Physico-Chemical Characteristics and Functional Properties of Chitin and Chitosan Produced by Mucor circinelloides Using Yam Bean as Substrate

Abstract
:1. Introduction
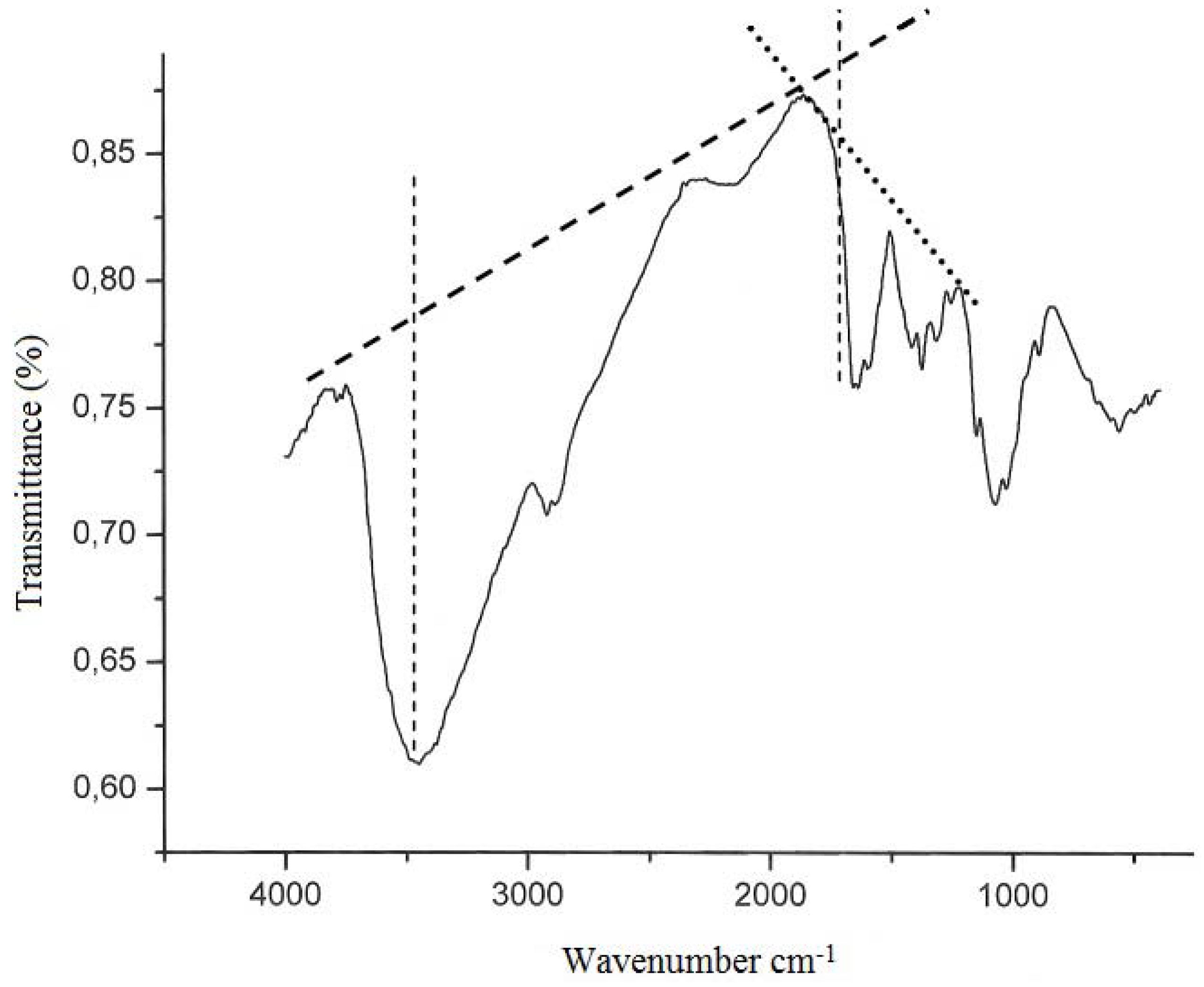
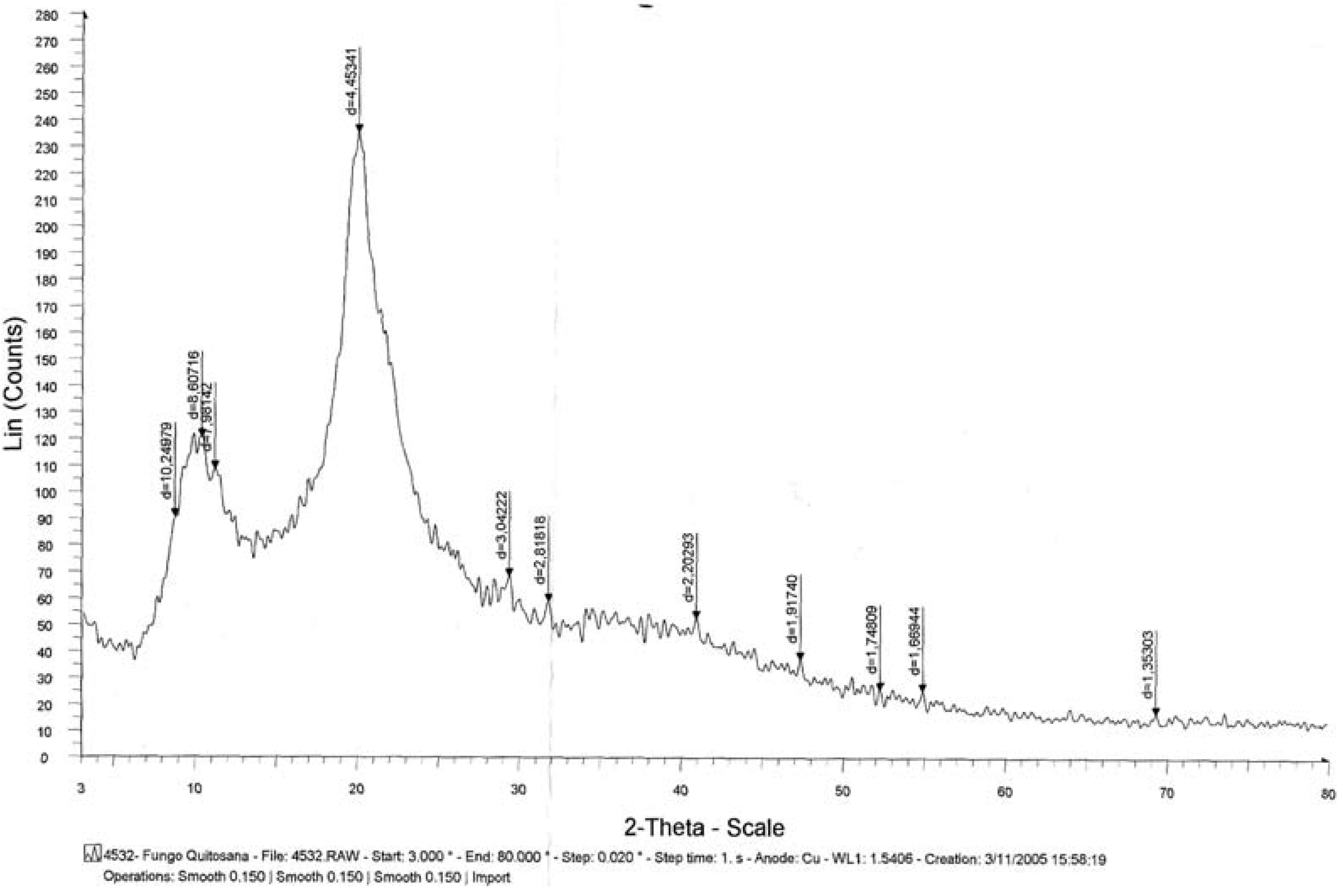
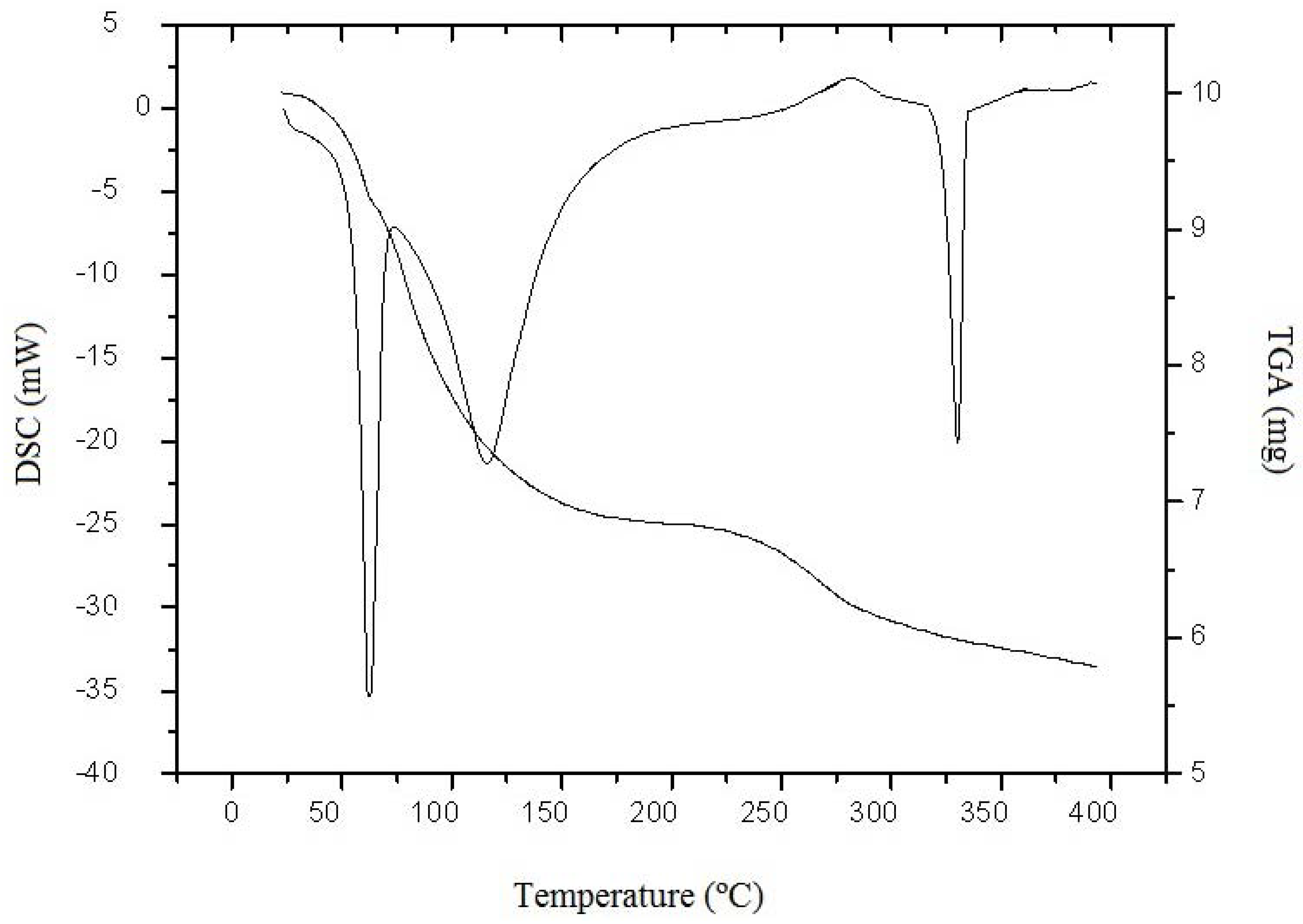
2. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
| M. circinelloides/Cultivation time (h) | Glucose consumption (g/L) | Nitrogen consumption (g/L) | pH | Biomass yield (g/L) | Chitin yield (mg/g) | Chitosan yield (mg/g) |
|---|---|---|---|---|---|---|
| 0 | 11.4 ± 0.2 | 8.72 ± 0.09 | 7 ± 0.1 | 0.0 ± 0.0 | 260 ± 0.4 | 33 ± 0.1 |
| 24 | 9.6 ± 0.1 | 7.2 ± 0.2 | 5.9 ± 0.2 | 11.6 ± 0.6 | 320 ± 0.2 | 64 ± 0.2 |
| 48 | 3.95 ± 0.1 | 1.4 ± 0.09 | 4.8 ± 0.2 | 16.8 ± 0.4 | 500 ± 0.2 | 64 ± 0.2 |
| 72 | 3.16 ± 0.1 | 0.84 ± 0.1 | 4.5 ± 0.1 | 20.7 ± 0.5 | 500 ± 0.2 | 64 ± 0.4 |
| 96 | 2.08 ± 0.1 | 0.01 ± 0.07 | 4.2 ± 0.1 | 18.9 ± 0.5 | 500 ± 0.0 | 64 ± 0.1 |
| Microorganisms | Substrate | Biomass (g.L−1) | Chitin (mg.g−1) | Chitosan (mg.g−1) | Reference |
|---|---|---|---|---|---|
| Mucor circinelloides | Yam bean | 20.70 | 500 | 64 | This study |
| Cunninghamella elegans | Yam bean | 24.30 | 440 | 66 | [25] |
| Cunninghamella elegans | Hesseltine and Anderson with added 5%NaCl and 6%glucose | 24.40 | 388 | 70 | [10] |
| Cunninghamella bertholletiae | Sugar cane juice | 7.70 | - | 128 | [14] |
| Aspergillus niger | Potato Dextrose Broth | 9.00 | - | 107 | [16] |
| Lentinus edodes | Potato Dextrose Broth | 1.4 | - | 33 | [16] |
| Zygosaccharomyces rouxii | Yeast Malt Extract Broth | 4.4 | - | 36 | [16] |
| Candida albicans | Yeast Malt Extract Broth | 1.8 | - | 44 | [16] |



3. Experimental Section
3.1. Microorganism
3.2. Fermentation Medium Peparation
3.3. Culture Growth Conditions
3.4. Glucose and Nitrogen Consumption and pH Determination
3.5. Chitin and Chitosan Extraction
3.6. Chitin and Chitosan Characterization
3.6.1. Infrared Spectroscopy (Deacetylation degree–DD%)
3.6.2. Molecular Weight
3.6.3. Thermal Analysis
3.6.4. X-ray Diffraction
3.7. Statistical Analysis
4. Conclusions
Acknowledgments
References
- Amorim, R.V.S.; Ledingham, W.M.; Fukushima, E.K.; Campos-Takaki, G.M. Screening of chitin deacetylase from Mucoralean strains (Zygomycetes) and its relationship to cell growth rate. J. Ind. Microbiol. Biotechnol. 2005, 31, 19–23. [Google Scholar]
- Andrade, V.S.; Neto, B.B.; Souza, W.; Campos-Takaki, G.M. A factorial designs analysis of chitin production by Cunninghamella elegans. Can. J. Microbiol. 2000, 46, 1042–1045. [Google Scholar]
- Campos-Takaki, G.M. The fungal versatility on the copolymers chitin and chitosan production. In Chitin and Chitosan Opportunities and Challenges; Dutta, P.K., Ed.; SSM International Publication: Midnapore, India, 2005; pp. 69–94. [Google Scholar]
- Tharanathan, R.N.; Kittur, F.S. Chitin-the undisputed biomolecule of great potential. Crit. Rev. Food Sci. Nutr. 2003, 43, 61–87. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Aider, M. Chitosan application for active bio-based films production and potential in the food industry: Review. LWT – Food Sci. Technol. 2010, 43, 837–842. [Google Scholar] [CrossRef]
- Chatterjee, S.; Adhya, M.; Guah, A.K.; Chatterjee, B.P. Chitosan from Mucor rouxii: Production and physico-chemical characterization. Process Biochem. 2005, 40, 395–400. [Google Scholar] [CrossRef]
- Santos, J.E.; Soares, J.P.; Dockal, E.R.; Campana Filho, S.P.; Cavalheiro, E.T.G. Caracterização de quitosanas comerciais de diferentes origens. Polím. Ciência e Tecnol. 2003, 13, 242–249. [Google Scholar]
- Silva, M.C.F.; Barros Neto, B.; Stamford, T.C.M.; Campos-Takaki, G.M. Effect of environmental conditions on chitin and chitosan production by Cunninghamella elegans UCP 542 using factorial design. Asian Chitin J. 2007, 3, 15–22. [Google Scholar]
- Silva, M.C.F.; Stamford, T.C.M.; Franco, L.O.; Campos-Takaki, G.M. Effects of salinity and glucose on chitin and chitosan production by Cunninghamella elegans. Asian Chitin J. 2006, 2, 29–38. [Google Scholar]
- Nadarajah, K.; Kader, J.; Mohd, M.; Paul, D.C. Production of chitosan by fungi. J. Biol. Sci. 2001, 4, 263–265. [Google Scholar]
- Nwe, N.; Chandrkrachang, S.; Stevens, W.F.; Maw, T.; Tan, T.K.; Khor, E.; Xong, S.M. Production of fungal chitosan by solid state and submerged fermentation. Carbohydr. Polym. 2002, 49, 235–237. [Google Scholar] [CrossRef]
- Tajdinia, F.; Ali Aminia, M.; Nafissi-Varchehc, N.; Faramarzi, M.A. Production, physiochemical and antimicrobial properties of fungal chitosan from Rhizomucor miehei and Mucor racemosus. Int. J. Biol. Macromol. 2010, 47, 180–183. [Google Scholar] [CrossRef]
- Amorim, R.V.S.; Pedrosa, R.P.; Kazutaka, F.; Martínez, C.R.; Ledingham, W.M.; Campos-Takaki, G.M. Alternative carbon sources from sugar cane process for submerged cultivation of Cunninghamella bertholletiae to produce chitosan. Food Technol. Biotechnol. 2006, 44, 519–523. [Google Scholar]
- Bento, R.B.; Stamford, T.L.M.; Stamford, T.C.M.; Andrade, S.A.C.; Souza, E.L. Sensory evaluation and inhibition of Listeria monocytogenes in bovine pâté added of chitosan from Mucor rouxii. LWT – Food Sci. Technol. 2011, 44, 588–591. [Google Scholar] [CrossRef]
- Pochanavanich, P.; Suntornsuk, W. Fungal chitosan production and its characterization. Lett. Appl. Microbiol. 2002, 35, 17–21. [Google Scholar] [CrossRef]
- Tan, S.C.; Tan, T.K.; Wong, S.M.; Khor, E. The chitosan yield of Zygomycetes at their optimum harvesting time. Carbohydr. Polym. 1996, 30, 239–242. [Google Scholar] [CrossRef]
- Synowiecki, J.; Al-Khatteb, N.A.A.Q. Mycelia of M. rouxii as a source of chitin and chitosan. Food Chem. 1997, 60, 605–610. [Google Scholar] [CrossRef]
- Maghsoodi, V.; Yaghmaei, S. Comparison of solid substrate and submerged fermentation for chitosan production by Aspergillus niger. Trans. C: Chem. Chem. Eng. 2010, 17, 153–157. [Google Scholar]
- Jaworska, M.; Sakurai, K.; Gaudon, P.G.E. Influence of chitosan characteristics on polymer: Crystallographic properties. Polym. Int. 2003, 52, 198–205. [Google Scholar] [CrossRef]
- Kittur, F.S.; Prashanth, K.V.H.; Sankar, K.V.; Tharanathan, R.N. Characterization of chitin, chitosan and their carboxymethyl derivatives by differential scanning calorimetry. Carbohydr. Polym. 2002, 49, 185–193. [Google Scholar] [CrossRef]
- Martínez-Camacho, A.P.; Cortez-Rocha, M.O.; Ezquerra-Brauer, J.M.; Graciano-Verdugo, A.Z.; Rodriguez-Félix, F.; Castillo-Ortega, M.M.; Yépiz-Gómez, M.S.; Plascencia-Jatomea, M. Chitosan composite films: Thermal, structural, mechanical and antifungal properties. Carbohydr. Polym. 2010, 82, 305–315. [Google Scholar] [CrossRef]
- Andrade, V.S.; Neto, B.B.; Fukushima, K.; Campos-Takaki, G.M. Effect of medium components and time of cultivation on chitin production by Mucor circinelloides (Mucor javanicus IFO 4570) – A factorial study. Rev. Iberoam. Micol. 2003, 20, 149–153. [Google Scholar]
- Franco, L.O.; Stamford, T.C.M.; Stamford, N.P.; Campos-Takaki, G.M. Cunningamella elegans (IFM 46109) como fonte de quitina e quitosana. Rev. Anal. 2005, 14, 40–44. [Google Scholar]
- Stamford, T.C.M.; Stamford, T.L.M.; Stamford, N.P.; Neto, B.B.; Campos-Takaki, G.M. Growth of Cunninghamella elegans UCP 542 and production of chitin and chitosan using yam bean medium. Electron. J. Biotechnol. 2007, 10. Available online: http://www.ejbiotechnology.info/content/vol10/issue1/full/1/ (accessed on 16 April 2011).
- Franco, L.O.; Maia, R.C.G.; Porto, A.L.F.; Messias, A.S.; Fukushima, K.; Campos-Takaki, G.M. Heavy metal biosorption by chitin and chitosan isolated from Cunninghamella elegans IFM 46109. Braz. J. Microbiol. 2004, 35, 243–247. [Google Scholar] [CrossRef]
- Amorim, R.V.S.; Souza, W.; Fukushima, K.; Campos-Takaki, G.M. Faster chitosan production by mucoralean strains in submerged culture. Braz. J. Microbiol. 2001, 32, 20–23. [Google Scholar] [CrossRef]
- Sarangbin, S.; Watanapokasin, Y. Yam bean starch: A novel substrate for citric acid production by the protease-negative mutant strain of Aspergillus niger. Carbohydr. Polym. 1999, 38, 219–224. [Google Scholar] [CrossRef]
- Chung, Y.-C.; Su, Y.-A.; Chen, C.-C.; Jia, G.; Wang, H.-L.; Wu, J.C.G.; Lin, J.G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar]
- Shigemasa, Y.; Minami, S. Applications of chitin and chitosan for biomaterials. Biotechnol. Genet. Eng. Rev. 1996, 17, 383–420. [Google Scholar]
- Koev, S.T.; Dykstra, P.H.; Luo, X.; Rubloff, G.W.; Bentley, W.E.; Payneb, G.F.; Ghodssi, R. Chitosan: An integrative biomaterial for lab-on-a-chip devices. Lab Chip 2010, 10, 3026–3042. [Google Scholar] [CrossRef]
- Francis Suh, J.-K.; Matthew, H.W.T. Application of chitosan-based polysaccharide biomaterials in cartilage tissue engineering: A review. Biomaterials 2000, 21, 2589–2598. [Google Scholar] [CrossRef]
- Qin, C.Q.; Xiao, Q.; Li, H.R.; Fang, M.; Liu, Y.; Chen, X.; Li, Q. Calorimetric studies of the action of chitosan-N-2-hydroxypropyl trimethyl ammonium chloride on the growth of microorganisms. Int. J. Biol. Macromol. 2004, 34, 121–126. [Google Scholar] [CrossRef]
- Liu, H.; Du, Y.; Yang, J.; Zhu, H. Structural characterization and antimicrobial activity of chitosan/betaine derivative complex. Carbohydr. Polym. 2004, 55, 291–297. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Fai, A.E.C.; Stamford, T.C.M.; Stamford-Arnaud, T.M.; Santa-Cruz, P.D.; Silva, M.C.F.d.; Campos-Takaki, G.M.; Stamford, T.L.M. Physico-Chemical Characteristics and Functional Properties of Chitin and Chitosan Produced by Mucor circinelloides Using Yam Bean as Substrate. Molecules 2011, 16, 7143-7154. https://doi.org/10.3390/molecules16087143
Fai AEC, Stamford TCM, Stamford-Arnaud TM, Santa-Cruz PD, Silva MCFd, Campos-Takaki GM, Stamford TLM. Physico-Chemical Characteristics and Functional Properties of Chitin and Chitosan Produced by Mucor circinelloides Using Yam Bean as Substrate. Molecules. 2011; 16(8):7143-7154. https://doi.org/10.3390/molecules16087143
Chicago/Turabian StyleFai, Ana Elizabeth C., Thayza C.M. Stamford, Thatiana M. Stamford-Arnaud, Petrus D´Amorim Santa-Cruz, Marta C. Freitas da Silva, Galba M. Campos-Takaki, and Tânia L.M. Stamford. 2011. "Physico-Chemical Characteristics and Functional Properties of Chitin and Chitosan Produced by Mucor circinelloides Using Yam Bean as Substrate" Molecules 16, no. 8: 7143-7154. https://doi.org/10.3390/molecules16087143