α-Mangostin Enhances Betulinic Acid Cytotoxicity and Inhibits Cisplatin Cytotoxicity on HCT 116 Colorectal Carcinoma Cells

Abstract
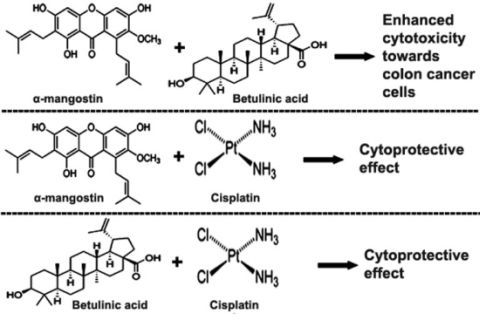
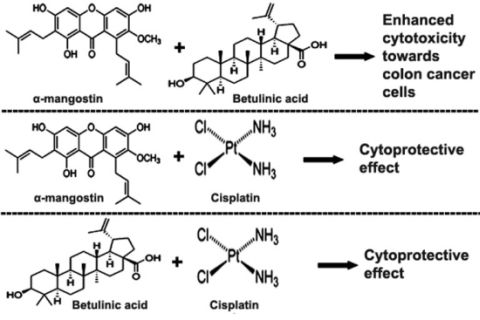
:
1. Introduction
2. Results
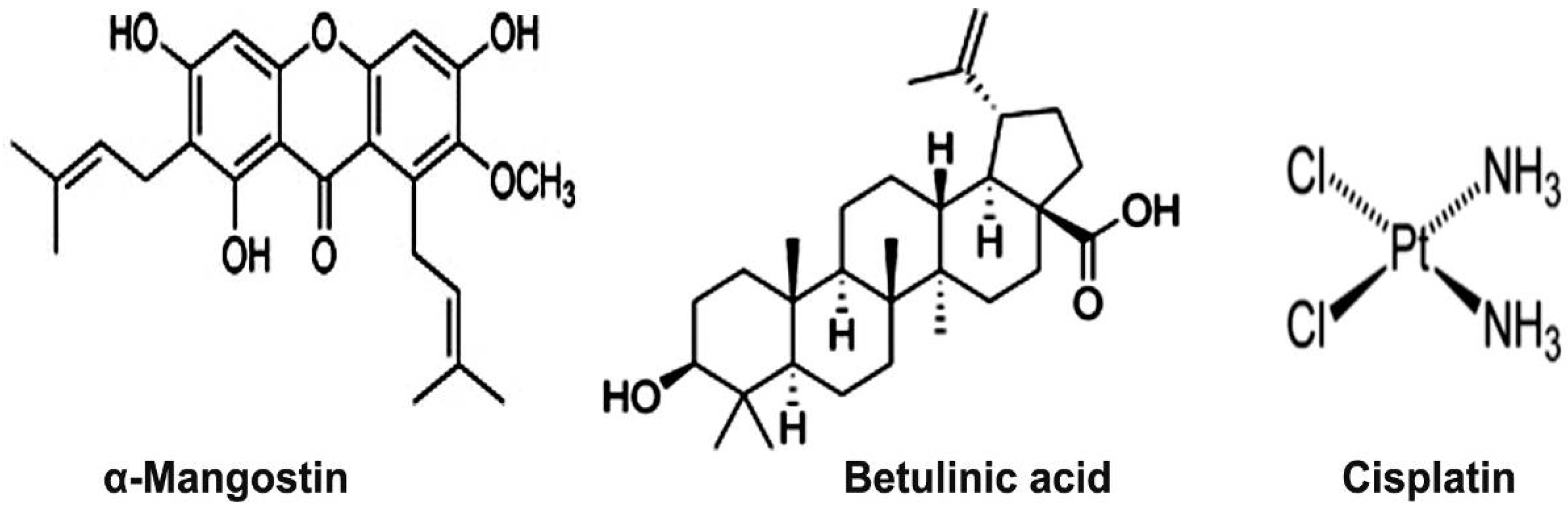
2.1. Phytochemical Analysis
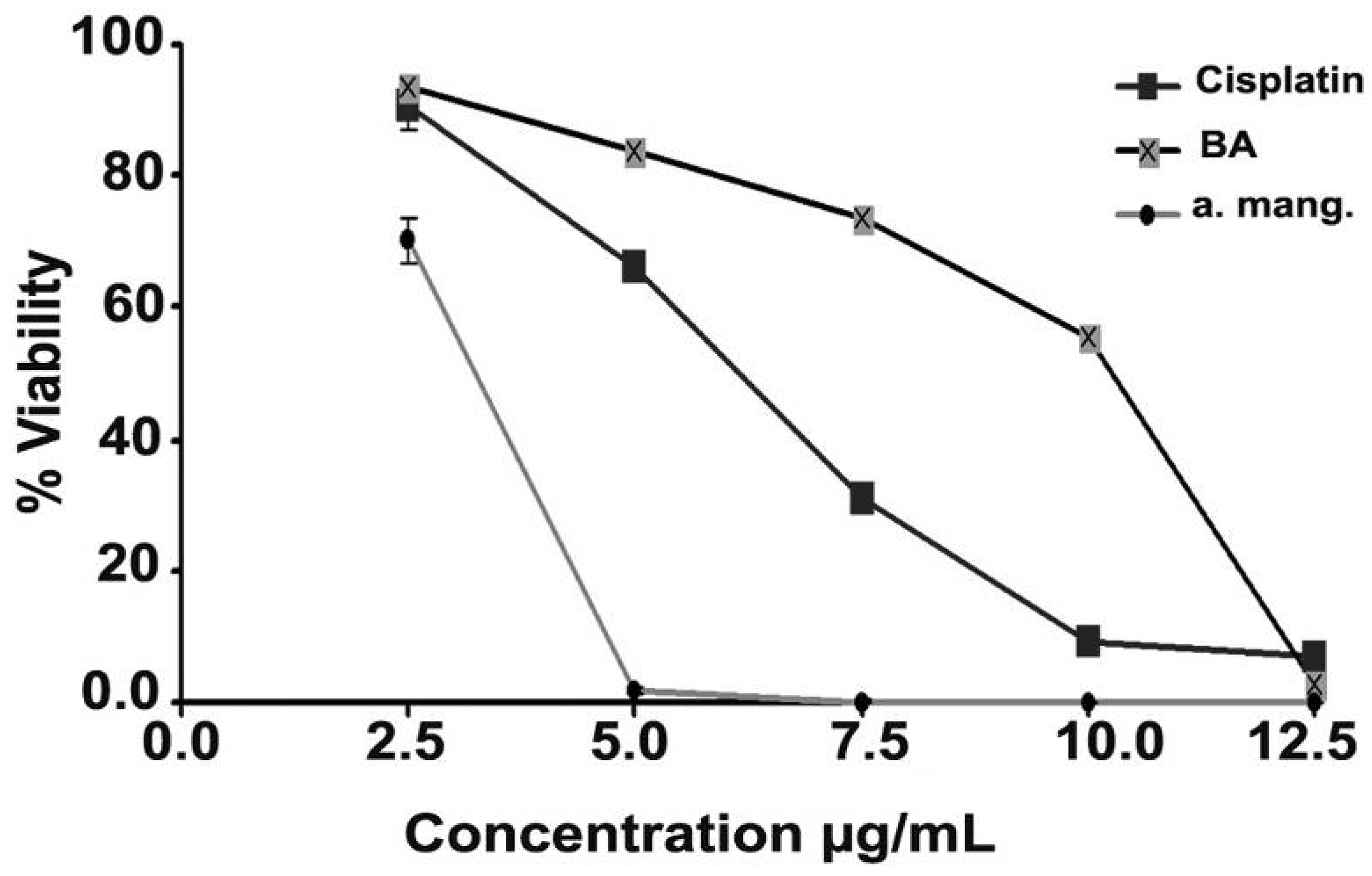
2.2. Cytotoxicity
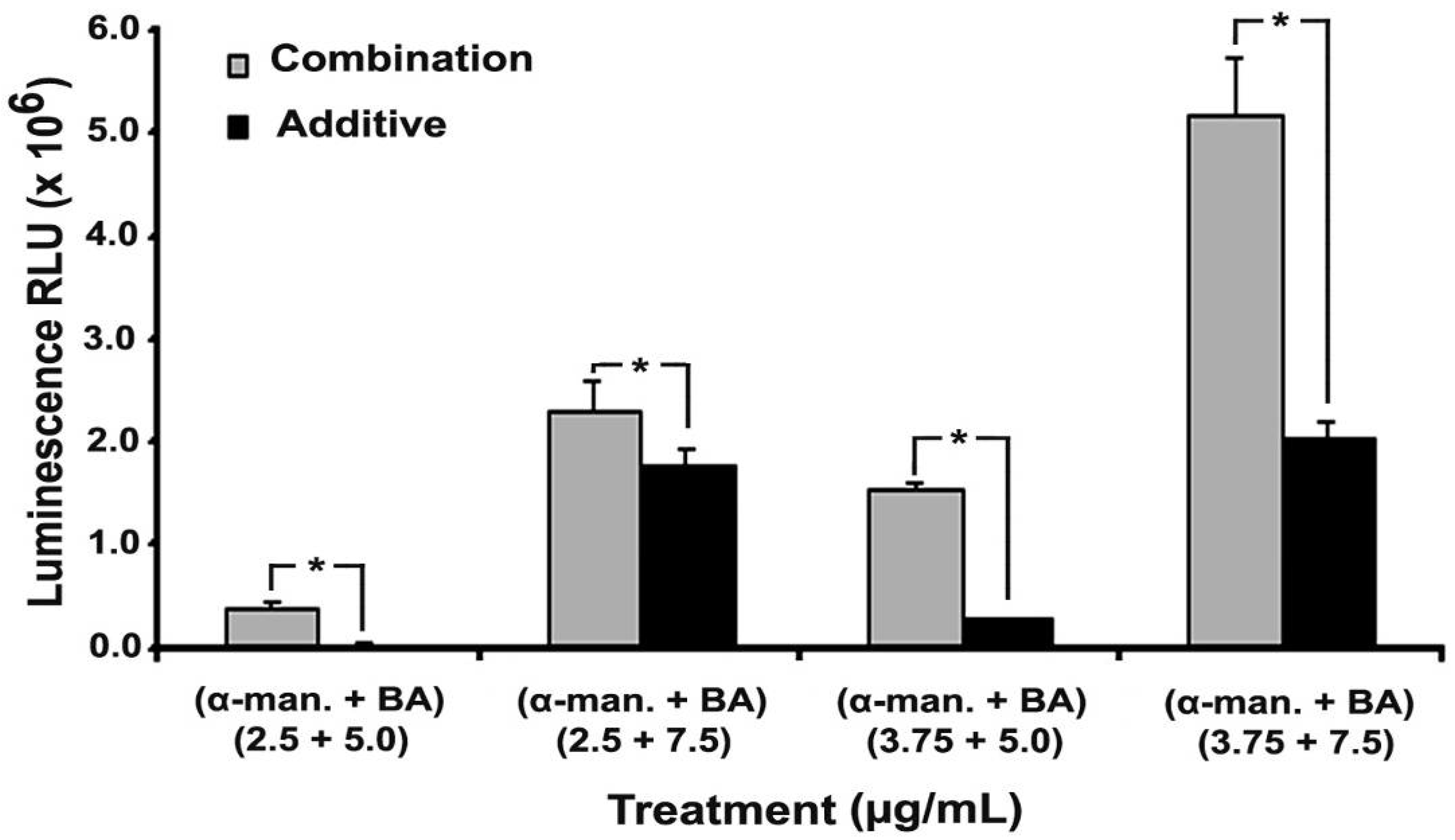
2.3. Combination of α-Mangostin, BA and Cisplatin
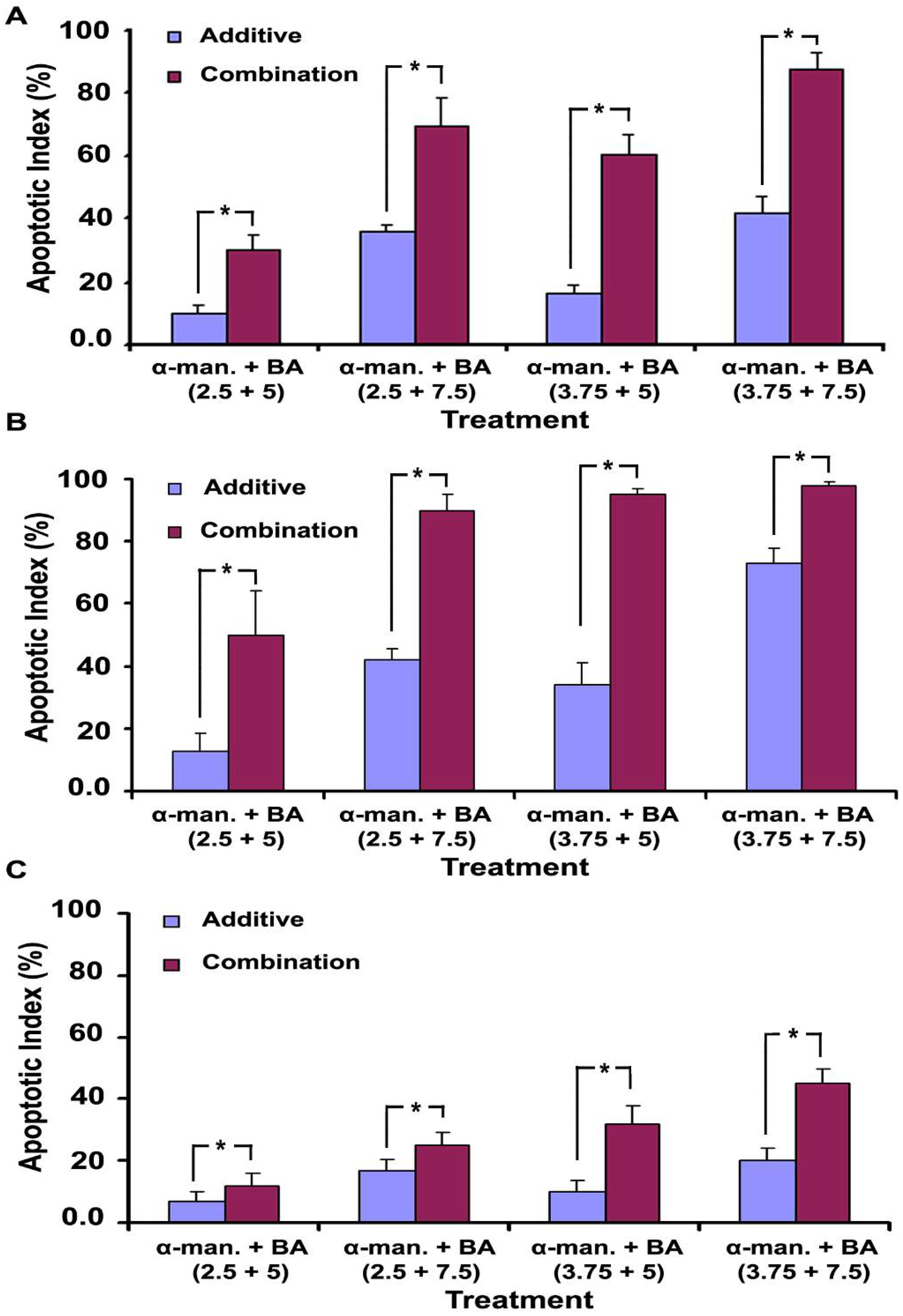
2.4. Effect on Caspases-3/7
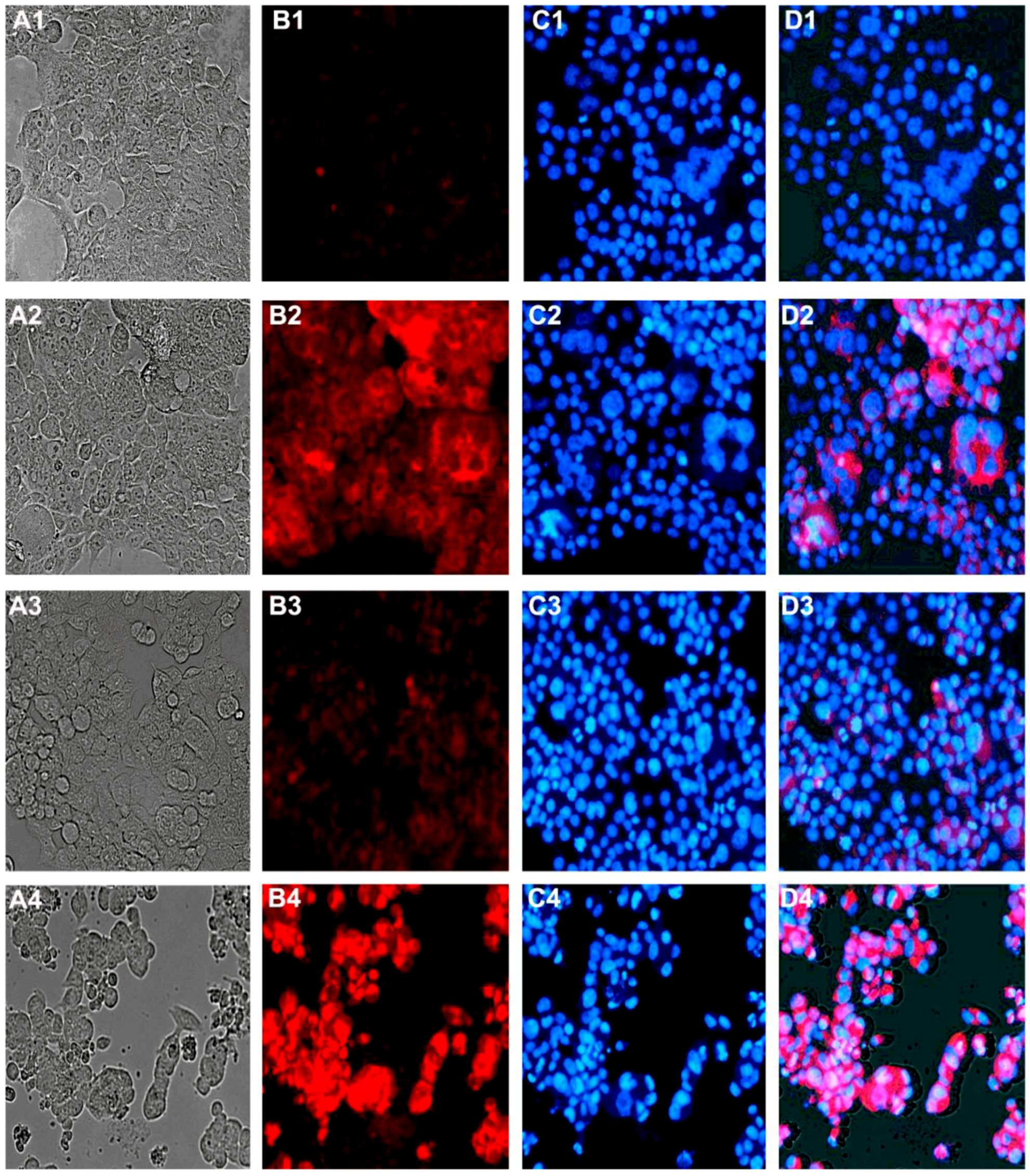
2.5. Effect on Cellular Morphology, Mitochondrial Membrane Potential and Chromatin Condensation
2.6. Effect on Cell Signaling Pathways
3. Discussion
4. Experimental
4.1. Cell Culture and Reagents
4.2. Isolation and Characterization of BA
4.3. Isolation and Characterization of α-Mangostin
4.4. Direct-Infusion Electrospray Mass Spectrometry
4.5. Cell Viability
4.6. Caspases-3/7
4.7. Mitochondrial Membrane Potential and Chromatin Condensation
4.8. Luciferase Assay
4.9. Statistical Analysis
5. Conclusions
Acknowledgements
References and Notes
- International Agency for Research on Cancer. All Cancers (excluding non-melanoma skin cancer) Incidence and Mortality Worldwide in 2008. In Globocan 2008; IARC: Lyon, France, 2010. [Google Scholar]
- Cercek, A.; Saltz, L.B. First-line treatment of patients with metastatic colorectal cancer: An overview of recent data on chemotherapy plus targeted agents. Clin. Colorectal Cancer 2008, 7 (Suppl 2), S47–S51. [Google Scholar] [CrossRef] [PubMed]
- Obolskiy, D.; Pischel, I.; Siriwatanametanon, N.; Heinrich, M. Garcinia mangostana L.: A phytochemical and pharmacological review. Phytother. Res. 2009, 23, 1047–1065. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Akao, Y.; Kobayashi, E.; Ohguchi, K.; Ito, T.; Tanaka, T.; Iinuma, M.; Nozawa, Y. Induction of apoptosis by xanthones from mangosteen in human leukemia cell lines. J. Nat. Prod. 2003, 66, 1124–1127. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Akao, Y.; Yi, H.; Ohguchi, K.; Ito, T.; Tanaka, T.; Kobayashi, E.; Iinuma, M.; Nozawa, Y. Preferential target is mitochondria in alpha-mangostin-induced apoptosis in human leukemia HL60 cells. Bioorg. Med. Chem. 2004, 12, 5799–5806. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Betulinic acid for cancer treatment and prevention. Int. J. Mol. Sci. 2008, 9, 1096–1107. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, R.; Jaggi, M.; Rajendran, P.; Siddiqui, M.J.A.; Srivastava, S.K.; Vardhan, A.; Burman, A.C. Betulinic acid and its derivatives as anti-angiogenic agents. Bioorg. Med. Chem. Lett. 2004, 14, 2181–2184. [Google Scholar] [CrossRef] [PubMed]
- Leong, H.; Mathur, P.S.; Greene, G.L. Green tea catechins inhibit angiogenesis through suppression of STAT3 activation. Breast Cancer Res. Treat. 2009, 117, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Lin, C.C.; Chou, C.C.; Hsu, C.P. The effect of Longan seed polyphenols on colorectal carcinoma cells. Eur. J. Clin. Invest. 2010, 40, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol. 1995, 49, 57–68. [Google Scholar] [CrossRef]
- Recio, M.C.; Giner, R.M.; Manez, S.; Gueho, J.; Julien, H.R.; Hostettmann, K.; Rios, J.L. Investigations on the steroidal anti-inflammatory activity of triterpenoids from Diospyros leucomelas. Planta Med. 1995, 61, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Chintharlapalli, S.; Papineni, S.; Ramaiah, S.K.; Safe, S. Betulinic acid inhibits prostate cancer growth through inhibition of specificity protein transcription factors. Cancer Res. 2007, 67, 2816–2823. [Google Scholar] [CrossRef] [PubMed]
- Chintharlapalli, S.; Papineni, S.; Lei, P.; Pathi, S.; Safe, S. Betulinic acid inhibits colon cancer cell and tumor growth and induces proteasome-dependent and -independent downregulation of specificity proteins (Sp) transcription factors. BMC Cancer 2011, 11. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.R.; Kim, K.J.; Choi, C.H.; Lee, T.B.; Han, S.I.; Han, H.K.; Lim, S.C. Effect of betulinic acid on anticancer drug-resistant colon cancer cells. Basic Clin. Pharmacol. Toxicol. 2007, 101, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Kroemer, G. Targeting mitochondrial apoptosis by betulinic acid in human cancers. Drug Discov. Today 2009, 14, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Siddik, Z.H. Cisplatin: Mode of cytotoxic action and molecular basis of resistance. Oncogene 2003, 22, 7265–7279. [Google Scholar] [CrossRef] [PubMed]
- Boulikas, T.; Vougiouka, M. Cisplatin and platinum drugs at the molecular level. Oncol. Rep. 2003, 10, 1663–1682. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, R.; Das, B.; Yeger, H.; Koren, G.; Shibuya, M.; Thorner, P.; Baruchel, S.; Malkin, D. Cisplatin treatment increases survival and expansion of a highly tumorigenic side-population fraction by upregulating VEGF/Flt1 autocrine signaling. Oncogene 2008, 27, 3923–3934. [Google Scholar] [CrossRef] [PubMed]
- Späth-Schwalbe, E.; Lange, C.; Genvresse, I.; Krüger, L.; Eucker, J.; Schweigert, M.; Sezer, O.; Budach, V.; Possinger, K. Influence of amifostine on toxicity of CHOP in elderly patients with aggressive non-Hodgkin’s lymphoma-a phase II study. Anticancer Drugs 2002, 13, 395. [Google Scholar] [CrossRef] [PubMed]
- Ozben, T. Mechanisms and strategies to overcome multiple drug resistance in cancer. FEBS Lett. 2006, 580, 2903–2909. [Google Scholar] [CrossRef] [PubMed]
- Mayer, L.D.; Shabbits, J.A. The role for liposomal drug delivery in molecular and pharmacological strategies to overcome multidrug resistance. Cancer Metastasis Rev. 2001, 20, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Jabr-Milane, L.S.; van Vlerken, L.E.; Yadav, S.; Amiji, M.M. Multi-functional nanocarriers to overcome tumor drug resistance. Cancer Treat. Rev. 2008, 34, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.M.; Denis-Gay, M.; Ratinaud, M.H. Assessment of fluorochromes for cellular structure and function studies by flow cytometry. Biol. Cell 1993, 78, 1–13. [Google Scholar] [CrossRef]
- Sensi, S.L.; Ton-That, D.; Sullivan, P.G.; Jonas, E.A.; Gee, K.R.; Kaczmarek, L.K.; Weiss, J.H. Modulation of mitochondrial function by endogenous Zn2+ pools. Proc. Natl. Acad. Sci. USA 2003, 100, 6157–6162. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, Z.; Traganos, F.; Staiano-Coico, L.; Kapuscinski, J.; Melamed, M. Interactions of rhodamine 123 with living cells studied by flow cytometry. Cancer Res. 1982, 42, 799–806. [Google Scholar] [PubMed]
- Fulda, S.; Debatin, K.M. Betulinic acid induces apoptosis through a direct effect on mitochondria in neuroectodermal tumors. Med. Pediatr. Oncol. 2000, 35, 616–618. [Google Scholar] [CrossRef]
- Rajput, A.; Dominguez San Martin, I.; Rose, R.; Beko, A.; Levea, C.; Sharratt, E.; Mazurchuk, R.; Hoffman, R.M.; Brattain, M.G.; Wang, J. Characterization of HCT116 human colon cancer cells in an orthotopic model. J. Surg. Res. 2008, 147, 276–281. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Fang, L.W.; Lu, S.P.; Chou, C.K.; Luh, T.Y.; Lai, M.Z. DNA-damaging reagents induce apoptosis through reactive oxygen species-dependent Fas aggregation. Oncogene 2003, 22, 8168–8177. [Google Scholar] [CrossRef] [PubMed]
- Pedraza-Chaverri, J.; Reyes-Fermin, L.M.; Nolasco-Amaya, E.G.; Orozco-Ibarra, M.; Medina-Campos, O.N.; Gonzalez-Cuahutencos, O.; Rivero-Cruz, I.; Mata, R. ROS scavenging capacity and neuroprotective effect of alpha-mangostin against 3-nitropropionic acid in cerebellar granule neurons. Exp. Toxicol. Pathol. 2009, 61, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Pérez, Y.; Morales-Bárcenas, R.; García-Cuellar, C.M.; López-Marure, R.; Calderon-Oliver, M.; Pedraza-Chaverri, J.; Chirino, Y.I. The [alpha]-mangostin prevention on cisplatin-induced apoptotic death in LLC-PK1 cells is associated to an inhibition of ROS production and p53 induction. Chem. Biol. Interact. 2010, 188, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.J.; Lee, Y.J.; Kim, J.S.; Kang, D.G.; Lee, H.S. Protective role of betulinic acid on TNF-[alpha]-induced cell adhesion molecules in vascular endothelial cells. Biochem. Biophys. Res. Commun. 2010, 391, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Szuster-Ciesielska, A.; Pilipów, K.; Kandefer-Szersze, M. Protective effect of betulin and betulinic acid on acetaminophen and ethanol-induced cytotoxicity and reactive oxygen species production in HepG2 cells. J. Pre-Clin. Clin. Res. 2010, 4, 96–100. [Google Scholar]
- Karna, E.; Szoka, L.; Palka, J.A. Betulinic acid inhibits the expression of hypoxia-inducible factor 1alpha and vascular endothelial growth factor in human endometrial adenocarcinoma cells. Mol. Cell. Biochem. 2010, 340, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Rzeski, W.; Stepulak, A.; Szymanski, M.; Sifringer, M.; Kaczor, J.; Wejksza, K.; Zdzisinska, B.; Kandefer-Szerszen, M. Betulinic acid decreases expression of bcl-2 and cyclin D1, inhibits proliferation, migration and induces apoptosis in cancer cells. Naunyn Schmiedebergs Arch. Pharmacol. 2006, 374, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; He, J.; Fang, J.; Hong, M. Antitumor effect of betulinic acid on human acute leukemia K562 cells in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2010, 30, 453–457. [Google Scholar] [PubMed]
- Aisha, A.F.A.; Abu-Salah, K.M.; Alrokayan, S.A.; Siddiqui, M.J.; Ismail, Z.; Abdul Majid, A.M.S. Syzygium aromaticum extracts as good source of betulinic acid and potential anti-breast cancer. Rev. Bras. Farmacogn. 2011. [Google Scholar] [CrossRef]
- Jost, L.M.; Kirkwood, J.M.; Whiteside, T.L. Improved short- and long-term XTT-based colorimetric cellular cytotoxicity assay for melanoma and other tumor cells. J. Immunol. Methods 1992, 147, 153–165. [Google Scholar] [CrossRef]
- Aisha, A.F.A.; Sahib, H.B.; Abu-Salah, K.M.; Darwis, Y.; Abdul Majid, A.M.S. Cytotoxic and anti-angiogenic properties of the stem bark extract of Sandoricum koetjape. Int. J. Cancer Res. 2009, 5, 105–114. [Google Scholar]
- Aisha, A.F.A.; Abu-Salah, K.M.; Nassar, Z.D.; Siddiqui, M.J.; Ismail, Z.; Abdul Majid, A.M.S. Antitumorigenicity of xanthones-rich extract from Garcinia mangostana fruit rinds on HCT 116 human colorectal carcinoma cells. Rev. Bras. Farmacogn. 2011, 21, 1025–1034. [Google Scholar] [CrossRef]
- Johnson, L.V.; Walsh, M.L.; Chen, L.B. Localization of mitochondria in living cells with rhodamine 123. Proc. Natl. Acad. Sci. USA 1980, 77, 990–994. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, S.S.; Yu, Y.; Nautiyal, J.; Patel, B.B.; Majumdar, A.P. The Wnt/beta-catenin pathway regulates growth and maintenance of colonospheres. Mol. Cancer 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Webb, J. Effect of More Than One Inhibitor, in Enzyme and Metabolic Inhibitors; Academic Press: New York, NY, USA, 1963; Volume 1, p. 72. [Google Scholar]
Sample Availability: Samples of the compound (α-mangostin) are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1-α-Mangostin (2.5 µg/mL) and betulinic acid (BA) | |||||
| BA (µg/mL) | BA (% inhibition) | α-Man. (% inhibition) | Combination (% inhibition) | Additive (% inhibition) | Fold change in BA cytotoxicity |
| 2.5 | 4 ± 0.2 | 2.0 ± 1.7 | 51 ± 2.5 | 6.5 ± 1.1 | 7.9 ± 0.9 |
| 5.0 | 9 ± 1.0 | 91 ± 1.6 | 11.6 ± 1.5 | 7.9 ± 0.8 | |
| 7.5 | 20 ± 1.5 | 99 ± 0.6 | 22.3 ± 1.8 | 4.5 ± 0.4 | |
| 10.0 | 50 ± 2.0 | 100 ± 0.1 | 51.4 ± 2.1 | 1.9 ± 0.1 | |
| 2-α-Mangostin (2.5 µg/mL) and cisplatin (Cis.) | |||||
| Cis. (µg/mL) | Cis. (% inhibition) | α-Man. (% inhibition) | Combination (%inhibition) | Additive (% inhibition) | Fold change in Cis. cytotoxicity |
| 2.5 | 16 ± 2.3 | 2 ± 1.7 | 18 ± 3.3 | 18.8 ± 2.2 | 1.0 ± 0.09 |
| 5.0 | 36 ± 1.5 | 12 ± 3.6 | 37.9 ±1.7 | 0.3 ± 0.08 | |
| 7.5 | 83 ± 1.0 | 9 ± 0.3 | 83.5 ±1.0 | 0.1 ± 0.01 | |
| 10.0 | 97 ± 1.0 | 19 ± 4.5 | 97.1 ±1.0 | 0.2 ± 0.04 | |
| 3-BA (2.5 µg/mL) and Cisplatin. | |||||
| Cis. (µg/mL) | Cis. (% inhibition) | BA (% inhibition) | Combination (% inhibition) | Additive (% inhibition) | Fold change in Cis. cytotoxicity |
| 2.5 | 16 ± 2.3 | 4.0 ± 0.2 | 18 ± 1.2 | 19.7 ± 1.9 | 0.9 ± 0.03 |
| 5.0 | 36 ± 1.5 | 28 ± 0.5 | 38.6 ± 1.6 | 0.7 ± 0.02 | |
| 7.5 | 83 ± 1.0 | 49 ± 1.8 | 83.7 ±1.0 | 0.6 ± 0.01 | |
| 10.0 | 97 ± 1.0 | 72 ± 1.2 | 97.1 ±1.0 | 0.7 ± 0.01 | |
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Aisha, A.F.A.; Abu-Salah, K.M.; Ismail, Z.; Majid, A.M.S.A. α-Mangostin Enhances Betulinic Acid Cytotoxicity and Inhibits Cisplatin Cytotoxicity on HCT 116 Colorectal Carcinoma Cells. Molecules 2012, 17, 2939-2954. https://doi.org/10.3390/molecules17032939
Aisha AFA, Abu-Salah KM, Ismail Z, Majid AMSA. α-Mangostin Enhances Betulinic Acid Cytotoxicity and Inhibits Cisplatin Cytotoxicity on HCT 116 Colorectal Carcinoma Cells. Molecules. 2012; 17(3):2939-2954. https://doi.org/10.3390/molecules17032939
Chicago/Turabian StyleAisha, Abdalrahim F. A., Khalid M. Abu-Salah, Zhari Ismail, and Amin Malik Shah Abdul Majid. 2012. "α-Mangostin Enhances Betulinic Acid Cytotoxicity and Inhibits Cisplatin Cytotoxicity on HCT 116 Colorectal Carcinoma Cells" Molecules 17, no. 3: 2939-2954. https://doi.org/10.3390/molecules17032939