALB/c mice, but not in B6D2F1 mice, which suggest that
in vivo there are different responses to similar doses of CsA [
12]. Here we observed that mice treated with consecutive minimal doses of CsA exhibited spontaneous premature rejection of the transplants after an average of five days, which was associated with an increase in the levels of IL-2, IL-12, IFN-γ and TNF-α but without affecting IL-10.
It is well-known that CsA downregulates transcription of the IL-2 gene. As a result the main target of this agent is the T helper cell. In contrast, CsA exhibited minimal toxicity on preformed cytotoxic/suppressor T cells, B lymphocytes, granulocytes and macrophages [
13,
14,
15,
16,
17,
18]. This drug is a very potent and relatively selective inhibitor of T cell responses, particularly those responsible for graft rejections [
19,
20]. However, relatively recent observations also suggest that CsA may act differentially
in vivo affecting distinct cell populations, for example it has been demonstrated that the same dose of CsA can suppress
in vivo both NKT and dendritic cell maturation, but not NK cells in response to alpha-galactosylceramide [
21].
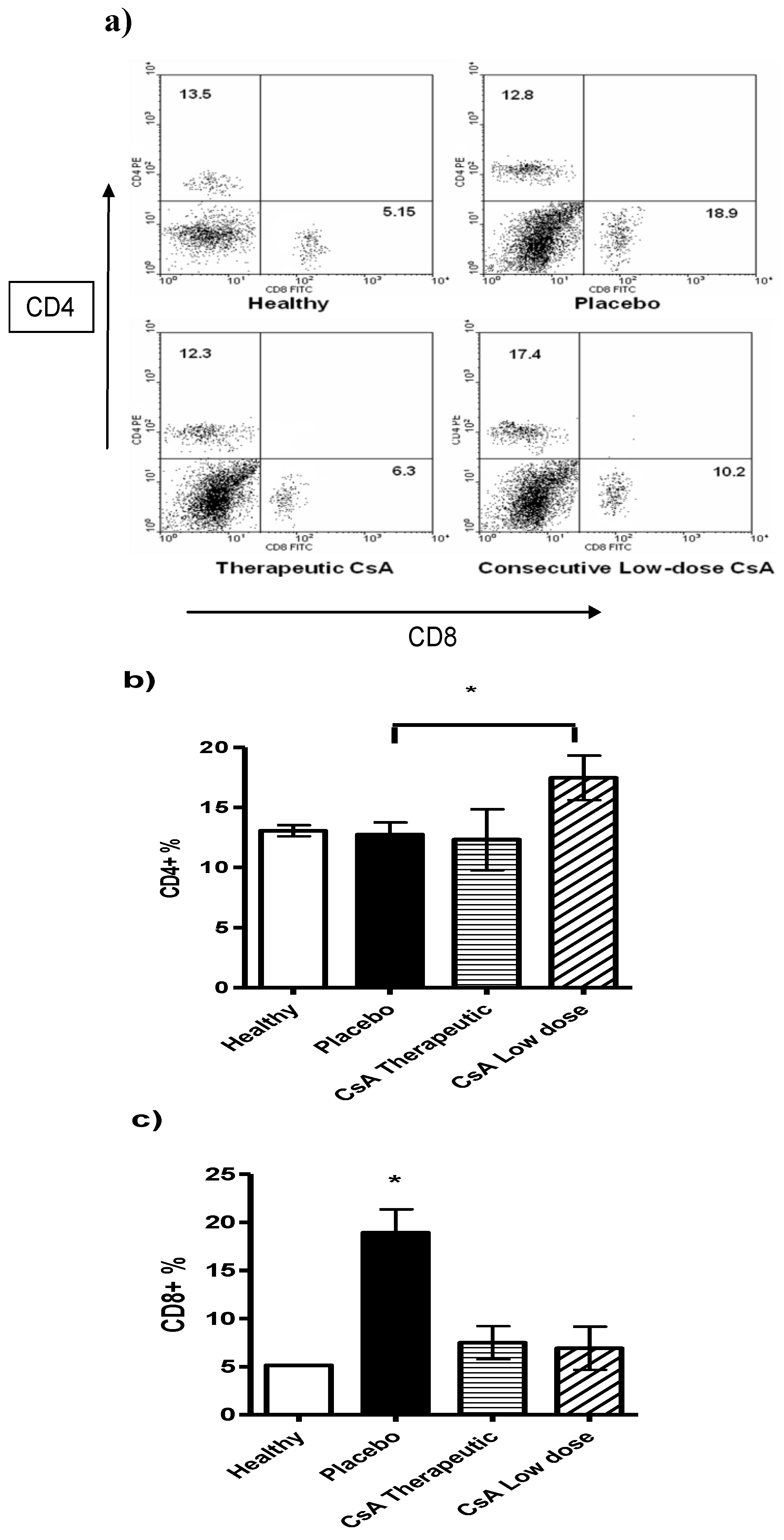
2.4. Minimal Doses of CsA Increase the CD4+ T Cell Subpopulation
Following the above idea we analyzed different lymphocytic populations such as CD8
+ and CD4
+ T cells in the spleens of mice with skin allograft that received the different treatments. Mice with skin allograft that received the placebo displayed a significant increase in the percentage of the CD8
+ T-cell subpopulation compared with naïve mice (
Figure 5). In contrast, animals given minimal doses of CsA showed significantly higher percentages of CD4
+ T cells in the spleens than mice given the therapeutic doses or the placebo. Interestingly, the placebo group also displayed more CD8
+ T cells than the therapeutic treatment group, but the difference was not statistically significant. In contrast, mice that received high doses of CsA had significantly less CD8
+ T cells than those treated with the placebo (
Figure 5c).
Figure 5.
Flow cytometry analysis of spleen cells from mice grafted and differentially treated with CsA. Spleen cells were obtained after graft rejection or after finished the follow-up period of time. CD4 and CD8 positive cells were analyzed. a) Representative dot plot of spleen cells from naïve mice, placebo-treated mice, therapeutic dose of CsA and consecutive low dose of CsA. Numbers indicate mean percentage of positive cells. b) Average of percentage of CD4+ cells and c) percentage of CD8+ cells. Bars show average ± SD, n = 4 mice per group; * P < 0.05.
Figure 5.
Flow cytometry analysis of spleen cells from mice grafted and differentially treated with CsA. Spleen cells were obtained after graft rejection or after finished the follow-up period of time. CD4 and CD8 positive cells were analyzed. a) Representative dot plot of spleen cells from naïve mice, placebo-treated mice, therapeutic dose of CsA and consecutive low dose of CsA. Numbers indicate mean percentage of positive cells. b) Average of percentage of CD4+ cells and c) percentage of CD8+ cells. Bars show average ± SD, n = 4 mice per group; * P < 0.05.
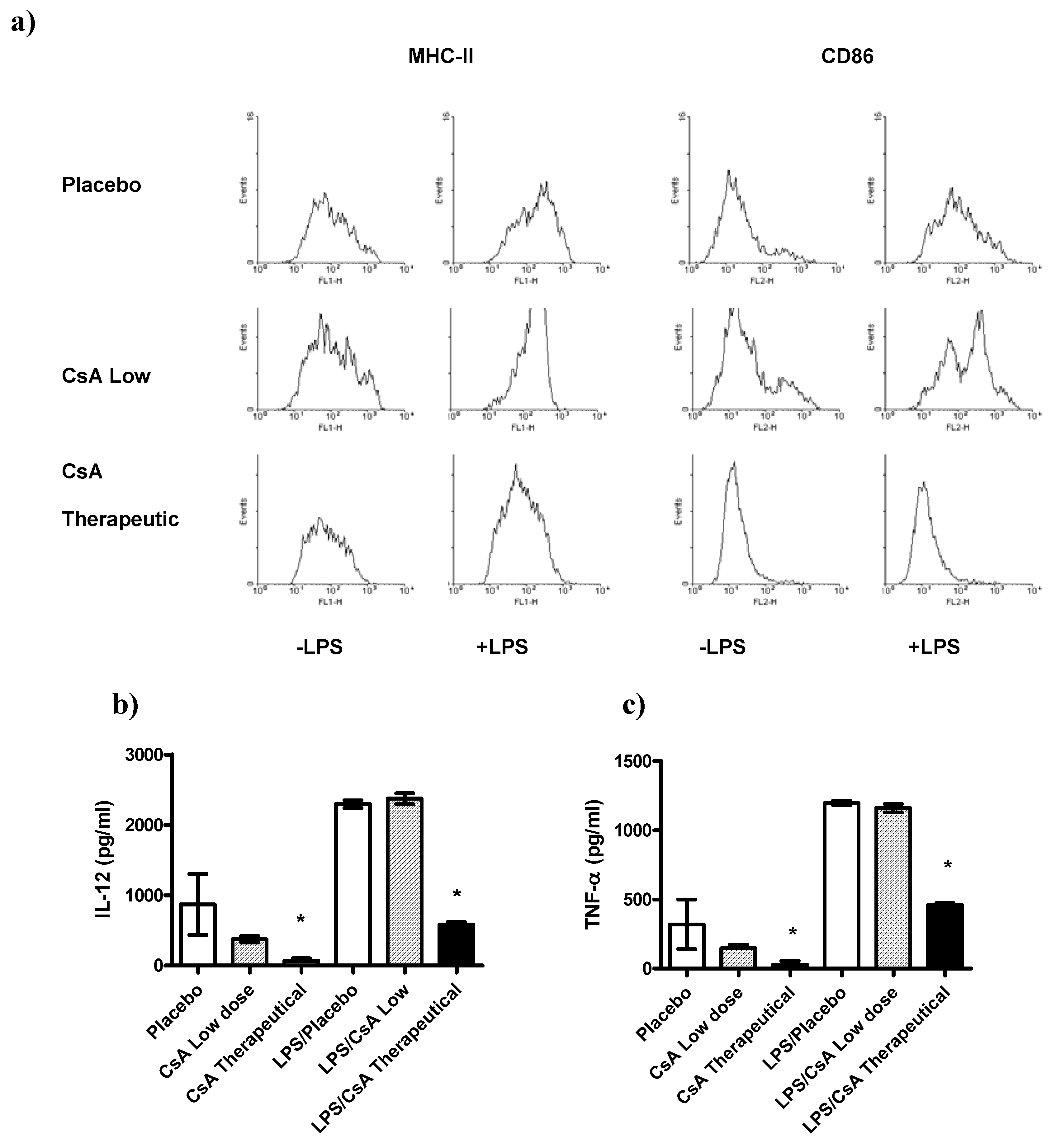
2.5. CsA May Affect Other Immune Cell Populations
Recently it has been demonstrated that other cell types besides T cells can be affected by CsA [
21,
22,
23]. In order to know whether or not other important regulatory cells were affected by the low doses of CsA treatment we analyzed whether exposure of DCs to therapeutic or low doses of CsA
in vitro may alter the pro-inflammatory response and maturation markers of those cells to LPS-mediated stimuli. The maturation markers (MHC-II and CD86) as well as the production of IL-12, and TNF-α two key cytokines associated with inflammatory responses, were assayed. We detected that DCs exposed to therapeutic dose of CsA inhibited the expression of the maturation markers MHC-II and CD86 (
Figure 6a), in contrast, DCs exposed to low dose of CsA displayed a typical maturation, similar to that obtained with placebo (
Figure 6a). In line with these observations, exposure of DCs to placebo or CsA low dose for 24 h did not induce significant secretion of IL-12 or TNF-α compared with unstimulated DCs (
Figure 6b–c). In contrast, DCs exposure to therapeutic CsA dose showed lower levels of these two inflammatory cytokines (
Figure 6b–c). As expected, we found that BMDCs stimulated with LPS (1 μg/mL), induced high production of the pro-inflammatory cytokines IL-12 and TNF-α (
Figure 6b–c). We then analyzed whether acute exposure of DCs to different doses of CsA may influence the response to LPS-mediated stimuli. BMDCs were exposed to CsA and immediately stimulated with LPS for 24 h. Exposure to therapeutic dose of CsA down-regulated the production of both pro-inflammatory cytokines tested (
Figure 6b–c). Interestingly, CsA in low dose did not alter the production of both IL-12 and TNF-α (
Figure 6b–c).
DCs are also important in modulating early responses, here we found that DCs were highly sensitive to therapeutical dose of CsA given that DCs exposed
in vitro to such amount of CsA were unable to maturate in response to LPS and also showed a significant low production of inflammatory cytokines, in contrast, exposure to low dose of CsA did not affect either maturation or cytokine production of DCs. These data are in line with those reported for the inhibition of DC maturation
in vivo using high doses of CsA [
24]. We also analyzed other cells such as NK cells which were unaltered (data not shown).
Figure 6.
Differential effect of CsA doses on dendritic cells (DC) maturation and activation. DCs were developed from bone marrow of healthy mice, and culture with GM-CSF for 5 days, then DCs were stimulated or not with LPS 1 μg/mL in presence of therapeutic or low dose of CsA or placebo. a) Low dose of CsA do not alter DC maturation, but therapeutic dose of CsA significantly inhibits DC expression of MHC-II and CD86. b-c) Twenty four hours later supernatants were collected and analyzed for IL-12 and TNF-α production by ELISA. * p < 0.05 compared with placebo. Data represent 2 independent experiments.
Figure 6.
Differential effect of CsA doses on dendritic cells (DC) maturation and activation. DCs were developed from bone marrow of healthy mice, and culture with GM-CSF for 5 days, then DCs were stimulated or not with LPS 1 μg/mL in presence of therapeutic or low dose of CsA or placebo. a) Low dose of CsA do not alter DC maturation, but therapeutic dose of CsA significantly inhibits DC expression of MHC-II and CD86. b-c) Twenty four hours later supernatants were collected and analyzed for IL-12 and TNF-α production by ELISA. * p < 0.05 compared with placebo. Data represent 2 independent experiments.
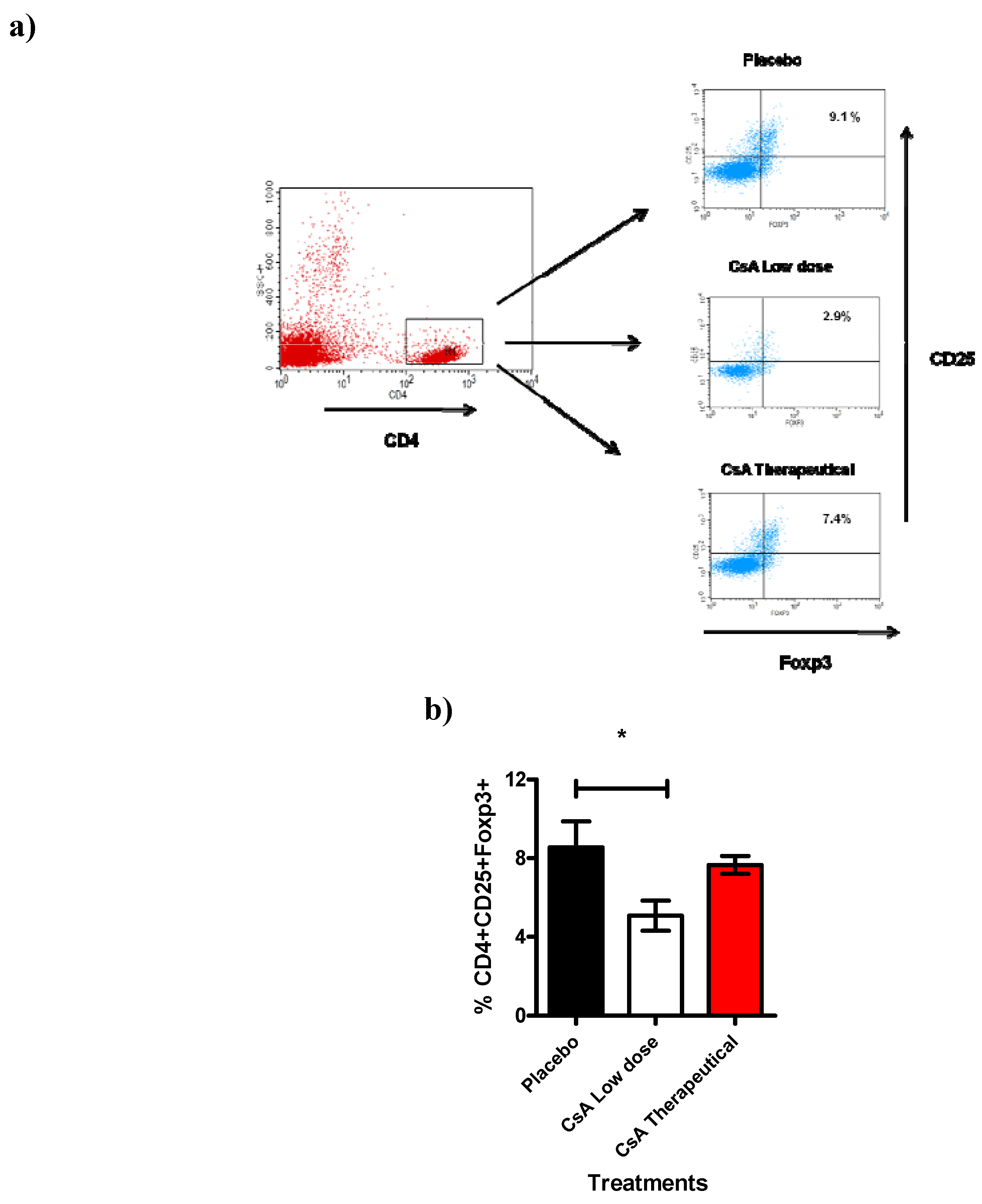
Next we analyzed another cell population also involved in transplantation such as the CD4
+CD25
+ T reg cells [
26]. Here we studied the total spleen cells 7 days after allograft. Spleens were removed and analyzed for the expression of CD4
+CD25
hiFoxp3
+ cells. Interestingly, we found a significant reduction (approx. 30 to 40%) of cells expressing CD4
+CD25
hiFoxp3
+ in mice that were treated with consecutive low dose of CsA during 7 days and that had already rejected the skin allograft (
Figure 7), treatment with placebo or therapeutic dose of CsA were similar and displayed higher percentages of Tregs. This intriguing result may support our data regarding to higher levels of inflammatory cytokines as well as the faster graft rejection, probably because the limited regulatory activity of the remaining Treg cells.
Figure 7.
Treg cell population is affected by Low dose of CsA. This figure shows the flow cytometric analysis of T regs population from the spleen. a) Representative dotplot indicating the percentage of CD25+Foxp3+ cells obtained from the gate of CD4+ cells from spleen of grafted mice under different treatments (7 days after graft). b) Analysis of total Tregs, data are representative of two independent experiments, bars indicate standard deviation, n = 4 mice per group. *P < 0.02 according to one way ANOVA.
Figure 7.
Treg cell population is affected by Low dose of CsA. This figure shows the flow cytometric analysis of T regs population from the spleen. a) Representative dotplot indicating the percentage of CD25+Foxp3+ cells obtained from the gate of CD4+ cells from spleen of grafted mice under different treatments (7 days after graft). b) Analysis of total Tregs, data are representative of two independent experiments, bars indicate standard deviation, n = 4 mice per group. *P < 0.02 according to one way ANOVA.
In different experimental transplant models, classical alloreactive CD4
+ or CD8
+ T cells along with specialized alloantibodies regulate allograft rejection reactions after transplantation. In addition, rejection may be inhibited by anti-CD4 or anti-CD8 antibodies or accelerated by depletion of CD4
+CD25
+ cells. These different immunoeffectors result in graft rejection through various mechanisms: (a) Alloreactive T cells may recruit and activate macrophages, initiating graft injury through a delayed-type hypersensitivity (DTH) reaction; (b) alloreactive T cytotoxic cells directly provoke lysis of endothelial and parenchymal cells of the graft; (c) antibodies attack the endothelial tissue, activate the adaptive immune system and cause injuries in the blood vessels of the graft and (d) the absence or lower percentages of CD4
+CD25
+ Foxp3
+ T regs may allow a more rapid and strong pro-inflammatory response that active and recruit innate cells such as neutrophils and macrophages [
6,
7,
8,
9]. In this study we found an association between the lower percentages of Tregs with faster graft rejection in mice receiving consecutive low dose of CsA which displayed stronger responses and 100% of allografts were rejected in less than ten days. This effect was also associated with neutrophil and macrophage accumulation and with higher levels of IL-2, IFN-γ and TNF-α.
The reported activation of the cell-mediated immune response by minimal doses of CsA may induce acute cellular rejection [
10], but this kind of rejection is characterized by necrosis of parenchymal cells and is often associated with infiltration of lymphocytes and macrophages. Here we observed mainly a severe neutrophil infiltration in the skin that was promptly rejected in mice treated with consecutive low dose of CsA, such important difference may involve a distinct mechanism of rejection.
Even thought that our results are surprising because the immunosuppressive effects of CsA have been attributed to the prevention of IL-2 transcription [
27], they are in line with a recent report by Miroux et al who found
in vitro that a low dose of CsA can inhibit T regulatory cells activity whereas a high dose had no effect in this T cell population [
23], interestingly here we observed a similar phenomena but
in vivo. It is well known that Treg cells can mediate the induction and maintenance of immunological self-tolerance as well as transplant tolerance, their inhibition by CsA might promote graft rejection [
28]. Thus, further research is guaranteed to evaluate whether or not consecutive low dose of CsA
in vivo can affect the activities of these subpopulations. Moreover, a recent report has indicated that
in vivo CsA reduced IL-2 production and DTH reactions in BALB/c mice but not in B6D2F1 mice, which suggest that
in vivo there are different responses to similar doses of CsA [
12]. Moreover, other drug such as cyclophosphamide has been used at low doses in immunotherapy for mesothelioma tumors where it was found that low doses of cyclophosphamide induced beneficial effects by preventing the increase of Treg cells [
25]. Consequently, minimal and consecutive doses of CsA deserve further and deeper research in order to, in a near future, may considered to stimulate innate immune responses in the treatment of certain illnesses, such as miliary tuberculosis, leprosy and some parasitic diseases [
29], given that CsA is already an authorized drug for human use. In fact, the puzzling aspect of the effect of different doses on the targets or on the complete individual is getting bigger relevance with other drugs such as rapamycin which depending of the dosage it can influence different pathways [
22] .

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






