Allicin Reduces the Production of α-Toxin by Staphylococcus aureus

Abstract
:1. Introduction
2. Results
2.1. Influence of Magnolol on S. aureus Growth

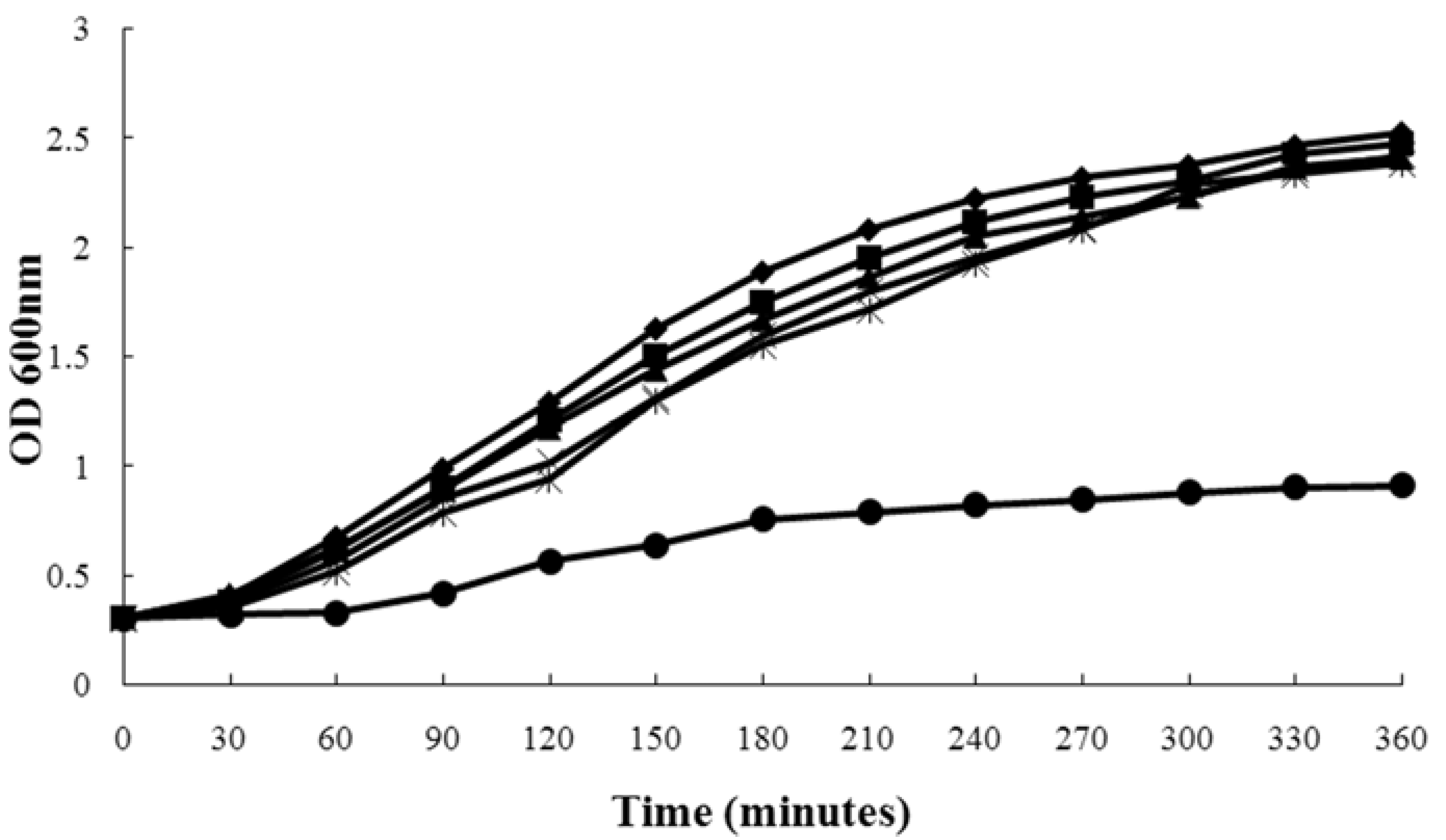{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. aureus strains | Description | Source | MIC (μg/mL) | |
|---|---|---|---|---|
| Oxacillin | Allicin | |||
| ATCC 29213 | β-Lactamase-producing oxacillin-susceptible strain, α-toxin-producing strain | ATCC | 0.25 | 64 |
| ATCC 10832 | Wood 46, a natural isolate that produces high levels of α-toxin | ATCC | 0.125 | 64 |
| BAA-1717 | USA300-HOU-MR, Isolated from adolescent patient with severe sepsis syndrome in Texas Children's Hospital, α-toxin-producing strain | ATCC | 256 | 64 |
| 8325-4 | A high-level α-toxin-producing strain derived from NCTC 8325 | Timothy J. Foster | 0.125 | 64 |
| DU 1090 | α-toxin-negative mutant of S. aureus 8325-4,prepared by insertion of a transposon in the hla gene | Timothy J. Foster | 0.125 | 32 |
| ATCC 25923 | A clinical isolate collected at Seattle in 1945 | ATCC | 0.25 | 32 |

2.2. Allicin Reduces α-Toxin Levels in S. aureus Culture Supernatants
| Haemolysis (%) of rabbit erythrocytes by culture supernatant a | |||||
|---|---|---|---|---|---|
| Strains | 0 | 2 μg/mL | 4 μg/mL | 8 μg/mL | 16 μg/mL |
| 8325-4 | 100 | 96.5 ± 3.0 | 75.1 ± 4.9 * | 64.3 ± 4.1 * | 22.1 ± 4.0 * |
| ATCC 29213 | 100 | 89.2 ± 4.5 | 62.5 ± 4.4 | 36.5 ± 5.1 * | 15.6 ± 3.5 ** |
| ATCC 10832 | 100 | 64.1 ± 4.7 | 43.2 ± 3.7 * | 30.0 ± 3.9 * | NO |
| BAA-1717 | 100 | 90.7 ± 5.2 | 58.1 ± 4.4 * | 24.3 ± 4.7 * | 6.23 ± 3.2 ** |


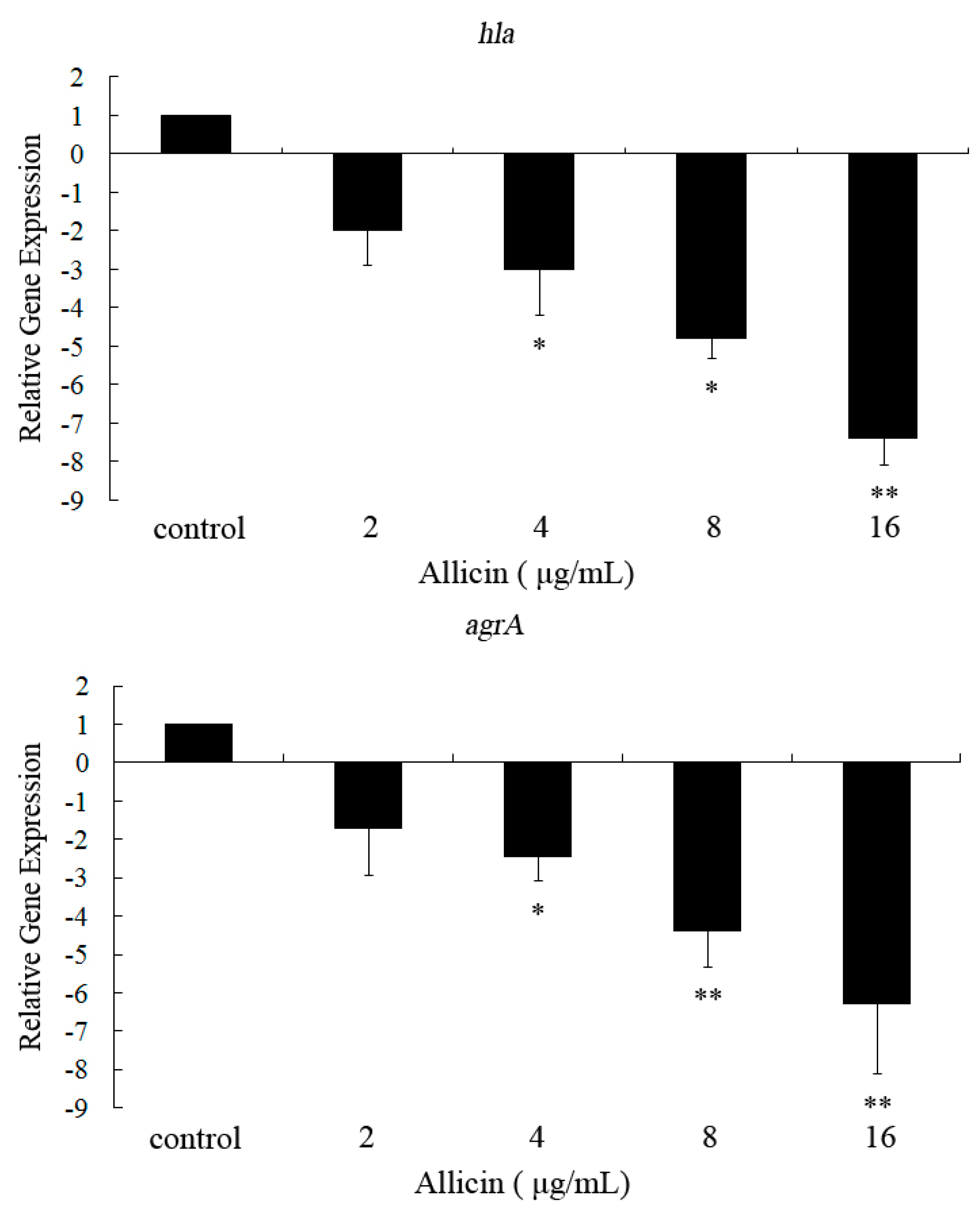
2.3. Allicin Inhibits the Transcription of hla and agrA in S. aureus

3. Discussion
4. Experimental
4.1. Bacterial Strains and Reagents
4.2. MIC Determination
4.3. Growth Curves
4.4. Haemolysis Assay
4.5. Western Blot Analysis and Proteolytic Activity Assay
4.6. RNA Isolation and Real-Time RT–PCR
| Primer | Sequence | Location within gene |
|---|---|---|
| 16S rRNA-fw | 5'-GCTGCCCTTTGTATTGTC-3' | 287–305 |
| 16S rRNA-rv | 5'-AGATGTTGGGTTAAGTCCC-3' | 446–465 |
| hla-fw | 5'-TTGGTGCAAATGTTTC-3' | 485–501 |
| hla-rv | 5'-TCACTTTCCAGCCTACT-3' | 569–586 |
| agrA-fw | 5'-TGATAATCCTTATGAGGTGCTT-3' | 111–133 |
| agrA-rv | 5'-CACTGTGACTCGTAACGAAAA-3' | 253–274 |
4.7. Statistical Analysis
5. Conclusions
Acknowledgements
- Sample Availability: Samples of the compounds are available from the authors.
References
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Wang, D.; Yu, L.; Xiang, H.; Fan, J.; He, L.; Guo, N.; Feng, H.; Deng, X. Global transcriptional profiles of Staphylococcus aureus treated with berberine chloride. FEMS Microbiol. Lett. 2008, 279, 217–225. [Google Scholar] [CrossRef]
- Athanassa, Z.; Siempos, II.; Falagas, M.E. Impact of methicillin resistance on mortality in Staphylococcus aureus VAP: A systematic review. Eur. Respir. J. 2008, 31, 625–632. [Google Scholar] [CrossRef]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar] [CrossRef]
- Kollef, M.H.; Shorr, A.; Tabak, Y.P.; Gupta, V.; Liu, L.Z.; Johannes, R.S. Epidemiology and outcomes of health-care-associated pneumonia: Results from a large US database of culture-positive pneumonia. Chest 2005, 128, 3854–3862. [Google Scholar] [CrossRef]
- Grimminger, F.; Rose, F.; Sibelius, U.; Meinhardt, M.; Potzsch, B.; Spriestersbach, R.; Bhakdi, S.; Suttorp, N.; Seeger, W. Human endothelial cell activation and mediator release in response to the bacterial exotoxins Escherichia coli hemolysin and staphylococcal alpha-toxin. J. Immunol. 1997, 159, 1909–1916. [Google Scholar]
- Jonas, D.; Walev, I.; Berger, T.; Liebetrau, M.; Palmer, M.; Bhakdi, S. Novel path to apoptosis: Small transmembrane pores created by staphylococcal alpha-toxin in T lymphocytes evoke internucleosomal DNA degradation. Infect. Immun. 1994, 62, 1304–1312. [Google Scholar]
- Bramley, A.J.; Patel, A.H.; O'Reilly, M.; Foster, R.; Foster, T.J. Roles of alpha-toxin and beta-toxin in virulence of Staphylococcus aureus for the mouse mammary gland. Infect. Immun. 1989, 57, 2489–2494. [Google Scholar]
- Bubeck Wardenburg, J.; Patel, R.J.; Schneewind, O. Surface proteins and exotoxins are required for the pathogenesis of Staphylococcus aureus pneumonia. Infect. Immun. 2007, 75, 1040–1044. [Google Scholar] [CrossRef]
- Callegan, M.C.; Engel, L.S.; Hill, J.M.; O'Callaghan, R.J. Corneal virulence of Staphylococcus aureus: Roles of alpha-toxin and protein A in pathogenesis. Infect. Immun. 1994, 62, 2478–2482. [Google Scholar]
- Patel, A.H.; Nowlan, P.; Weavers, E.D.; Foster, T. Virulence of protein A-deficient and alpha-toxin-deficient mutants of Staphylococcus aureus isolated by allele replacement. Infect. Immun. 1987, 55, 3103–3110. [Google Scholar]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Shah, S.; Stapleton, P.D.; Taylor, P.W. The polyphenol (-)-epicatechin gallate disrupts the secretion of virulence-related proteins by Staphylococcus aureus. Lett. Appl. Microbiol. 2008, 46, 181–185. [Google Scholar]
- Qiu, J.; Wang, D.; Xiang, H.; Feng, H.; Jiang, Y.; Xia, L.; Dong, J.; Lu, J.; Yu, L.; Deng, X. Subinhibitory concentrations of thymol reduce enterotoxins A and B and alpha-hemolysin production in Staphylococcus aureus isolates. PLoS one 2010, 5, e9736. [Google Scholar]
- Lang, A.; Lahav, M.; Sakhnini, E.; Barshack, I.; Fidder, H.H.; Avidan, B.; Bardan, E.; Hershkoviz, R.; Bar-Meir, S.; Chowers, Y. Allicin inhibits spontaneous and TNF-alpha induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin. Nutr. 2004, 23, 1199–1208. [Google Scholar] [CrossRef]
- Block, E. The chemistry of garlic and onions. Sci. Am. 1985, 252, 114–119. [Google Scholar] [CrossRef]
- Yamada, Y.; Azuma, K. Evaluation of the in vitro antifungal activity of allicin. Antimicrob. Agents Chemother. 1977, 11, 743–749. [Google Scholar] [CrossRef]
- Ankri, S.; Miron, T.; Rabinkov, A.; Wilchek, M.; Mirelman, D. Allicin from garlic strongly inhibits cysteine proteinases and cytopathic effects of Entamoeba histolytica. Antimicrob. Agents Chemother. 1997, 41, 2286–2288. [Google Scholar]
- Tsai, Y.; Cole, L.L.; Davis, L.E.; Lockwood, S.J.; Simmons, V.; Wild, G.C. Antiviral Properties of Garlic: In vitro Effects on Influenza B, Herpes Simplex and Coxsackie Viruses. Planta Med. 1985, 51, 460–461. [Google Scholar] [CrossRef]
- Ankri, S.; Mirelman, D. Antimicrobial properties of allicin from garlic. Microbe. Infect. 1999, 1, 125–129. [Google Scholar] [CrossRef]
- Herbert, S.; Barry, P.; Novick, R.P. Subinhibitory clindamycin differentially inhibits transcription of exoprotein genes in Staphylococcus aureus. Infect. Immun. 2001, 69, 2996–3003. [Google Scholar] [CrossRef]
- Bernardo, K.; Pakulat, N.; Fleer, S.; Schnaith, A.; Utermohlen, O.; Krut, O.; Muller, S.; Kronke, M. Subinhibitory concentrations of linezolid reduce Staphylococcus aureus virulence factor expression. Antimicrob. Agents Chemother. 2004, 48, 546–555. [Google Scholar] [CrossRef]
- Ohlsen, K.; Ziebuhr, W.; Koller, K.P.; Hell, W.; Wichelhaus, T.A.; Hacker, J. Effects of subinhibitory concentrations of antibiotics on alpha-toxin (hla) gene expression of methicillin-sensitive and methicillin-resistant Staphylococcus aureus isolates. Antimicrob. Agents Chemother. 1998, 42, 2817–2823. [Google Scholar]
- Rabinkov, A.; Miron, T.; Konstantinovski, L.; Wilchek, M.; Mirelman, D.; Weiner, L. The mode of action of allicin: trapping of radicals and interaction with thiol containing proteins. Biochim. Biophys. Acta 1998, 1379, 233–244. [Google Scholar]
- Gonzalez-Fandos, E.; Garcia-Lopez, M.L.; Sierra, M.L.; Otero, A. Staphylococcal growth and enterotoxins (A-D) and thermonuclease synthesis in the presence of dehydrated garlic. J. Appl. Bacteriol. 1994, 77, 549–552. [Google Scholar] [CrossRef]
- Arzanlou, M.; Bohlooli, S.; Jannati, E.; Mirzanejad-Asl, H. Allicin from garlic neutralizes the hemolytic activity of intra- and extra-cellular pneumolysin O in vitro. Toxicon 2011, 57, 540–545. [Google Scholar] [CrossRef]
- Arzanlou, M.; Bohlooli, S. Inhibition of streptolysin O by allicin—An active component of garlic. J. Med. Microbiol. 2010, 59, 1044–1049. [Google Scholar] [CrossRef]
- Cheung, A.; Li, D.M. Repression of hla by rot is dependent on sae in Staphylococcus aureus. Infect. Immun. 2008, 76, 1068–1075. [Google Scholar] [CrossRef]
- Novick, R.P. Autoinduction and signal transduction in the regulation of staphylococcal virulence. Mol. Microbiol. 2003, 48, 1429–1449. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute, Performance Standards for Antimicrobial Susceptibility Testing, CLSI: Wayne, PA, USA, 2005; Fifteenth informational supplement, CLSI/NCCLS document M100-S15.
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Deng, X.; Qiu, J.; Jiang, Y.; Xia, L.; Xiang, H.; Feng, H.; Pu, S.; Huang, N.; Yu, L. Subinhibitory concentrations of licochalcone A decrease alpha-toxin production in both methicillin-sensitive and methicillin-resistant Staphylococcus aureus isolates. Lett. Appl. Microbiol. 2010, 50, 223–229. [Google Scholar] [CrossRef]
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Leng, B.-F.; Qiu, J.-Z.; Dai, X.-H.; Dong, J.; Wang, J.-F.; Luo, M.-J.; Li, H.-E.; Niu, X.-D.; Zhang, Y.; Ai, Y.-X.; et al. Allicin Reduces the Production of α-Toxin by Staphylococcus aureus. Molecules 2011, 16, 7958-7968. https://doi.org/10.3390/molecules16097958
Leng B-F, Qiu J-Z, Dai X-H, Dong J, Wang J-F, Luo M-J, Li H-E, Niu X-D, Zhang Y, Ai Y-X, et al. Allicin Reduces the Production of α-Toxin by Staphylococcus aureus. Molecules. 2011; 16(9):7958-7968. https://doi.org/10.3390/molecules16097958
Chicago/Turabian StyleLeng, Bing-Feng, Jia-Zhang Qiu, Xiao-Han Dai, Jing Dong, Jian-Feng Wang, Ming-Jing Luo, Hong-En Li, Xiao-Di Niu, Yu Zhang, Yong-Xing Ai, and et al. 2011. "Allicin Reduces the Production of α-Toxin by Staphylococcus aureus" Molecules 16, no. 9: 7958-7968. https://doi.org/10.3390/molecules16097958