Extracts of Lycoris aurea Induce Apoptosis in Murine Sarcoma S180 Cells

Abstract
:1. Introduction
2. Results and Discussion
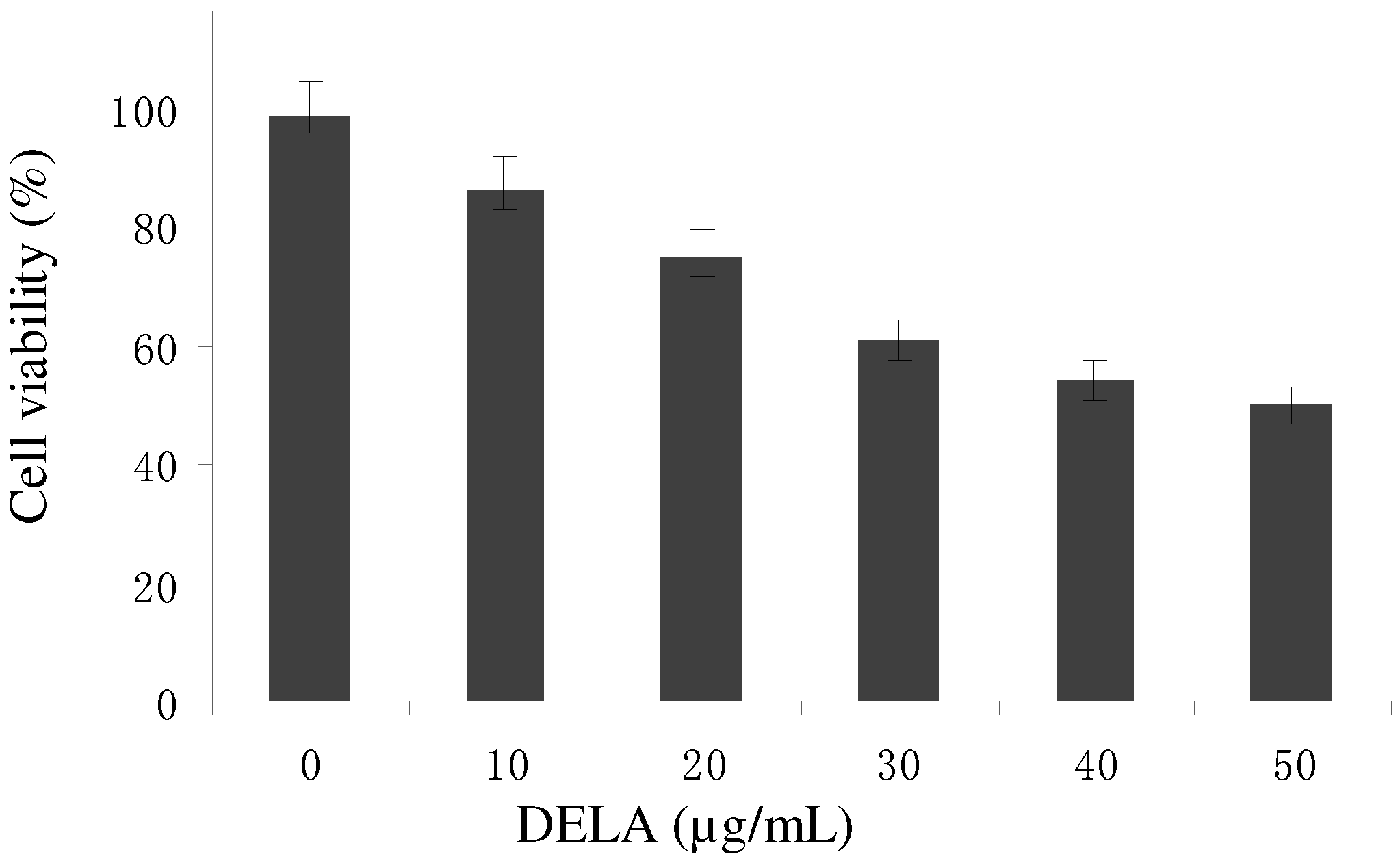
2.1. Antitumour Activity in Vitro and in Vivo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Dose (mg/kg) | Weight (g) | Tumor weight (g) | Inhibitory rate (%) | Thymus index (mg/g) | Spleen index (mg/g) | |
|---|---|---|---|---|---|---|---|
| Before treatment | After treatment | ||||||
| model control | - | 19.49 ± 1.21 | 28.36 ± 3.05 | 1.29 ± 0.38 | 2.19 ± 0.80 | 7.43 ± 1.29 | |
| CTX | 20 | 19.33 ± 1.38 | 22.57 ± 1.47 | 0.41 ± 0.12 ** | 68.22 | 1.03 ± 0.16 | 5.28 ± 1.04 |
| DELA | 120 | 19.18 ± 1.57 | 27.94 ± 2.25 | 0.60 ± 0.14 ** | 53.49 | 2.98 ± 0.61 *a | 9.95 ± 1.87 *a |
| DELA | 40 | 19.44 ± 1.82 | 28.35 ± 2.61 | 0.81 ± 0.19 | 37.21 | 2.46 ± 0.53 a | 9.22 ± 1.59 a |
| DELA | 20 | 19.61 ± 1.16 | 29.17 ± 2.79 | 0.93 ± 0.22 | 27.9 | 2.23 ± 0.59 a | 8.98 ± 1.40 a |
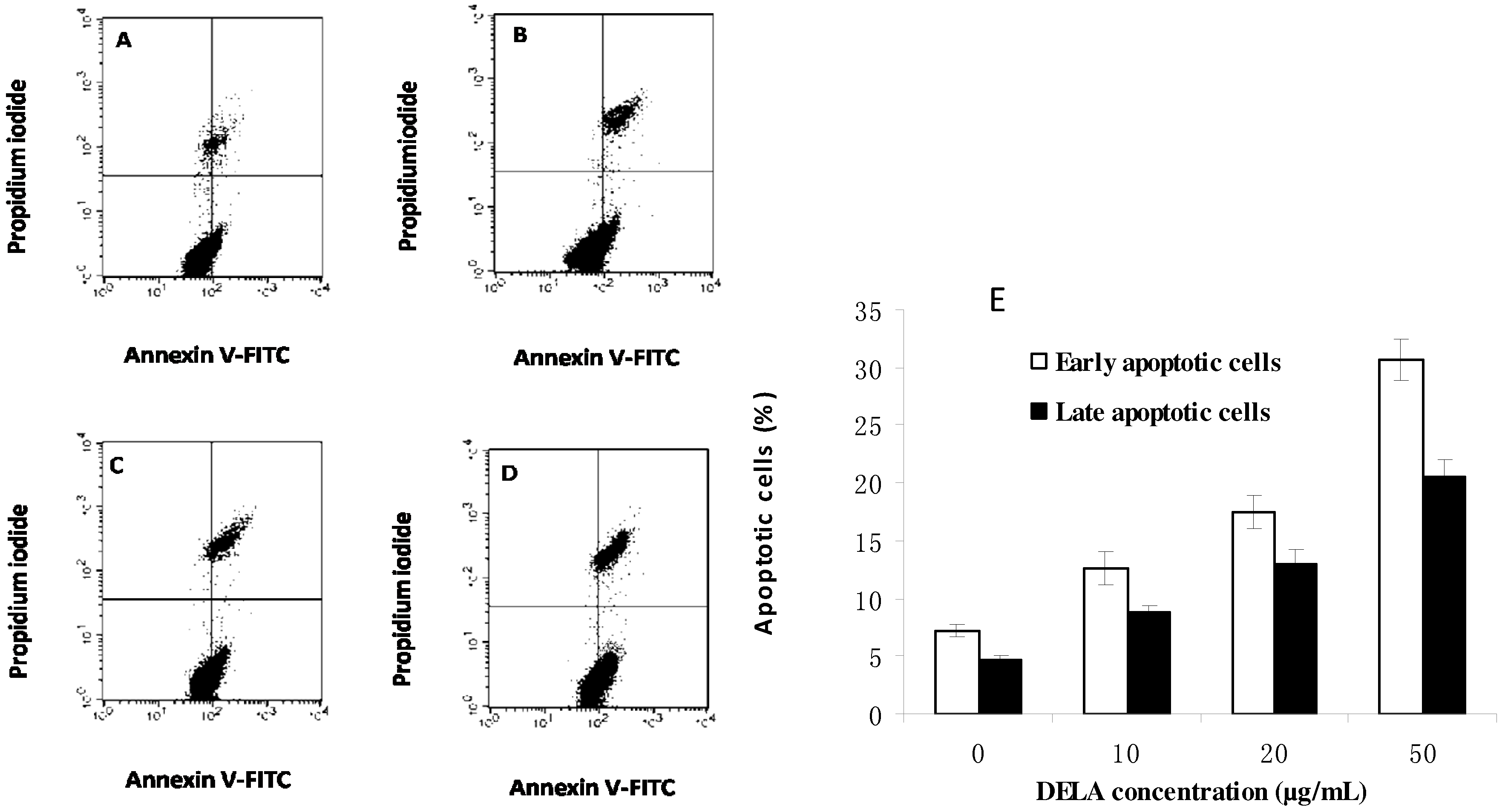
2.2. DELA-Induced Apoptosis in Murine Sarcoma S180 Cells
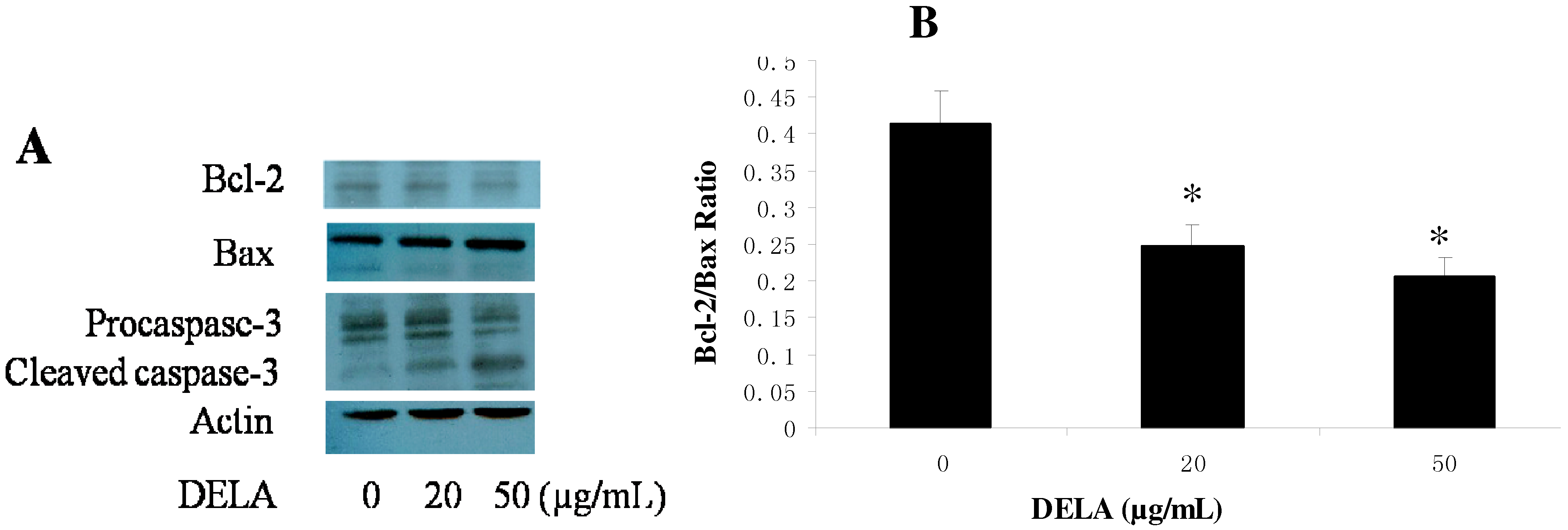
2.3. DELA-Induced Procaspase-3 Protein Cleaved in Murine Sarcoma S180 Cell


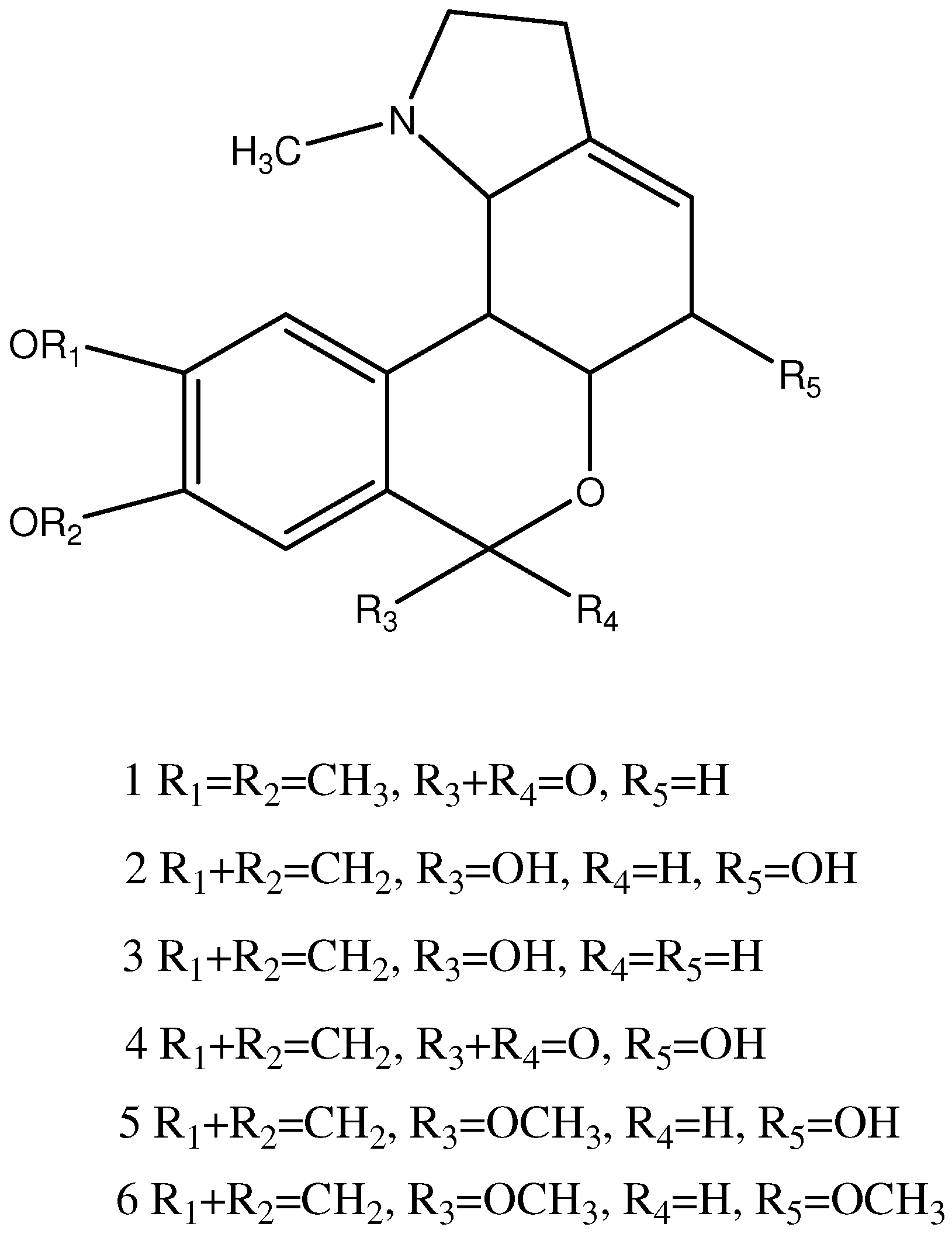
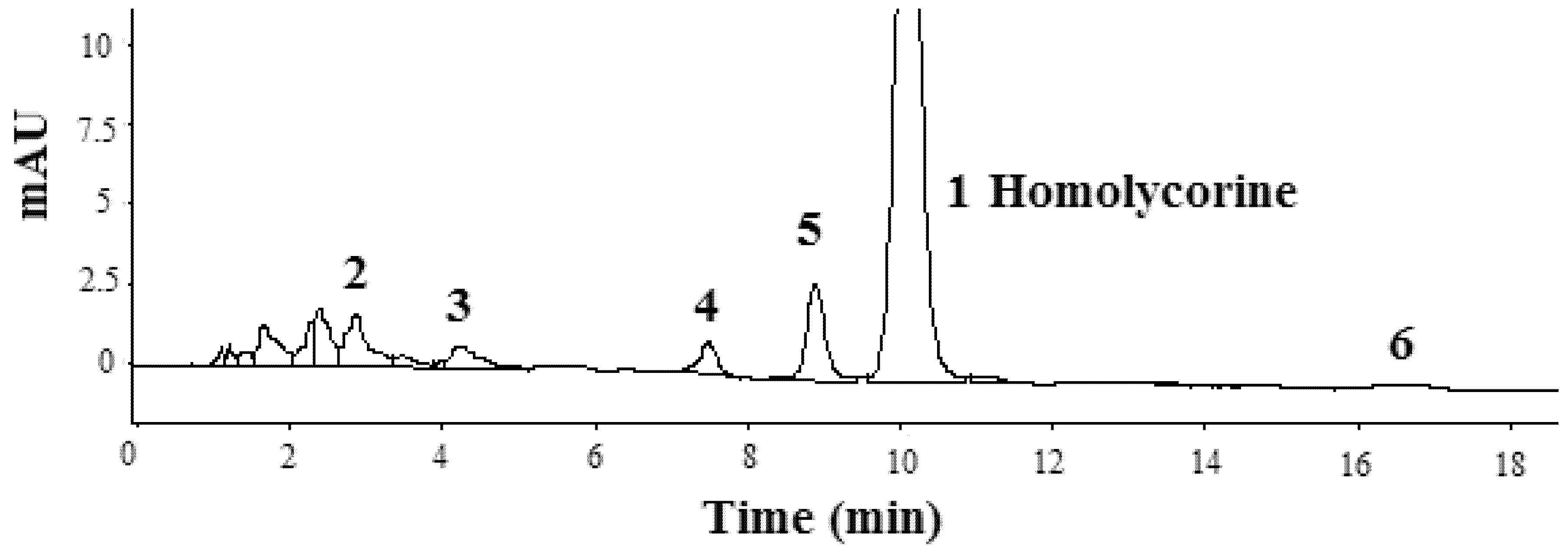
2.4. Structural Elucidation of Isolated Compound
| Peak No. | Identification | t R (min) | [M+H]+ | MS2 Characteristic ions (m/z) |
|---|---|---|---|---|
| 2 | 2α-Hydroxyoduline | 3.2 | 318 | 318, 282, 253, 193 |
| 3 | Oduline [23] | 4.4 | 302 | 302, 284, 256, 226, 193, 177, 145, 110, 94 |
| 4 | Hippeastrine [22] | 7.8 | 316 | 316, 298, 280, 239, 223, 191, 126, 96 |
| 5 | 2α-Hydroxy-6-O-methyloduline [22] | 9.2 | 332 | 332, 314, 300, 282, 257, 239, 223, 201, 175, 96 |
| 6 | 2α-Methoxy-6-O-methyloduline | 16.9 | 346 | 346, 314, 282, 264, 239, 199, 175 |

3. Experimental
3.1. Materials
3.2. Extraction from L. aurea and Isolation of Anticancer Compounds

3.3. Cell Cultures
3.4. MTT Assay
3.5. Anticancer Activity in Vivo
3.6. Flow Cytometry Assay
3.7. Western Blotting Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Yang, B.F.; Lu, Y.J.; Wang, Z.G. MicroRNAs and apoptosis: Implications in the molecular therapy of human disease. Clin. Exp. Pharmacol. Physiol. 2009, 36, 951–960. [Google Scholar]
- Schwartz, P.S.; Waxman, D.J. Cyclophosphamide induces caspase 9-dependent apoptosisin 9L tumor cells. Mol. Pharmacol. 2001, 60, 1268–1279. [Google Scholar]
- Chamorro-Cevallos, G.; Garduno-Siciliano, L.; Barron, B. Chemoprotective effect of Spirulina (Arthrospira) against cyclophosphamide-induced mutagenicity in mice. Food. Chem. Toxicol. 2008, 46, 567–574. [Google Scholar]
- Thi Ngoc Tram, N.; Titorenkova, T.V.; St Bankova, V.; Handjieva, N.V.; Popov, S.S. Crinum L. Amaryllidaceae. Fitoterapia 2002, 73, 183–208. [Google Scholar] [CrossRef]
- Liu, J.; Hu, W.X.; He, L.F; Ye, M.; Li, Y. Effects of lycorine on HL-60 cells via arresting cell cycle and inducing apoptosis. FEBS Lett. 2004, 578, 245–250. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Tang, L.J.; Shi, Y.W.; Ren, W.; Hu, W.X. Apoptosis induced by lycorine in KM3 cells is associated with the G0/G1 cell cycle arrest. Oncol. Rep. 2007, 17, 377–384. [Google Scholar]
- Liu, X.; Jiang, J.; Jiao, X.; Wu, Y.; Lin, J.; Cai, Y. Lycorine induces apoptosis and down-regulation of Mcl-1 in human leukemia cells. Cancer Lett. 2009, 274, 16–24. [Google Scholar]
- Yuan, H.; Song, J.; Li, X.; Li, N.; Dai, J. Immunomodulation and antitumor activity of [kappa]-carrageenan oligosaccharides. Cancer Lett. 2006, 243, 228–234. [Google Scholar]
- Wang, W.; Guo, Q.L.; You, Q.D.; Zhang, K.; Yang, Y.; Yu, J.; Liu, W.; Zhao, L.; Gu, H.Y.; Hu, Y.; Tan, Z.; et al. The anticancer activities of wogonin in murine sarcoma S180 both in vitro and in vivo. Biol. Pharm. Bull. 2006, 29, 1132–1137. [Google Scholar] [CrossRef]
- Cesta, M.F. Normal structure, function, and histology of the spleen. Toxicol. Pathol. 2006, 34, 455–465. [Google Scholar] [CrossRef]
- Feng, X.; Zhang, L.; Zhu, H. Comparative anticancer and antioxidant activities of different ingredients of Ginkgo biloba extract (EGb 761). Planta Med. 2009, 75, 792–796. [Google Scholar] [CrossRef]
- Wang, Y.; Meng, Q.; Qiao, H.; Jiang, H.; Sun, X. Role of the spleen in cyclophosphamide-induced hematosuppression and extramedullary hematopoiesis in mice. Arch. Med. Res. 2009, 40, 249–255. [Google Scholar]
- Zeng, S.; Liu, W.; Nie, F.; Zhao, Q.; Rong, J.; Wang, J.; Tao, L.; Qi, Q.; Lu, N.; Li, Z.; et al. LYG-202, a new flavonoid with a piperazine substitution, shows antitumor effects in vivo and in vitro. Biochem. Biophys. Res. Commun. 2009, 385, 551–556. [Google Scholar] [CrossRef]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar]
- Rinner, B.; Kretschmer, N.; Knausz, H.; Mayer, A.; Boechzelt, H.; Hao, X.J.; Heubl, G.; Efferth, T.; Schaider, H.; Bauer, R. A petrol ether extract of the roots of Onosma paniculatum induces cell death in a caspase dependent manner. J. Ethnopharmacol. 2010, 129, 182–188. [Google Scholar] [CrossRef]
- Wu, Y.; Fan, Q.; Lu, N.; Tao, L.; Gao, Y.; Qi, Q.; Guo, Q. Breviscapine‐induced apoptosis of human hepatocellular carcinoma cell line HepG2 was involved in its antitumor activity. Phytother. Res. 2010, 24, 1188–1194. [Google Scholar]
- Buolamwini, J.K. Novel anticancer drug discovery. Curr. Opin. Chem. Biol. 1999, 3, 500–509. [Google Scholar]
- Reed, J.C.; Stein, C.; Subasinghe, C.; Haldar, S.; Croce, C.; Yum, S.; Cohen, J. Antisense-mediatedinhibition of BCL2 protooncogene expression and leukemic cell growth and survival: Comparisons of phosphodiester and phosphorothioate oligodeoxynucleotides. Cancer Res. 1990, 50, 6565. [Google Scholar]
- Manna, S.; Banerjee, S.; Mukherjee, S.; Das, S.; Panda, C.K. Epigallocatechin gallate induced apoptosis in Sarcoma180 cells in vivo: Mediated by p53 pathway and inhibition in U1B, U4-U6 UsnRNAs expression. Apoptosis 2006, 11, 2267–2276. [Google Scholar]
- Li, Q.; Jiang, C.; Zu, Y.; Song, Z.; Zhang, B.; Meng, X.; Qiu, W.; Zhang, L. SFE-CO2 extract from Typhonium giganteum Engl. Tubers, induces apoptosis in human hepatoma SMMC-7721 cells involvement of a ROS-mediated mitochondrial pathway. Molecules 2011, 16, 8228–8243. [Google Scholar] [CrossRef]
- Schnoes, H.; Smith, D.; Burlingame, A.; Jeffs, P.; Döpke, W. Mass spectra of amaryllidaceae alkaloids: The lycorenine series. Tetrahedron 1968, 24, 2825–2837. [Google Scholar]
- Almanza, G.R.; Fernández, J.M.; Wakori, E.W.T.; Viladomat, F.; Codina, C.; Bastida, J. Alkaloids from Narcissus cv. Salome. Phytochemistry 1996, 43, 1375–1378. [Google Scholar] [CrossRef]
- Kreh, M.; Matusch, R.; Witte, L. Capillary gas chromatography-mass spectrometry of Amaryllidaceae alkaloids. Phytochemistry 1995, 38, 773–776. [Google Scholar]
- Hartwell, J.L. Plants used against cancer. A survey. Lloydia 1967, 30, 379–436. [Google Scholar]
- Hartwell, J.L. Plants Used Against Cancer; Quarterman Publication: Lawrencee, MA, USA, 1982. [Google Scholar]
- Cook, J.W.; Loudon, J.D. Chapter XI Alkaloids of the Amaryllidaceae. In The Alkaloids:Chemistry and Physiology; Academic Press: New York, NY, USA, 1952; Volume 2, pp. 331–352. [Google Scholar]
- Evidente, A.; Kireev, A.S.; Jenkins, A.R.; Romero, A.E.; Steelant, W.F.; Van Slambrouck, S.; Kornienko, A. Biological evaluation of structurally diverse Amaryllidaceae alkaloids and their synthetic derivatives: Discovery of novel leads for anticancer drug design. Planta Med. 2009, 75, 501–507. [Google Scholar]
- Berkov, S.; Bastida, J.; Sidjimova, B.; Viladomat, F.; Codina, C. Phytochemical differentiation of Galanthus nivalis and Galanthus elwesii (Amaryllidaceae): A case study. Biochem. Syst. Ecol. 2008, 36, 638–645. [Google Scholar]
- Yui, S.; Mikami, M.; Kitahara, M.; Yamazaki, M. The inhibitory effect of lycorine on tumor cell apoptosis induced by polymorphonuclear leukocyte-derived calprotectin. Immunopharmacology 1998, 40, 151–162. [Google Scholar]
- Ceriotti, G. Narciclasine: An antimitotic substance from Narcissus bulbs. Nature 1967, 213, 595–596. [Google Scholar] [CrossRef]
- Carrasco, L.; Fresno, M.; Vazquez, D. Narciclasine: An antitumour alkaloid which blocks peptide bond formation by eukaryotic ribosomes. FEBS Lett. 1975, 52, 236–239. [Google Scholar]
- Dumont, P.; Ingrassia, L.; Rouzeau, S.; Ribaucour, F.; Thomas, S.; Roland, I.; Darro, F.; Lefranc, F.; Kiss, R. The Amaryllidaceae isocarbostyril narciclasine induces apoptosis by activation of the death receptor and/or mitochondrial pathways in cancer cells but not in normal fibroblasts. Neoplasia 2007, 9, 766. [Google Scholar]
- Pettit, G.R.; Gaddamidi, V.; Cragg, G.M. Antineoplastic agents, 105. Zephyranthes grandiflora. J. Nat. Prod. 1984, 47, 1018–1020. [Google Scholar] [CrossRef]
- Kekre, N.; Griffin, C.; McNulty, J.; Pandey, S. Pancratistatin causes early activation of caspase-3 and the flipping of phosphatidyl serine followed by rapid apoptosis specifically in human lymphoma cells. Cancer Chemother. Pharmacol. 2005, 56, 29–38. [Google Scholar]
- McNulty, J.; Nair, J.J.; Codina, C.; Bastida, J.; Pandey, S.; Gerasimoff, J.; Griffin, C. Selective apoptosis-inducing activity of crinum-type Amaryllidaceae alkaloids. Phytochemistry 2007, 68, 1068–1074. [Google Scholar]
- Liu, J.; Li, Y.; Ren, W.; Hu, W.X. Apoptosis of HL-60 cells induced by extracts from Narcissus tazetta var. chinensis. Cancer Lett. 2006, 242, 133–140. [Google Scholar] [CrossRef]
- Van Goietsenoven, G.; Hutton, J.; Becker, J.P.; Lallemand, B.; Robert, F.; Lefranc, F.; Pirker, C.; Vandenbussche, G.; Van Antwerpen, P.; Evidente, A.; et al. Targeting of eEF1A with Amaryllidaceae isocarbostyrils as a strategy to combat melanomas. FASEB J. 2010, 24, 4575–4584. [Google Scholar] [CrossRef]
- Van Goietsenoven, G.; Andolfi, A.; Lallemand, B.; Cimmino, A.; Lamoral-Theys, D.; Gras, T.; Abou-Donia, A.; Dubois, J.; Lefranc, F.; Mathieu, V.; et al. Amaryllidaceae alkaloids belonging to different structural subgroups display activity against apoptosis-resistant cancer cells. J. Nat. Prod. 2010, 73, 1223–1227. [Google Scholar] [CrossRef]
- Luchetti, G.; Johnston, R.; Mathieu, V.; Lefranc, F.; Hayden, K.; Andolfi, A.; Lamoral-Theys, D.; Reisenauer, M.R.; Champion, C.; Pelly, S.C.; et al. Bulbispermine: A crinine-type Amaryllidaceae alkaloid exhibiting cytostatic activity toward apoptosis-resistant glioma cells. Chem. Med. Chem. 2012. [Google Scholar] [CrossRef]
- Weniger, B.; Italiano, L.; Beck, J.; Bastida, J.; Bergoñon, S.; Codina, C.; Lobstein, A.; Anton, R. Cytotoxic activity of Amaryllidaceae alkaloids. Planta Med. 1995, 61, 77–79. [Google Scholar]
- Hawksworth, W.; Jeefs, P.; Tidd, B.; Toube, T. 357. The alkaloids of the amaryllidaceae. Part XII. The aromatic oxygenation patterns and stereochemistry of some trioxyaryl alkaloids of the hemiacetal and lactone series. J. Chem. Soc. 1965, 1991–2001. [Google Scholar]
- Antoun, M.D.; Mendoza, N.T.; Ríos, Y.R.; Proctor, G.R.; Wickramaratne, D.B.; Pezzuto, J.M.; Kinghorn, A.D. Cytotoxicity of Hymenocallis expansa alkaloids. J. Nat. Prod. 1993, 56, 1423–1425. [Google Scholar] [CrossRef]
- Evidente, A.; Kornienko, A. Anticancer evaluation of structurally diverse Amaryllidaceae alkaloids and their synthetic derivatives. Phytochem. Rev. 2009, 8, 449–459. [Google Scholar]
- Szlavik, L.; Gyuris, A.; Minarovits, J.; Forgo, P.; Molnar, J.; Hohmann, J. Alkaloids from Leucojum vernum and antiretroviral activity of Amaryllidaceae alkaloids. Planta Med. 2004, 70, 871–873. [Google Scholar]
- Jeffs, P.W.; Abou-Donia, A.; Campau, D.; Staiger, D. Alkaloids of the Amaryllidaceae. 27. Structures of 9-O-demethylhomolycorine and 5α-hydroxyhomolycorine. Alkaloids of Crinum defixum, C. scabrum, and C. latifolium. Assignment of aromatic substitution patterns from 1H-coupled carbon-13 spectra. J. Org. Chem. 1985, 50, 1732–1737. [Google Scholar] [CrossRef]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liao, N.; Ao, M.; Zhang, P.; Yu, L. Extracts of Lycoris aurea Induce Apoptosis in Murine Sarcoma S180 Cells. Molecules 2012, 17, 3723-3735. https://doi.org/10.3390/molecules17043723
Liao N, Ao M, Zhang P, Yu L. Extracts of Lycoris aurea Induce Apoptosis in Murine Sarcoma S180 Cells. Molecules. 2012; 17(4):3723-3735. https://doi.org/10.3390/molecules17043723
Chicago/Turabian StyleLiao, Na, Mingzhang Ao, Peng Zhang, and Longjiang Yu. 2012. "Extracts of Lycoris aurea Induce Apoptosis in Murine Sarcoma S180 Cells" Molecules 17, no. 4: 3723-3735. https://doi.org/10.3390/molecules17043723