Carotenoids of Lettuce (Lactuca sativa L.) Grown on Soil Enriched with Spent Coffee Grounds




Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of SCG on Carotenoid and Chlorophyll Content in Lettuce Leaves

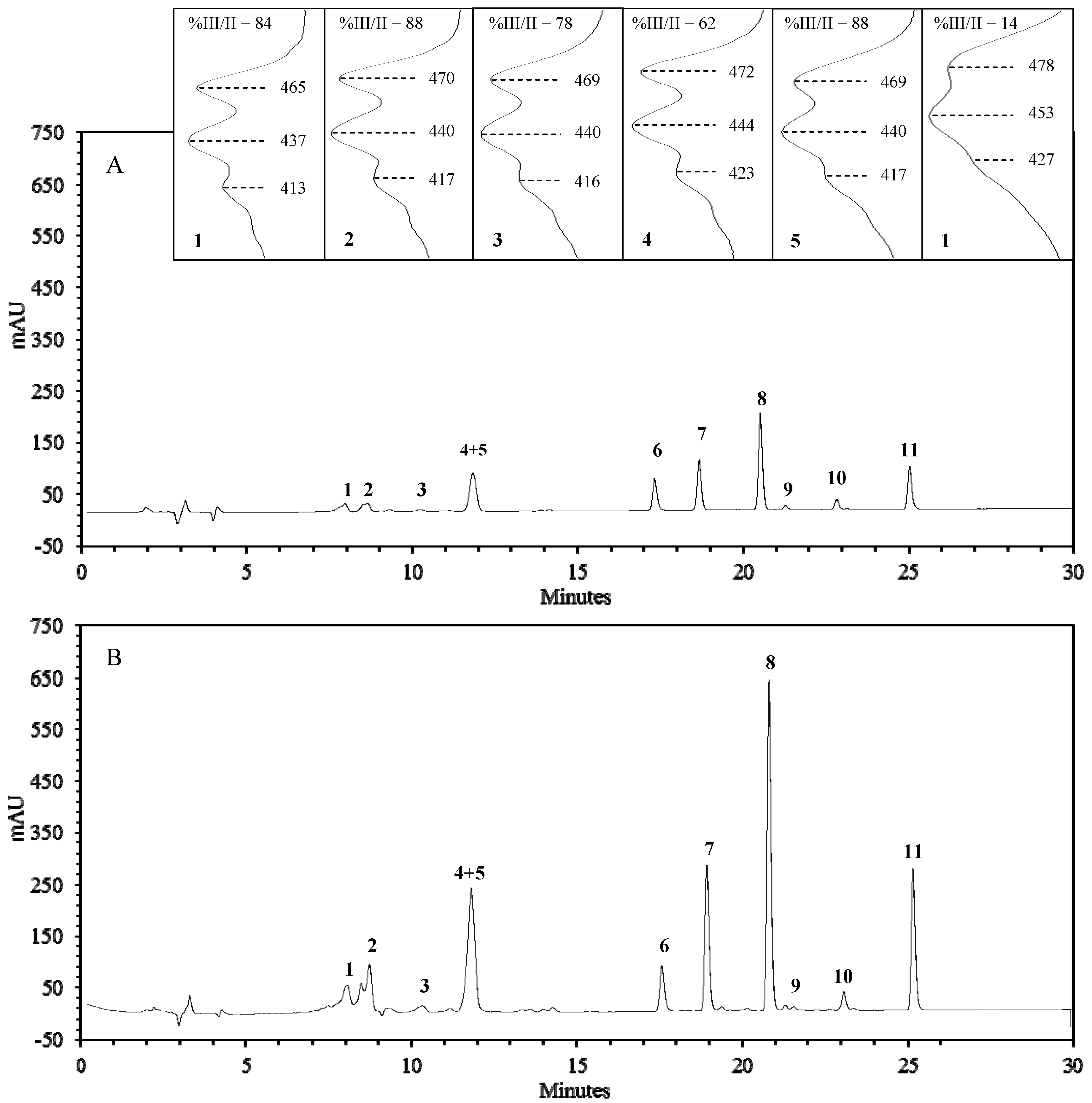{kind=link}
| % SCG | Group | Lutein | β-Carotene | Chlorophyll a | Chlorophyll b |
|---|---|---|---|---|---|
| mg/100 g | mg/100 g | mg/100 g | mg/100 g | ||
| 0 | 1 | 3.77 ± 0.03 | 3.51 ± 0.33 | 12.70 ± 0.06 | 3.44 ± 0.85 |
| 2 | 3.06 ± 0.10 | 2.27 ± 0.07 | 8.04 ± 0.10 | 3.42 ± 1.10 | |
| 3 | 4.21 ± 0.70 | 3.52 ± 0.57 | 13.00 ± 1.74 | 4.98 ± 0.55 | |
| 4 | 3.87 ± 0.55 | 3.31 ± 0.44 | 12.61 ± 1.80 | 4.11 ± 0.46 | |
| 5 | 4.49 ± 0.18 | 3.84 ± 0.30 | 11.62 ± 1.03 | 4.03 ± 0.10 | |
| n = 15 | 3.92 ± 0.65 a | 3.33 ± 0.66 a | 11.94 ± 2.06 a | 4.00 ± 0.79 a | |
| 2.5 | 1 | 4.85 ± 0.90 | 4.24 ± 0.30 | 13.04 ± 0.06 | 5.06 ± 0.62 |
| 2 | 4.01 ± 0.03 | 4.84 ± 0.96 | 12.79 ± 0.03 | 5.68 ± 2.19 | |
| 3 | 4.16 ± 0.56 | 3.55 ± 0.39 | 13.40 ± 1.10 | 4.85 ± 0.92 | |
| 4 | 6.92 ± 0.00 | 4.85 ± 0.18 | 11.02 ± 0.09 | 4.10 ± 0.28 | |
| 5 | 4.48 ± 0.86 | 3.57 ± 0.37 | 13.71 ± 0.06 | 4.21 ± 1.46 | |
| n = 15 | 4.55 ± 0.92 a | 4.21 ± 0.79 a | 12.96 ± 1.11 a,b | 4.78 ± 1.13 a,b | |
| 5 | 1 | 6.72 ± 0.42 | 4.55 ± 0.27 | 15.61 ± 1.15 | 6.11 ± 0.48 |
| 2 | 7.54 ± 0.00 | 4.82 ± 1.13 | 13.80 ± 2.99 | 5.65 ± 1.09 | |
| 3 | 6.39 ± 0.61 | 4.30 ± 0.32 | 14.71 ± 1.34 | 4.68 ± 1.13 | |
| 4 | 5.74 ± 0.26 | 4.03 ± 0.26 | 13.95 ± 0.73 | 4.75 ± 0.66 | |
| n = 12 | 6.41 ± 0.69 b | 4.42 ± 0.64 b | 14.58 ± 1.64 b,c | 5.41 ± 0.95 a,b | |
| 10 | 1 | 7.38 ± 0.90 | 4.97 ± 0.53 | 16.96 ± 1.93 | 6.96 ± 0.41 |
| 2 | 8.43 ± 0.45 | 5.54 ± 0.22 | 18.80 ± 2.09 | 5.94 ± 1.83 | |
| 3 | 7.13 ± 0.04 | 4.38 ± 0.96 | 14.82 ± 3.64 | 5.38 ± 0.30 | |
| 4 | 5.20 ± 0.30 | 3.57 ± 0.45 | 12.48 ± 0.91 | 5.00 ± 0.10 | |
| 5 | 5.51 ± 0.00 | 4.57 ± 0.51 | 15.50 ± 2.12 | 4.94 ± 1.18 | |
| n= 15 | 6.83 ± 1.34 b,c | 4.55 ± 0.84 b | 15.84 ± 3.05 c,d | 5.64 ± 1.11 a,b | |
| 15 | 1 | 8.58 ± 0.23 | 6.83 ± 1.14 | 17.01 ± 3.83 | 10.51 ± 2.43 |
| 2 | 7.21 ± 0.77 | 6.23 ± 1.36 | 17.68 ± 1.19 | 7.93 ± 1.88 | |
| 3 | 7.34 ± 0.36 | 4.96 ± 0.94 | 18.79 ± 0.01 | 7.22 ± 1.38 | |
| 4 | 7.02 ± 0.04 | 5.31 ± 0.06 | 17.79 ± 0.03 | 6.63 ± 0.21 | |
| 5 | 7.42 ± 0.64 | 4.74 ± 0.37 | 15.24 ± 1.56 | 6.26 ± 1.33 | |
| n = 15 | 7.44 ± 0.67 c | 5.70 ± 1.23 c | 17.20 ± 1.87 d | 7.96 ± 2.19 c | |
| 20 | 1 | 6.49 ± 0.02 | 5.05 ± 0.01 | 18.96 ± 0.03 | 5.23 ± 1.82 |
| 2 | 4.64 ± 0.01 | 3.80 ± 0.25 | 11.32 ± 0.00 | 6.05 ± 1.25 | |
| 3 | 6.66 ± 1.27 | 4.96 ± 1.35 | 17.57 ± 3.62 | 6.44 ± 1.46 | |
| 4 | 5.96 ± 0.54 | 4.66 ± 0.74 | 14.75 ± 0.04 | 6.27 ± 2.19 | |
| 5 | 8.18 ± 0.01 | 5.17 ± 0.41 | 19.67 ± 0.05 | 5.17 ± 0.31 | |
| 6 | 8.08 ± 1.29 | 6.07 ± 0.59 | 19.88 ± 0.00 | 5.55 ± 2.81 | |
| n =18 | 6.61 ± 1.35 b,c | 4.95 ± 0.92 b,c | 17.33 ± 3.14 d | 5.79 ± 1.44 b |

2.2. Correlation with Other Parameters Evaluated
| % SCG | Leaves biomass | Organic nitrogen | Total carotenoids | Total chlorophylls |
|---|---|---|---|---|
| (g) | (g/100g DW) | (mg/100g, FW) | (mg/100g, FW) | |
| 0 | 22.13 ± 7.11 a,b | 3.84 ± 0.25 a | 9.74 ± 1.15 a | 15.59 ± 2.47 a |
| 2.5 | 21.71 ± 4.06 a,b | 3.60 ± 0.02 a,b,c | 12.51 ± 1.44 a | 17.57 ± 1.38 a,b |
| 5 | 27.56 ± 6.40 a | 3.34 ± 0.11 b | 17.01 ± 1.42 b | 19.82 ± 1.31 a,b,c |
| 10 | 17.28 ± 6.36 b | 3.67 ± 0.21 c | 17.86 ± 3.05 b | 21.36 ± 2.97 b,c |
| 15 | 9.65 ± 4.64 c | 2.87 ± 0.08 d | 19.53 ± 2.28 b | 25.01 ± 2.26 c |
| 20 | 5.16 ± 2.91 c | 2.87 ± 0.29 d | 17.29 ± 2.97 b | 22.81 ± 3.07 c |
| Parameters | Soil | SCG |
|---|---|---|
| Moisture (%) | 55 | 63 |
| Apparent density (g.L−1) | 311 | 600 |
| pH (1:5 H2O) | 6.0 | 5.6 |
| EC (mS.cm−1) | 1.2 | 1.5 |
| Total N (%) | 0.3 * | n.d. |
| Organic N (%) | n.d. | 1.2 |
| P (%) | 0.02 | 0.02 |
| K (%) | 0.31 | 0.35 |
| Mg (%) | 0.13 | 0.10 |
| Caffeine (%) | n.d. | 0.18 |
3. Experimental
3.1. Spent Coffee Grounds
3.2. Plant Material and Growth Conditions
3.3. Carotenoid and Chlorophyll Extraction
3.4. HPLC Separation and Quantification
| Lutein (440 nm) | β-Carotene (440 nm) | Chlorophyll a (412 nm) | |
|---|---|---|---|
| Working range (μg/mL) | 1.4–21.2 | 1.8–27.8 | 1.9–58.0 |
| R2 | 0.9995 | 0.9860 | 0.9965 |
| Retention time (min) | 11.96 ± 0.10 | 24.54 ± 0.07 | 22.27 ± 0.18 |
| Precision (CV%) | 3% | 2% | 4% |
3.4. Other Chemical Analyses
3.5. Statistical Analyses
4. Conclusions
Acknowledgements
- Sample Availability: Not available.
References and Notes
- Mussatto, S.I.; Machado, E.M.S.; Martins, S.; Teixeira, J.A. Production, composition, and application of coffee and its industrial residues. Food Bioprocess Technol. 2011, 4, 661–672. [Google Scholar] [CrossRef] [Green Version]
- Franca, A.S.; Oliveira, L.S. Coffee processing solid wastes. In Agricultural Wastes; Ashworth, G.S., Azevedo, P., Eds.; Nova Science Publisher, Inc.: Hauppauge, NY, USA, 2009. [Google Scholar]
- Kasongo, R.K.; Verdoodt, A.; Kanyankagote, P.; Baert, G.; Van Ranst, E. Coffee waste as an alternative fertilizer with soil improving properties for sandy soils in humid tropical environments. Soil Use Manag. 2011, 27, 94–102. [Google Scholar] [CrossRef]
- Fan, L.; Soccol, A.T.; Pandey, A.; Vandenberghe, L.P.S.; Soccol, C.R. Effect of caffeine and tannins on cultivation and fructification of Pleurotuson coffee husks. Braz. J. Microbiol. 2006, 37, 420–424. [Google Scholar] [CrossRef]
- Kimura, M.; Rodriguez-Amaya, D.B. Carotenoid composition of hydroponic leafy vegetables. J. Agric. Food Chem. 2003, 51, 2603–2607. [Google Scholar] [CrossRef]
- Oh, M.; Trick, H.N.; Rajashekar, C.B. Secondary metabolism and antioxidants are involved in environmental adaptation and stress tolerance in lettuce. J. Plant Physiol. 2009, 166, 180–191. [Google Scholar] [CrossRef]
- Polvka, T.; Frank, H.A. Molecular factors controlling photosynthetic high harvesting by carotenoids. Acc. Chem. Res. 2010, 43, 1125–1134. [Google Scholar] [CrossRef]
- Bode, S.; Quentmeier, C.C.; Liao, P.-N.; Hafi, N.; Barros, T.; Wilk, L.; Bittner, F.; Walla, P.J. On the regulation of photosynthesis by excitonic interactions between carotenoids and chlorophylls. Proc. Natl. Acad. Sci. USA 2009, 106, 12311–12316. [Google Scholar]
- Khoo, H.; Prasad, K.N.; Kong, K.; Jiang, Y.; Ismail, A. Carotenoids and their isomers: Color pigments in fruits and vegetables. Molecules 2011, 16, 1710–1738. [Google Scholar]
- Rodriguez-Amaya, D.B. Quantitative analysis, in vitro assessment of bioavailability and antioxidant activity of food carotenoids-A review. J. Food Compos. Anal. 2010, 23, 726–740. [Google Scholar] [CrossRef]
- Maiani, G.; Periago Castón, M.J.; Catasta, G.; Toti, E.; Cambrodón, I.G.; Bysted, A.; Granado-Lorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53, S194–S218. [Google Scholar] [CrossRef]
- Ferruzzi, M.G.; Blakeslee, J. Digestion, absorption, and cancer preventative activity of dietary chlorophyll derivatives. Nutr. Res. 2007, 27, 1–12. [Google Scholar] [CrossRef]
- Britton, G. UV/Visible spectroscopy. In Carotenoids; Britton, G., Liasen-Jensen, S., Pfander, H., Eds.; Birkhäuser Verlag: Basel, Switzerland, 1995; Volume 1B, pp. 13–62, Chapter 2. [Google Scholar]
- Cardoso, P.C.; Lucia, C.M.; Stringheta, P.C.; Chaves, J.B.P.; Pinheiro-Sant’Ana, H.M. Carotene and provitamin A content of vegetables sold in Viçosa, MG, Brazil, during spring and winter. Braz. J. Pharm. Sci. 2009, 45, 527–537. [Google Scholar] [CrossRef]
- Kim, H.; Fonseca, J.M.; Choi, J.; Kubota, C.; Kwon, D.Y. Salt in irrigation water affects the nutritional and visual properties of Romaine Lettuce (Lactuca sativa L.). J. Agric. Food Chem. 2008, 56, 3772–3776. [Google Scholar] [CrossRef]
- Mou, B. Genetic variation of beta-carotene and lutein contents inlettuce. J. Am. Soc. Hortic. Sci. 2005, 130, 870–876. [Google Scholar]
- Agüero, M.V.; Barg, M.V.; Yommi, A.; Camelo, A.; Roura, S.I. Postharvest changes in water status and chlorophyll content of lettuce (Lactuca Sativa L.) and their relationship with overall visual quality. J. Food Sci. 2008, 71, S47–S55. [Google Scholar]
- Bumgarner, N.R.; Scheerens, J.C.; Mullen, R.W.; Bennett, M.A.; Ling, P.P.; Kleinhenz, M.D. Root-zone temperature and nitrogen affect the yield and secondary metabolite concentration of fall- and spring-grown, high-density leaf lettuce. J. Sci. Food Agric. 2011, 92, 116–124. [Google Scholar]
- Lynch, D.H.; Voroney, R.P.; Warman, P.R. Nitrogen availability from composts for humid region perennial grass and legume-grass forage production. J. Environ. Qual. 2004, 33, 1509–1520. [Google Scholar] [CrossRef]
- Hochmuth, G.; Hanlon, E.; Nagata, R.; Snyder, G.; Schueneman, T. Fertilization recommendations for crisphead lettuce grown on organic soils in Florida. University of Florida IFAS Extension 2009, SP153, 1–10. [Google Scholar]
- Casano, L.M.; Martin, M.; Sabate, B. Sensitivity of superoxide dismutase transcript levels and activities to oxidative stress is lower in mature-senescent than in young barley leaves. Plant Physiol. 1994, 106, 1033–1039. [Google Scholar]
- Strzalka, K.; Kostecka-Gugala, A.; Latowski, D. Carotenoids and environmental stress in plants: Significance of carotenoid-mediated modulation of membrane physical properties. Russ. J. Plant Physiol. 2003, 50, 168–172. [Google Scholar] [CrossRef]
- Mohanpuria, P.; Yadav, S.K. Retardation in seedling growth and induction of early senescence in plant upon caffeine exposure is related to its negative effect on Rubisco. Photosynthetica 2009, 47, 293–297. [Google Scholar] [CrossRef]
- Caldwell, C.R.; Britz, S.J. Effect of supplemental ultraviolet radiation on the carotenoid and chlorophyll composition of greenhouse-grown leaf lettuce (Lactuca sativa L.) cultivars. J. Food Compos. Anal. 2006, 19, 637–644. [Google Scholar] [CrossRef]
- Nicolle, C.; Carnat, A.; Fraisse, D.; Lamaison, J.L.; Rock, E.; Michel, H.; Amouroux, P.; Remesy, C. Characterization and variation of antioxidant micronutrients in lettuce (Lactuca sativa folium). J. Sci. Food Agric. 2004, 84, 2061–2069. [Google Scholar] [CrossRef]
- Zhou, C.; Zhao, D.; Sheng, Y.; Tao, J.; Yang, Y. Carotenoids in fruits of different Persimmon cultivars. Molecules 2011, 16, 624–636. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cruz, R.; Baptista, P.; Cunha, S.; Pereira, J.A.; Casal, S. Carotenoids of Lettuce (Lactuca sativa L.) Grown on Soil Enriched with Spent Coffee Grounds. Molecules 2012, 17, 1535-1547. https://doi.org/10.3390/molecules17021535
Cruz R, Baptista P, Cunha S, Pereira JA, Casal S. Carotenoids of Lettuce (Lactuca sativa L.) Grown on Soil Enriched with Spent Coffee Grounds. Molecules. 2012; 17(2):1535-1547. https://doi.org/10.3390/molecules17021535
Chicago/Turabian StyleCruz, Rebeca, Paula Baptista, Sara Cunha, José Alberto Pereira, and Susana Casal. 2012. "Carotenoids of Lettuce (Lactuca sativa L.) Grown on Soil Enriched with Spent Coffee Grounds" Molecules 17, no. 2: 1535-1547. https://doi.org/10.3390/molecules17021535