Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review


College of Agricultural Resource and Environment, Heilongjiang University, Harbin 150080, China
*
Author to whom correspondence should be addressed.
Molecules 2012, 17(3), 2367-2377; https://doi.org/10.3390/molecules17032367
Submission received: 21 January 2012
/
Revised: 15 February 2012
/
Accepted: 17 February 2012
/
Published: 24 February 2012
(This article belongs to the Section Natural Products Chemistry)
Abstract
:Beauvericin is a cyclic hexadepsipeptide mycotoxin, which has insecticidal, antimicrobial, antiviral and cytotoxic activities. It is a potential agent for pesticides and medicines. This paper reviews the bioactivity, fermentation and biosynthesis of the fungal product beauvericin.
1. Introduction
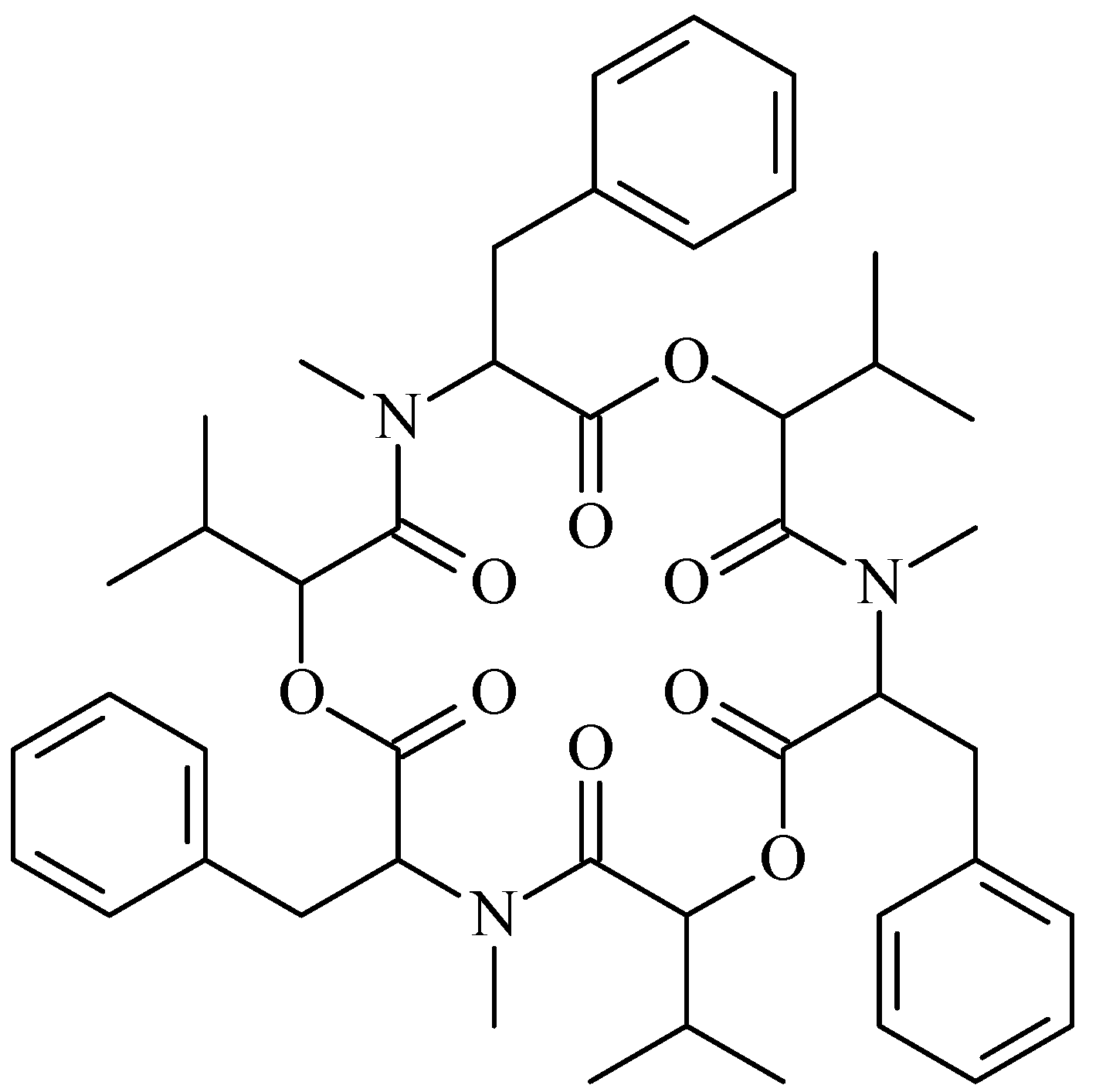
Beauvericin is a famous mycotoxin produced by many fungi, such as Beaveria bassiana and Fusarium spp. [1,2]. Beauvericin is a cyclic hexadepsipeptide (Figure 1) that belongs to the enniatin antibiotic family. It contains three D-hydroxyisovaleryl and three N-methylphenylalanyl residues in an alternating sequence [1,3]. It is structurally similar to the enniatins, which are also produced by a number of Fusarium species, but beauvericin differs in the nature of the N-methylamino acid. Owing to this difference between beauvericin and the enniatins, their bioactivities are obviously different [4]. Beauvericin was first isolated from Beaveria bassiana, which is a common and commercial entomopathogenic mycoinsecticide [1]. Beauvericin was one of the active constituents of B. bassiana [1,5,6,7] and was confirmed to have antimicrobial and anti-tumor activities. Because beauvericin is a mycotoxin, the toxicity to normal human cells has been investigated, and the detection methods for food safety have been developed; these studies are reviewed elsewhere [8,9,10,11,12,13,14]. However, there is no review about the biological activity and biosynthesis of beauvericin; therefore, this review is the first to focus on the bioactivity, fermentation and biosynthesis of beauvericin.
Figure 1.
Beauvericin structure.

2. Bioactivity of Beauvericin
2.1. Insecticidal Activity
The insecticidal activity of beauvericin was first discovered by Hamill et al. [1]. Beauvericin was confirmed as the active compound from B. bassiana against Artimia salina, which was considered a model organism to study insecticidal activity. Subsequently, the insecticidal effect of beauvericin on a microgram level was investigated on Calliphora erythrocephala, Aedes aegypti, Lygus spp., Spodoptera frugiperda and Schizaphis graminum [3,15,16,17]. Although beauvericin has a strong insecticidal activity against a broad spectrum of insect pests, it has been applied as a commercial insecticidal agent for two main reasons: first, because of the movement of insects, using an entomopathogenic fungus produced beauvericin as an insecticidal agent has more advantages than the direct use of the compound. The entomopathogenic fungus could propagate in insect bodies and be spread widely by insect movement. The entomopathogenic fungus would give rise to a good control efficiency of insects even if a small amount of the spores of the entomopathogenic fungus were used. Second, the careful evaluation of beauvericin production should ensure that beauverin production does not increase above threshold limits set by the EPA [17]. Although beauvericin is not applied directly as a commercial insecticidal agent, the insecticidal mechanism of beauvericin is still worth investigating. There are few reports about the insecticidal mechanism of beauvericin. Despite similarities between the chemical structures of beauvericin and other cyclic hexadepsipeptide mycotoxins, beauvericin is more effective against Aedes aegypti [3] and may have a unique mechanism of action. The discovery of the active mechanism of beauvericin against insects will be helpful to find new commercial insecticidal agents, reduce the threat of insecticidal agents to human cells and reveal the active mechanism of other mycotoxins.
2.2. Antitumor Activity
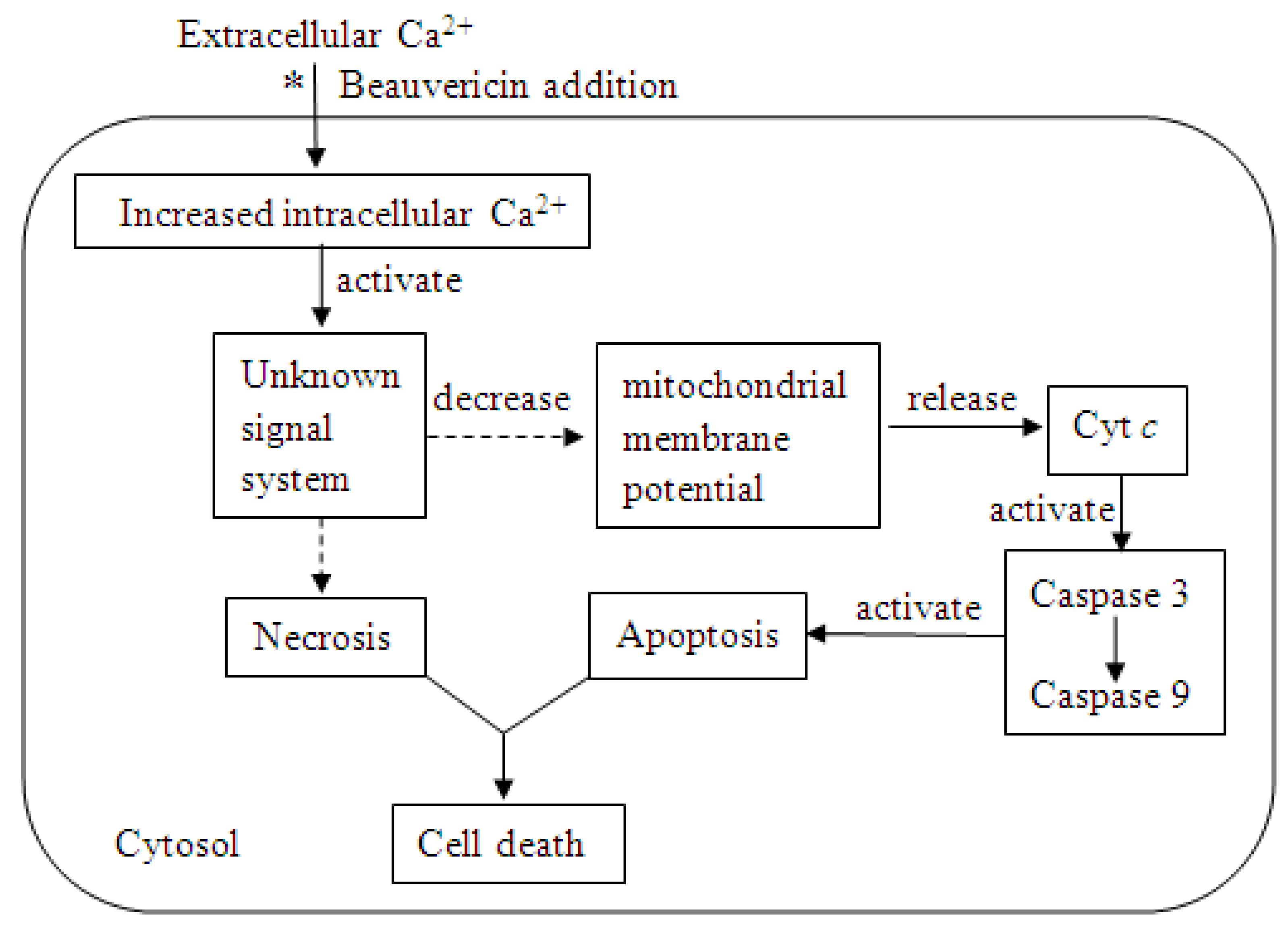
Recently, more attention has been paid to the antitumor activity of beauvericin (Table 1) [18,19,20,21,22,23,24,25]. The cytotoxicity of beauvericin to human leukemia cells has been frequently reported. We summarize the partial mechanism in Figure 2 [20,21,22,23].

{kind=link}
{kind=link}
{kind=link}
| Cell line | IC50 (μg/mL) | Reference |
|---|---|---|
| African green monkey kidney fibroblast Vero | 10 | [19] |
| Human monocytic lymphoma cells U-937 | 24 | [21] |
| Human breast cancer BC-1 | 15 | [19] |
| Human breast cancer MCF-7 | 1.4 | [24] |
| Human CNS cancer (glioma) SF-268 | 1.8 | [24] |
| Human epidermoid carcinoma KB | >20 | [19] |
| Human leukemia cell CCRF-CEM | 1–2 | [20] |
| Human non-small cell lung cancer (NSCLC) A549 | 2.4–7.8 | [22] |
| Human non-small cell lung cancer NCI-H460 | 1.1 | [24] |
| Human pancreatic carcinoma MIA Pa Ca-2 | 1.3 | [24] |
| Human promyelocytic leukemia HL-60 | 12 | [21] |
| Human retinoblastoma Y79 | 0.4–4 | [25] |
Figure 2.
Mechanism of beauvericin cytotoxicity to a human leukemia cell (*beauvericin induced extracellular Ca2+ movement into the cell resulting in an increase in the level of intracellular Ca2+; dashed arrows indicate the specific mechanisms that are unclear).
Figure 2.
Mechanism of beauvericin cytotoxicity to a human leukemia cell (*beauvericin induced extracellular Ca2+ movement into the cell resulting in an increase in the level of intracellular Ca2+; dashed arrows indicate the specific mechanisms that are unclear).

First, beauvericin induces movement of extracellular Ca2+ into the cytosol, which leads to an increased intracellular Ca2+ level. The “unknown signal system” is activated by a high level of Ca2+ and results in the release of Cyt c from the mitochondria. Finally, the caspase that is activated by Cyt c triggers apoptosis. To date, it is unclear how an increased intracellular Ca2+ concentration regulates the signal system resulting in cell death. Reports on the cytotoxicity of beauvericin to other cell lines also found that its mechanism is related to Ca2+ flux [18,26,27,28]. Thus, the unknown part shown in Figure 2 needs further investigation. The cytotoxic mechanism of beauvericin for leukemia cells could be used to determine other mechanisms of beauvericin cytotoxicities, including its role as an insecticide and an antifungal compound.
2.3. Antimicrobial Activity
2.3.1. Antibacterial Activity
Beauvericin has a strong antibacterial activity against human, animal and plant pathogenic bacteria (Table 2) [29,30,31,32], with no selectively between Gram-positive and Gram-negative bacteria. Unlike other antibiotics (such as penicillin) that block the peptidoglycan biosynthesis of Gram-positive bacteria, the bacterial cell wall is not the antibacterial mode of action of beauvericin, although these antibiotics and beauvericin are both from amino acids that are produced by fungi. It is possible that other cell organelles or enzyme systems are the targets of beauvericin. Despite its broad-spectrum antibacterial activity, the antifungal activity of beauvericin as a single agent is rarely reported. Therefore, the target of beauvericin is different between bacteria and fungi and could include targets such as the ribosome or the cell nucleus. The activity of beauvericin should be investigated against drug resistant bacteria. Based on the antibacterial activity against plant pathogens [32], beauvericin could be utilized in the control of non-food crop diseases. As a potential antibacterial agent, beauvericin could be used to solve the problems of drug resistance, deadly bacterial infections [30,31] and non-food crop disease.
| Gram-positive bacteria | Reference | Gram-negative bacteria | Reference |
|---|---|---|---|
| Bacillus spp. | [29,32] | Agrobacterium tumefaciens | [32] |
| Bifidobacterium adolescentis | [29] | Escherichia coli CECT 4782 | [31] |
| Clostridium perfringens | [29,31] | Escherichia coli | [32] |
| Enterococcus faecium | [31] | Pseudomonas aeruginosa | [31] |
| Eubaterium biforme | [29] | Pseudomonas lachrymans | [32] |
| Listeria monocytogenes | [31] | Salmonella enterica | [31] |
| Mycobacterium tuberculosis | [30] | Shigella dysenteriae | [31] |
| Peptostreptococcus spp. | [29] | Xanthomonas vesicatoria | [32] |
| Staphylococcu haemolyticus | [32] | Yersinia enterocolitica | [31] |
| Paenibacillus spp. | [29] |
2.3.2. Antifungal Activity
The lack of antifungal activity of beauvericin as a single agent could be because it is a fungal product. However, Zhang et al. [33] and Fukuda et al. [34] report the antifungal activity of beauvericin in combination with ketoconazole or miconazole. Beauvericin (0.5 mg/kg) combined with ketoconazole (0.5 mg/kg) had remarkable antifungal activity against Candida parapsilosis, which can quickly cause high mortality rates, particularly in neonates. Both beauvericin and ketoconazole alone have little to no effect on C. parapsilosis. If the antifungal mechanism of beauvericin is similar to the cytotoxic mechanism in leukemia cells, it would indicate that the fungus itself can inhibit the “unknown signal system” (Figure 2) until another compound, such as ketoconazole, is added to unlock the signaling system. The method of combining beauvericin with another compound offers a new way to develop and utilize the biological activity of beauvericin.
3. Biosynthesis of Beauvericin
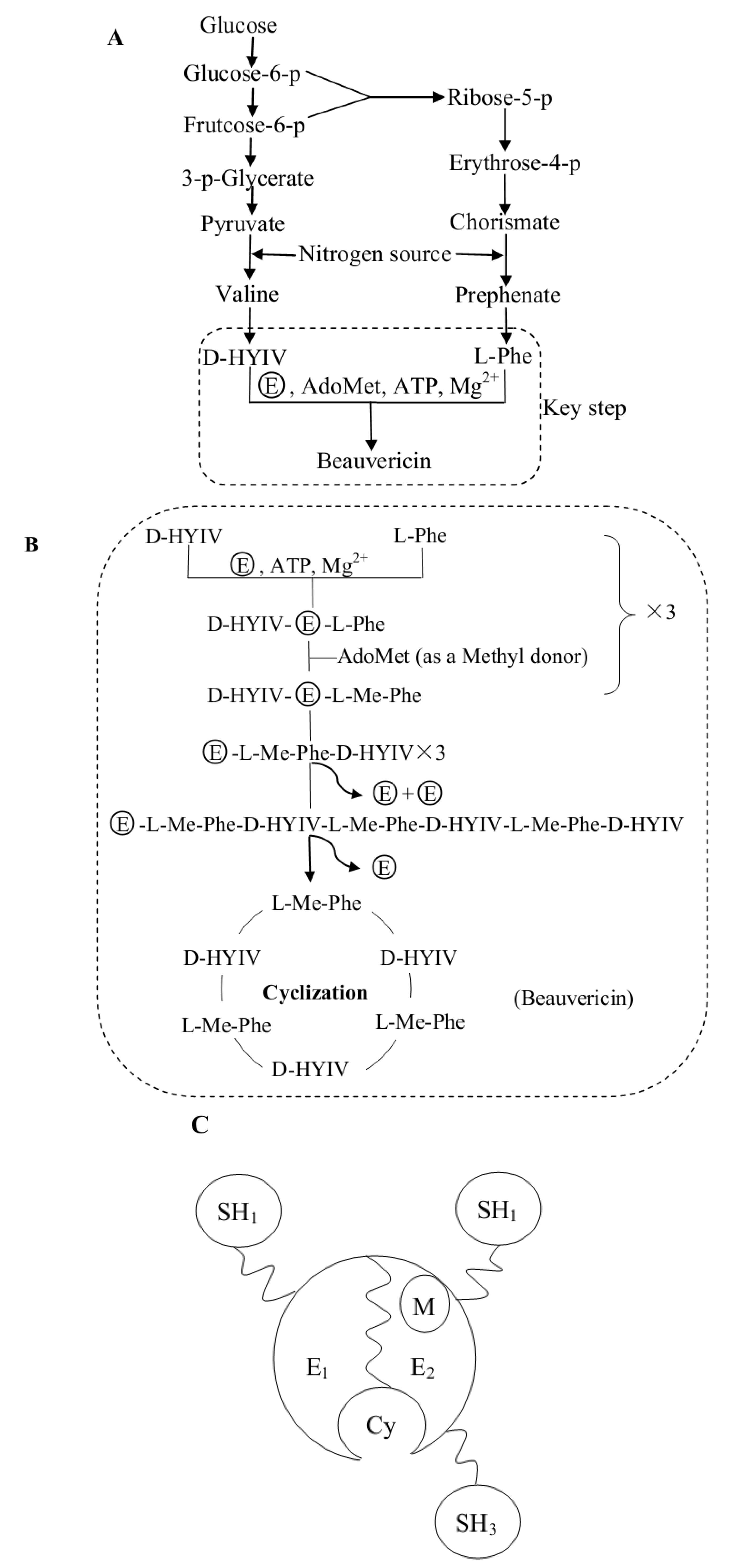
Based on the biosynthetic studies of beauvericin, in addition to a comparison to cyclic depsipeptide biosynthesis, we built a beauvericin biosynthetic pathway as shown in Figure 3A, B, C [35,36,37,38,39,40]. In Figure 3A, the “nitrogen source” could be any amino acid that offers nitrogen to L-phenylalanine or valine by transamination. Also, a hexose or a pentose could be the carbon source instead of glucose. Of seven potential carbon sources, glucose was the most effective for beauvericin biosynthesis, as reported by Xu et al. [41]. The last step in the pathway is key (Figure 3A) and is strictly dependent on the presence of the amino acid L-phenylalanine (L-Phe), the hydroxy acid D-hydroxyisovaleric acid (D-HYIV), ATP/Mg2+, S-adenosyl-methionine (AdoMet), and the beauvericin synthetase. AdoMet is the source of the methyl group for the L-phenylalanyl residues. The beauvericin synthetase (shown in Figure 3C) is a multifunctional enzyme that catalyzes depsipetide formation and is a single polypeptide chain with a molecular mass of approximately 250 kDa. Beauvericin biosynthesis is catalyzed by the beauvericin synthetase via a nonribosomal, thiol-templated mechanism. As shown in Figure 3B, we summarize the five intermediates in the key step of beauvericin biosynthesis. First, two beauvericin synthetase modules (Figure 3C) are activated by the corresponding L-Phe and D-HYIV, which are covalently attached to the enzyme-bound 4'-phosphopantetheinyl arm as thioesters. Next, the L-phenylalanyl residues are N-methylated by AdoMet. Third, a peptide bond is formed. Fourth, a linear hexadepsipeptide intermediate is synthesized. In the final step, the linear hexadepsipeptide is cyclized to make beauvericin. The optimum pH for beauvericin formation was pH 7.2, and the optimum temperature was 25~27 °C; the beauvericin synthetase was inactive if the temperature was more than 30 °C [35,37]. To date, the amino acid sequence and the gene encoding the beauvericin synthetase are unclear. However, the specific recognition sites of the beauvericin synthetase vary considerably and may contribute to the biosynthesis of beauvericin analogs. For example, the L-Phe could be replaced by a different aromatic or aliphatic amino acid [37,38]. The analogs could give more support to apply beauvericin as a lead for potential medicines and pesticides.
Figure 3.
A, Beauvericin biosynthesis pathway; B, The specific substeps of the key step ( ![Molecules 17 02367 i001]() represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
 represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
Figure 3.
A, Beauvericin biosynthesis pathway; B, The specific substeps of the key step ( ![Molecules 17 02367 i001]() represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
represents the beauvericin synthetase; the dashed frame indicates the key step of beauvericin synthesis); C, The possible structure of the beauvericin synthetase (E1 is the D-HYIV module; E2 is the L-Phe module; SH1,2,3 are the 4'-phosphopantetheine residues corresponding to D-HYIV, L-Phe, and the linear hexadepsipeptide acceptor; M is the N-methyltransferase domain; Cy is the cyclization cavity).
4. Production of Beauvericin by Fungal Fermentation
At present, many compounds that were isolated from fungi are used as medicines and pesticides. Fermentation conditions for beauvericin production have been studied recently as it is a potential commercial fungal product. The conclusions from carbon and nitrogen source screening for beauvericin production indicated that the optimal carbon source was glucose, the optimal organic nitrogen source was peptone and the optimal inorganic nitrogen source was NaNO3 [40,41]. Using valine and phenylalanine as nitrogen sources did not increase beauvericin production, which indicated that the fungus offered enough precursors for beauvericin production by itself when an optimal medium was used. Substrate (glucose) inhibition was identified in the fermentation of beauvericin using F. redolens Dzf2. The fed-batch (glucose was fed to the culture) was successfully used in beauvericin fermentation, which provided ample substrates for beauvericin production and reduced the substrate inhibition [42]. Beauvericin is an intracellular product; only a small amount of beauvericin is exported to the medium. Interestingly, macroporous polystyrene can efficiently extract beauvericin from mycelium cells. An integrated fermentation, with an in situ product recovery process using macroporous polystyrene, was used to effectively enhance beauvericin recovery in the mycelial liquid culture [43]. It facilitated the extraction and separation of beauvericin from the fungal mycelia. In addition, a modified Monod model of the fed-batch method for efficient beauvericin production was developed. This model with substrate (glucose) inhibition, together with the stoichiometric relationships for biomass, substrate and product, can be applied to predict the optimization scheme for fed-batch fermentation. Overall, mycelial fermentation of fungi, such as Fusarium spp., is a feasible and promising process for the production of beauvericin. Although beauvericin production was increased to 400 mg/L in the mycelial liquid culture by optimization of the fermentation process [43], the quantities of beauvericin produced were lower than those of other industrial and commercial antibiotics. The study of the fermentation process and the biosynthesis should be continued to improve beauvericin production. For example, two-phase fermentation could be investigated to increase beauvericin production. It is possible that intracellular beauvericin could be extracted by using an organic and non-polar solution in the fermentation process to enhance production.
5. Perspectives
The detection of beauvericin has been adequately studied (picogram level) [8,9,10,11,12,13,14], but it is our contention that the bioactive development of beauvericin is relatively ignored. Studies on medicines and pesticides indicate that it will be impossible to find a “perfect compound” without any side effects in a long time. Therefore, we believe that development of existing compounds (such as beauvericin) could be the best way to discover new medicines and pesticides. It has been confirmed that beauvericin is a fungal product with not only various kinds of bioactivities, but also with unique uncharacterized active mechanisms. From now on, we should put our focus to unravel each active mechanism of beauvericin rather than only testing more microorganisms or cell lines. We should pay more attention to the control or reduction of the beauvericin risk to humans rather than setting up more detection methods for beauvericin with all kinds of instruments. For example, it is critical to investigate the reason why fungi cannot be inhibited by beauvericin alone. If this mechanism is revelaed, it will be expected that we can better manipulate the bioactivity of beauvericin, to some extent, similar to a “light switch” that we can control. It will be a great discovery in pharmacology and toxicology. Beauvericin is a bioactive compound with the potential for use as a medicine or a pesticide, especially because of its potential use in deadly diseases such as cancer or viral and bacterial infections. We believe beauvericin will become a star compound. In addition, beauvericin can only be biosynthesized by several specific fungal genera: Beauveria, Paecilomyces, Polyporus, Isaria and Fusarium [2,44,45]. It has been suggested that the content of beauvericin can serve as a chemotaxonomic marker of fungi. The reason why beauvericin is only biosynthesized by those specific fungal species should be investigated. According to the biosynthetic mechanism, fermentation process for beauvericin production will increase its commercial interest. Because of its broad and significant bioactivities, beauvericin could become a commercial product from fungi in the future.
Acknowledgements
Jianyong Wu (Hong Kong Polytechnic University) and Ligang Zhou (China Agricultural University) recommended us to write a review on beauvericin. We appreciate their support for this review. This review is funded by the National Natural Science Foundation of China (30970487 and 31100036).
- Sample Availability: Beauvericin is available from the authors.
References and Notes
- Hamill, R.L.; Higgens, G.E.; Boaz, H.E.; Gorman, M. The structure of beauvericin, a new desipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 49, 4255–4258. [Google Scholar]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin Production by Fusarium Species. Appl. Environ. Microb. 1998, 64, 3084–3088. [Google Scholar]
- Grove, J.F.; Pople, M. The insecticidal activity of beauvericin and the enniatin complex. Mycopathologia 1980, 70, 103–105. [Google Scholar] [CrossRef]
- Shin, C.G.; An, D.G.; Song, H.H.; Lee, C. Beauvericin and enniatins H, I and MK1688 are new potent inhibitors of human immunodeficiency virus type-1 integrase. J. Antibiot. 2009, 62, 687–690. [Google Scholar] [CrossRef]
- Champlin, F.R.; Grula, E.A. Noninvolvement of beauvericin in the entomopathogenicity of Beauveria bassiana. Appl. Environ. Microb. 1979, 37, 1122–1126. [Google Scholar]
- Steinkraus, D.C.; Tugwell, N.P. Beauveria bassiana (Deuteromycotina: Moniliales) effects on Lygus lineolaris (Hemiptera: Miridae). J. Entomol. Sci. 1997, 31, 79–90. [Google Scholar]
- Wagner, B.L.; Lewis, L.C. Colonization of corn, Zea mays, by the entomopathogenic fungus Beauveria bassiana. Appl. Environ. Microb. 2000, 66, 68–73. [Google Scholar]
- Plattner, R.D.; Nelson, P.E. Production of beauvericin by a strain of Fusarium proliferatum isolated from corn fodder for swine. Appl. Environ. Microb. 1994, 60, 3894–3896. [Google Scholar]
- Krska, R.; Schuhmacher, R.; Grasserbauer, M.; Scott, P.M. Determination of the Fusarium mycotoxin beauvericin at μg/kg levels in corn by high-performance liquid chromatography with diode-array detection. J. Chromatogr. A 1996, 746, 233–238. [Google Scholar] [CrossRef]
- Logrieco, A.; Rizzo, A.; Ferrancane, R.; Ritieni, A. Occurrence of beauvericin and enniatins in wheat affected by Fusarium avenaceum Head Blight. Appl. Environ. Microb. 2002, 68, 82–85. [Google Scholar] [CrossRef]
- Ambrosino, P.; Galvano, F.; Fogliano, V.; Logrieco, A.; Fresa, R.; Ritieni, A. Supercritical fluid extraction of beauvericin from maize. Talanta 2004, 62, 523–530. [Google Scholar] [CrossRef]
- Sørensen, J.L.; Nielsen, K.F.; Rasmussen, P.H.; Thrane, U. Development of a LC-MS/MS method for the analysis of enniatins and beauvericin in whole fresh and ensiled maize. J. Agric. Food Chem. 2008, 56, 10439–10443. [Google Scholar]
- Logrieco, A.; Moretti, A.; Ritieni, A.; Caiaffa, M.F.; Macchia, L. Beauvericin: chemistry, biology and significance. In Advances in Microbial Toxin Research and Its Biotechnological Exploitation; Upadhyay, R., Ed.; Kluwer Academic: New York, NY, USA, 2002; pp. 23–30. [Google Scholar]
- Ganassi, S.; Moretti, A.; Pagliai, A.M.B.; Logrieco, A.; Sabatini, M.A. Effects of beauvericin on Schizaphis graminum (Aphididae). J. Invertebr. Pathol. 2002, 80, 90–96. [Google Scholar] [CrossRef]
- Jestoi, M. Emerging Fusarium -Mycotoxins Fusaproliferin, Beauvericin, Enniatins, And Moniliformin—A Review. Crit. Rev. Food Sci. 2008, 48, 21–49. [Google Scholar] [CrossRef]
- Fornelli, F.; Minervini, F.; Logrieco, A. Cytotoxicity of fungal metabolites to lepidopteran (Spodoptera frugiperda) cell line (SF-9). J. Invertebr. Pathol. 2004, 85, 74–79. [Google Scholar] [CrossRef]
- Leland, J.E.; McGuire, M.R.; Grace, J.A.; Jaronski, S.T.; Ulloa, M.; Park, Y.; Plattner, R.D. Strain selection of a fungal entomopathogen, Beauveria bassiana, for control of plant bugs (Lygus spp.)(Heteroptera: Miridae). Biol. Control 2005, 35, 104–114. [Google Scholar]
- Que, F.G.; Gores, G.I.; LaRusso, N.F. Development and initial application of an in vitro model of apoptosis in rodent cholangiocytes. Am. J. Phys. 1997, 272, 106–115. [Google Scholar]
- Nilanonta, C; Isaka, M.; Kittakoop, P.; Trakulnaleamasai, S.; Tanticharoen, M.; Thebtaranonth, Y. Precursor-directed biosynthesis of beauvericin analogs by the insect pathogenic fungus Paecilomyces tenuipes BCC 1614. Tetrahedron 2002, 58, 3355–3360. [Google Scholar]
- Jow, G.; Chou, C.; Chen, B.; Tsai, J. Beauvericin induces cytotoxic effects in human acute lymphoblastic leukemia cells through cytochrome c release, caspase 3 activation: The causative role of calcium. Cancer Lett. 2004, 216, 165–173. [Google Scholar] [CrossRef]
- Calo, L.; Fornelli, F.; Ramires, R.; Nenna, S.; Tursi, A.; Caiaffa, M.F.; Macchia, L. Cytotoxic effects of the mycotoxin beauvericin to human cell lines of myeloid origin. Pharmacol. Res. 2004, 49, 73–77. [Google Scholar] [CrossRef]
- Lin, H.; Lee, Y.; Chen, B.; Tsai, M.; Lu, J.; Chou, C.; Jow, G. Involvement of Bc1-2 family, cytochrome c and caspase 3 in induction of apotosis by beauvericin in human non-small cell lung cancer cells. Cancer Lett. 2004, 230, 248–259. [Google Scholar]
- Chen, B.; Tsai, M.; Jow, G. Induction of calcium influx from extracellular fluid by beauvericin in human leukemia cells. Biochem. Biophs. Res. Commun. 2006, 340, 134–139. [Google Scholar] [CrossRef]
- Zhan, J.; Burns, A.M.; Liu, M.X.; Faeth, S.H.; Gunatilaka, A.A.L. Search for cell motility and angiogenesis inhibitors with potential anticancer activity: Beauvericin and other constituents of two endophytic strains of Fusarium oxysporum. J. Nat. Prod. 2007, 70, 227–232. [Google Scholar] [CrossRef]
- Cheng, C.; Chang, K.; Lee, Y. Antiproliferative effect of beauvericin on retinoblastoma. Fu-Jen J. Med. 2009, 7, 167–169. [Google Scholar]
- Ojcjus, D.M.; Zychlinsky, A; Zheng, L.M.; Young, J.D. Inophore-induced apoptosis: Role of DNA fragmentation and calcium fluxes. Exp. Cell Res. 1991, 197, 43–49. [Google Scholar]
- Wu, S.N.; Chen, H.; Liu, Y.C.; Chiang, H.T. Block of L-type Ca2+ current by beauvericin, a toxic cyclopeptide, in the NG108-15 neuronal cell line. Chem. Res. Toxicol. 2002, 15, 854–860. [Google Scholar] [CrossRef]
- Kouti, K.; Lemmens, M.; Lemmens-Gruber, R. Beauvericin induced channels in ventricular myocytes and liposomes. Biochim. Biophys. Acta 2003, 1609, 203–210. [Google Scholar] [CrossRef]
- Castlebury, L.A.; Sutherland, J.B.; Tanner, L.A.; Henderson, A.L.; Cerniglia, C.E. Use of a bioassay to evaluate the toxicity of beauvericin to bacteria. World J. Microb. Biot. 1999, 15, 119–121. [Google Scholar] [CrossRef]
- Nilanonta, C.; Isaka, M.; Kittakoop, P.; Palittapongarnpim, P.; Kamchonwongpaisan, S.; Pittayakhajonwut, D.; Tanticharoen, M.; Thebtaranonth, Y. Antimycobacterial and antiplasmodial cyclodepsipeptides from the insect pathogenic fungus Paecilomyces tenuipes BCC1614. Planta Med. 2000, 66, 756–758. [Google Scholar] [CrossRef]
- Meca, G.; Sospedra, I.; Soriano, J.M.; Ritieni, A.; Moretti, A.; Manes, J. Antibacterial effect of the bioactive compound beauvericin produced by Fusarium proliferatum on solid medium of wheat. Toxicon 2010, 56, 349–354. [Google Scholar] [CrossRef]
- Xu, L.; Wang, J.; Zhao, J.; Li, P.; Shan, T.; Wang, J.; Li, X.; Zhou, L. Beauvericin from the endophytic fungus, Fusarium redolens, isolated from Dioscorea zingiberensis and its antibacterial activity. Nat. Prod. Commun. 2010a, 5, 811–814. [Google Scholar]
- Zhang, L.; Yan, K.; Zhang, Y.; Huang, R.; Bian, J.; Zheng, C.; Sun, H.; Chen, Z.; Sun, N.; An, R.; et al. High-throughput synergy screening identifies microbial metabolites as combination agents for the treatment of fungal infections. Proc. Nat. Acad. Sci. USA 2007, 104, 4606–4611. [Google Scholar]
- Fukuda, K.; Arai, M.; Yamaguchi, Y.; Masuma, R. New beauvericins, potentiators of antifungal miconazole activity produced by Beauveria sp. FKI-1366. J. Antibiot. 2004, 110–116. [Google Scholar]
- Peeters, H.; Zocher, R.; Madry, N.; Kleinkauf, H. Incorporation of radioactive precursors into beauvericin produced by Paecilomyces fumoso-roseus. Phytochemistry 1983, 22, 1719–1720. [Google Scholar]
- Peeters, H.; Zocher, R.; Madry, N. Oelrichs, P.B. Cell-free synthesis of the desipeptide beauvericin. J. Antibiot. 1983, 36, 1762–1766. [Google Scholar]
- Peeters, H.; Zocher, R.; Kleinkauf, H. Synthesis beauvericin by a multifunctional enzyme. J. Antibiot. 1988, 12, 352–359. [Google Scholar]
- Kleinkauf, H.; Dohren, H. Nonribosomal biosynthesis of peptide antibiotic. Eur. J. Biochem. 1990, 192, 1–15. [Google Scholar] [CrossRef]
- Hornbogen, T.; Glinski, M.; Zocher, R. Biosynthesis of depsipeptide mycotoxins in Fusarium. Eur. J. Plant Pathol. 2002, 108, 713–718. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Optimization of a liquid medium for beauvericin production in Fusarium redolens Dzf2 mycelial culture. Biotechnol. Bioproc. Eng. 2010, 15, 460–466. [Google Scholar] [CrossRef]
- Lee, H.S.; Song, H.H.; Ahn, J.H.; Shin, C.G.; Lee, G.P.; Lee, C. Statistical optimization of growth medium for the production of the entomopathogenic and phytotoxic cyclic depsipeptide beauvericin from Fusarium oxysporum KFCC 11363P. J. Microbiol. Biotechnol. 2008, 18, 138–144. [Google Scholar]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Modeling of Fusarium redolens Dzf2 mycelial growth kinetics and optimal fed-batch fermentation for beauvericin production. J. Ind. Microbiol. Biot. 2011, 38, 1187–192. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Zhou, L.; Wu, J. Enhanced beauvericin production with in situ adsorption in mycelial liquid culture of Fusarium redolens Dzf2. Process Biochem. 2009, 44, 1063–1067. [Google Scholar] [CrossRef]
- Luangsa-ard, J.J.; Berkaew, P.; Ridkaew, R.; Hywel-jones, N.L.; Isaka, M. Abeauvericin hot spot in the genus Isaria. Mycol. Res. 2009, 113, 1389–1395. [Google Scholar] [CrossRef]
- Moretti, A.; Mule, G.; Ritieni, A.; Logrieco, A. Further data on the production of beauvericin, enniatins and fusaproliferin and toxicity to Artemia salina by Fusarium species of Gibberella fujikuroi species complex. Int. J. Food Microbiol. 2007, 118, 158–163. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Wang, Q.; Xu, L. Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review. Molecules 2012, 17, 2367-2377. https://doi.org/10.3390/molecules17032367
AMA Style
Wang Q, Xu L. Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review. Molecules. 2012; 17(3):2367-2377. https://doi.org/10.3390/molecules17032367
Chicago/Turabian StyleWang, Qinggui, and Lijian Xu. 2012. "Beauvericin, a Bioactive Compound Produced by Fungi: A Short Review" Molecules 17, no. 3: 2367-2377. https://doi.org/10.3390/molecules17032367