Luffa echinata Roxb. Induces Human Colon Cancer Cell (HT-29) Death by Triggering the Mitochondrial Apoptosis Pathway

Abstract
:1. Introduction
2. Results and Discussion
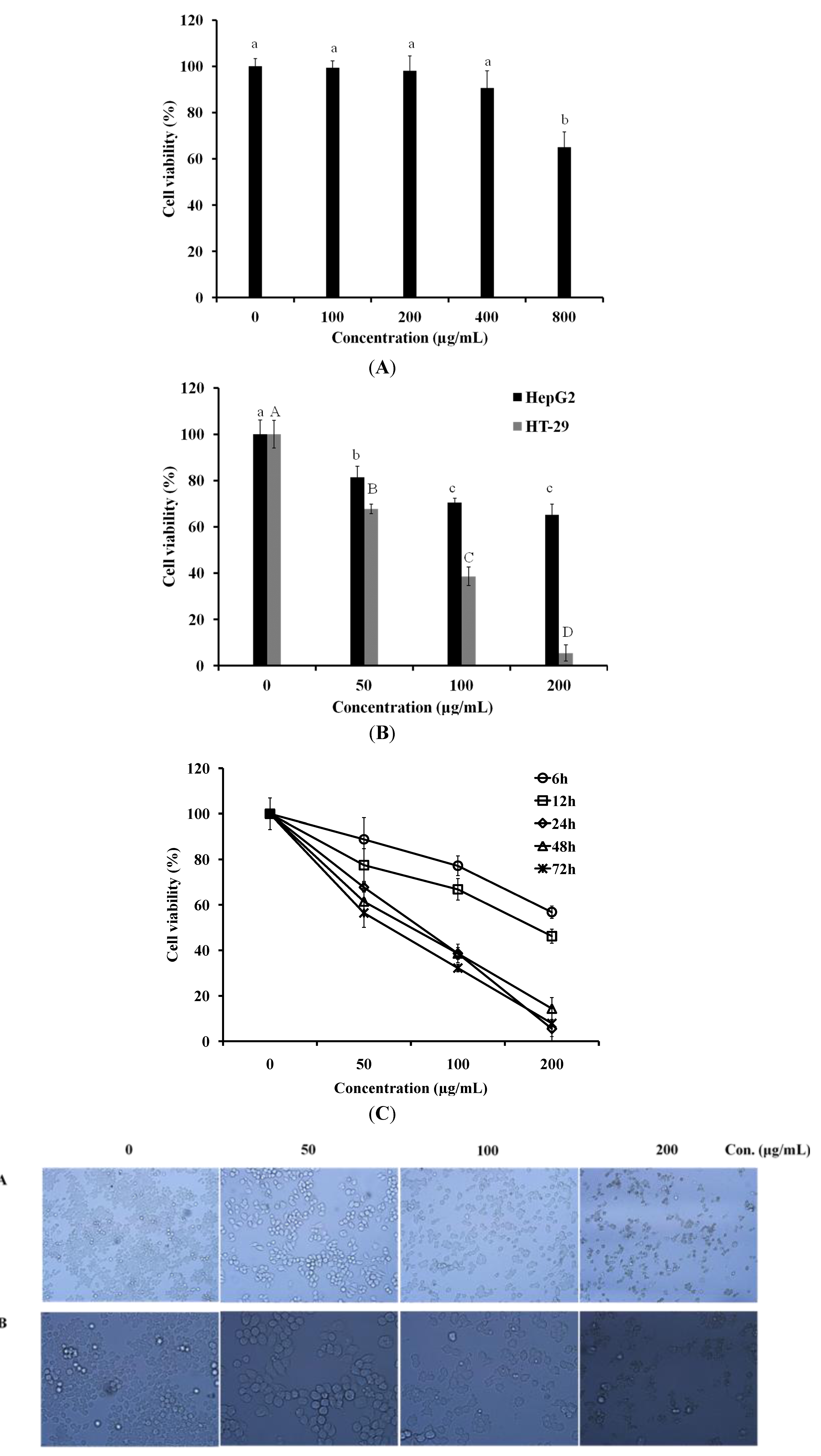
2.1. LER Inhibits the Proliferation of HT-29 Cells

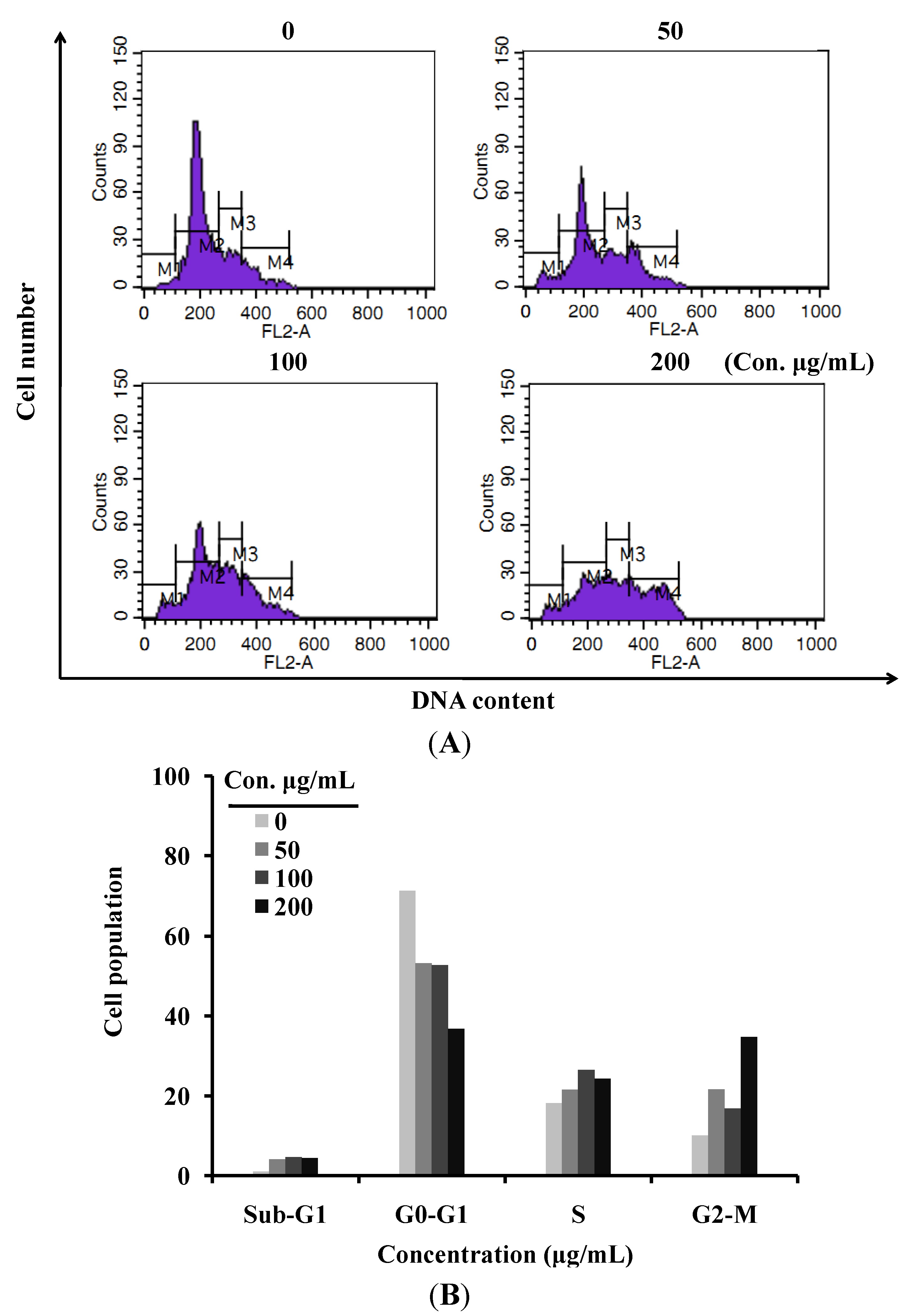
2.2. LER Extract Induces a G2/M Cell Cycle Arrest in HT-29 Cells

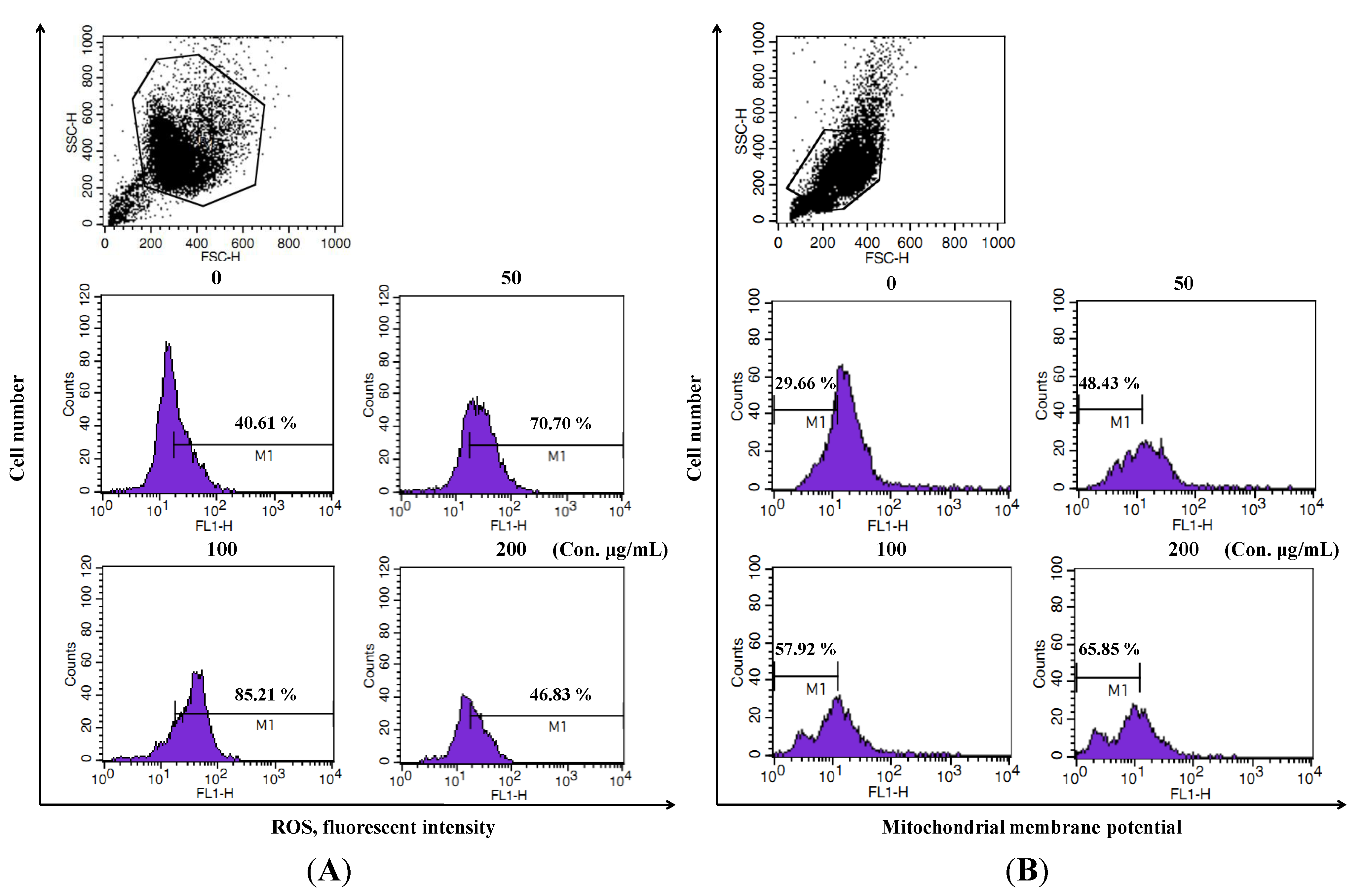
2.3. LER Extract Induces Cell Death by ROS Generation and Mitochondrial Membrane Potential (MMP) Lose
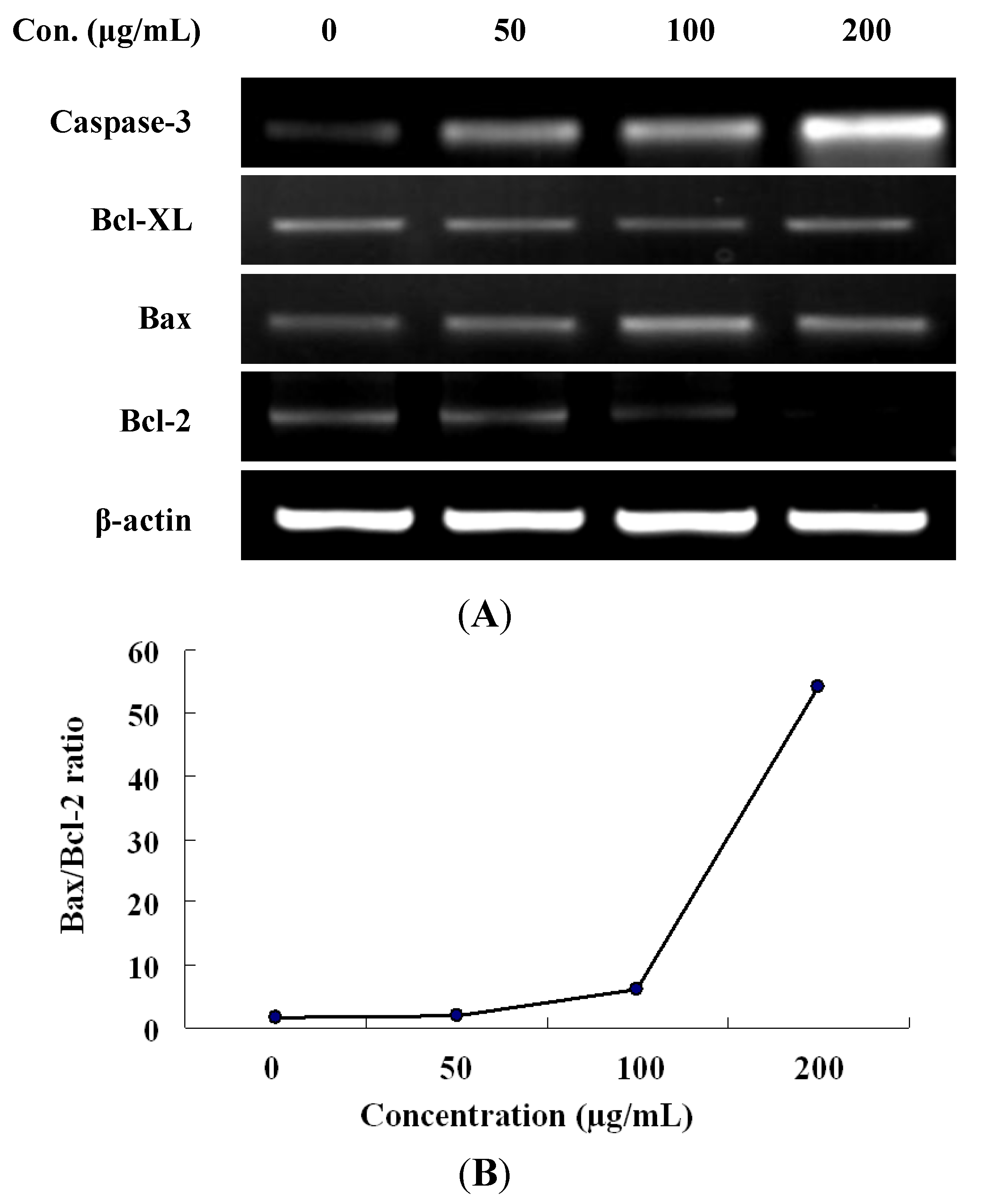
2.4. Expression of Apoptotic-Related Genes in LER Extract-Treated Cells


2.5. Discussion
3. Experimental
3.1. Plant Material and Extract Preparation
3.2. Cell Lines and Cell Culture
3.3. Cytotoxicity and Anti-Proliferation Test
3.4. Light Microscopic Examination
3.5. Cell Cycle Analysis
3.6. Measurement of Reactive Oxygen Species (ROS) Level
3.7. Measurement of Mitochondrial Membrane Potential (MMP) Level
3.8. RNA Isolation and Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Forward primer (5'–3') | Forward primer (5'–3') |
|---|---|---|
| capspase-3 | TCACAGCAAAAGGAGCAGTTT | CGTCAAAGGAAAAGGACTCAA |
| Bcl-XL | CCAGAAGGGACTGAATCG | CCTTGTCACGCTTTCCAC |
| Bax | TCCACCAAGAAGCTGAGCGA | GTCCAGCCCATGATGGTTCT |
| Bcl-2 | TGTGGCCTTCTTTGAGTTCG | TCACTTGTGGCTCAGATAGG |
| β-actin | TCACCCTGAAGTACCCCATC | CCATCTCTTGCTGCAAGTCC |
3.9. Statistical Analyses
4. Conclusions
Acknowledgements
References and Notes
- Feeley, T.H.; Cooper, J.; Foels, T.; Mahoney, M.C. Efficacy expectations for colorectal cancer screening in primary care: Identifying barriers and facilitators for patients and clinicians. Health Commun. 2009, 24, 304–315. [Google Scholar] [CrossRef]
- Urruticoechea, A.; Alemany, R.; Balart, J.; Villanueva, A.; Vinals, F.; Capella, G. Recent advances in cancer therapy: An overview. Curr. Pharm. Des. 2010, 16, 3–10. [Google Scholar] [CrossRef]
- Matsuhashi, N.; Saio, M.; Matsuo, A.; Sugiyama, Y.; Saji, S. Apoptosis induced by 5-fluorouracil, cisplatin and paclitaxel are associated with p53 gene status in gastric cancer cell lines. Int. J. Oncol. 2005, 26, 1563–1567. [Google Scholar]
- Hirao, M.; Fujitani, K.; Tsujinaka, T. Phase I study of the combination of nedaplatin, adriamycin and 5-fluorouracil for treatment of advanced esophageal cancer. Dis. Esophagus 2004, 17, 247–250. [Google Scholar] [CrossRef]
- Legarza, K.; Yang, L.X. New molecular mechanisms of action of camptothecin-type drugs. Anticancer Res. 2006, 26, 3301–3305. [Google Scholar]
- Slater, T.F.; Sawyer, B.; Strauli, U.D. Studies on succinate-tetrazolium reductase systems. III. Points of coupling of four different tetrazolium salts. Biochim. Biophys. Acta 1963, 77, 383–393. [Google Scholar] [CrossRef]
- Fulda, S.; Debatin, K.M. Apoptosis pathways in neuroblastoma therapy. Cancer Lett. 2003, 197, 131–135. [Google Scholar] [CrossRef]
- Hu, W.; Lee, S.K.; Jung, M.J.; Heo, S.I.; Hur, J.H.; Wang, M.H. Induction of cell cycle arrest and apoptosis by the ethyl acetate fraction of Kalopanax pictus leaves in human colon cancer cells. Bioresour. Technol. 2010, 101, 9366–9372. [Google Scholar]
- Kirtikar, K.R.; Basu, B.D. Indian Medicinal Plants; L.M. Basu: Allahabad, India, 1933; Volume 2, p. 587. [Google Scholar]
- Chaudhary, G.R.; Sharma, V.N.; Siddiqui, S. Amariin: A bitter constituent of Luffa species. 1951, 10B, 26. [Google Scholar]
- Bhatt, R.H.; Khurana, M.L. Studies on Luffa echinata. Indian J. Pharm. 1957, 19, 208–210. [Google Scholar]
- Khorana, M.L.; Raisinghani, K.H. Studies of Luffa echinata III. The oil and the saponin. J. Pharmacol. Sci. 1961, 50, 687–689. [Google Scholar] [CrossRef]
- Lavie, D.; Shvo, Y.; Gottlieb, O.R.; Desai, R.B.; Khorana, M.L. The occurrence of 2-epicucurbitacin B in Luffa echinata. J. Chem. Soc. 1962, 3, 3259–3260. [Google Scholar]
- Mesbah, U.A.; Haque, M.E.; Sutradhar, R.K. Bitter principe of Luffa echinata. Phytochemistry 1994, 36, 421–423. [Google Scholar]
- Lauria, P.; Sharma, V.N.; Vanjani, S.; Sangal, B.C. Role of Luffa echinata in liver injury and its other pharmacological actions. Indian J. Pharmacol. 1976, 8, 129–133. [Google Scholar]
- Bhatt, R.H.; Khorana, M.L.; Gaitonde, B.B.; Raiker, K.P.; Patel, J.R.; Kekre, M.S. Diuretic properties of saponin of Luffa echinata Roxb. J. Gr. Hosp. Grant. Med. Coll. 1958, 3, 110. [Google Scholar]
- Aswal, B.S.; Bhakuni, D.S.; Goel, A.K.; Kar, K.; Mehrotra, B.N.; Mukherjee, K.C. Screening of Indian plants for biological activity. Part X. Indian J. Exp. Biol. 1984, 22, 312–332. [Google Scholar]
- Nadkarni, K.M. Dr. K. M. Nadkarni’s Indian Materia Medica, Part III, 3rd ed; Popular Prakashan: Mumbai, India, 1976; p. 268. [Google Scholar]
- Bhatt, R.H.; Khorana, M.L.; Patel, J.R.; Gaitonde, B.B.; Kekre, M.S. Pharmacological studies of saponins of the fruits of Luffa echinata Roxb and seeds of Trigonella foenum-graecum Linn. Indian J. Physiol. Pharmacol. 1958, 2, 309. [Google Scholar]
- Ahmed, B.; Alam, T.; Khan, S. Hepatoprotective activity of Luffa echinata fruits. J. Ethnopharmacol. 2001, 76, 187–189. [Google Scholar] [CrossRef]
- Yang, H.L.; Chen, C.S.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Chen, C.C.; Hseu, T.H.; Kuo, C.T.; Hseu, Y.C. Growth inhibition and induction of apoptosis in MCF-7 breast cancer cells by Antrodia camphorata. Cancer Lett. 2006, 231, 215–227. [Google Scholar] [CrossRef]
- Mossman, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Debatin, K.M.; Krammer, P.H. Death receptors in chemotherapy and cancer. Oncogene 2004, 23, 2950–2966. [Google Scholar] [CrossRef]
- Kaufman, S.H. Induction of endonucleolytic DNA cleavage in human acute myelogenous leukemia cells by etoposide, camptothecin and other cytotoxic anticancer drugs: A cautionary note. Cancer Res. 1989, 49, 5870–5878. [Google Scholar]
- Huschtscha, L.I.; Bartier, W.A.; Ross, C.E.A.; Tattersall, M.H. Characteristics in cancer death after exposure to cytotoxic drugs in vitro. Br. J. Cancer 1996, 73, 54–60. [Google Scholar] [CrossRef]
- Hoyt, M.A. A new checkpoints take shape. Nat. Cell Biol. 2004, 6, 801–803. [Google Scholar] [CrossRef]
- Kumar, N.; Afeyan, R.; Kim, H.D.; Lauffenburger, D.A. Multipathway model enables prediction of kinase inhibitor cross-talk effects on migration of Her2-overexpressing mammary epithelial cells. Mol. Pharmacol. 2008, 73, 1668–1678. [Google Scholar] [CrossRef]
- Hsu, S.C.; Ou, C.C.; Li, J.W.; Chuang, T.C.; Kuo, H.P.; Liu, J.Y.; Chen, C.S.; Lin, S.C.; Su, C.H.; Kao, M.C. Ganoderma tsugae extracts inhibit colorectal cancer cell growth via G2/M cell cycle arrest. J. Ethnopharmacol. 2008, 120, 394–401. [Google Scholar] [CrossRef]
- Wang, J.; Wu, A.; Xu, Y.; Liu, J.; Qian, X. M2-A induces apoptosis and G2-M arrest via inhibiting PI3 K/Akt pathway in HL60 cells. Cancer Lett. 2009, 283, 193–202. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Shen, X.; Chao, X.; Ho, C.C.; Cheng, X.L.; Zhang, Y.; Lin, R.C.; Du, K.J.; Luo, W.J.; Chen, J.Y.; et al. Ergosta-4, 6, 8(14), 22-tetraen-3-one induces G2/M cell cycle arrest and apoptosis in human hepatocellular carcinoma HepG2 cells. Biochim. Biophys. Acta 2011, 1810, 384–390. [Google Scholar] [CrossRef]
- Hadfield, J.A.; Ducki, S.; Hirst, N.; McGown, A.T. Tubulin and microtubules as targets for anticancer drugs. Prog. Cell Cycle Res. 2003, 5, 309–325. [Google Scholar]
- Boonstra, J.; Post, J.A. Molecular events associated with reactive oxygen species and cell cycle progression in mammalian cells. Gene 2004, 337, 1–13. [Google Scholar] [CrossRef]
- Wu, W.S. The signaling mechanism of ROS in tumor progression. Cancer Metastasis Rev. 2006, 25, 695–705. [Google Scholar] [CrossRef]
- Kamata, H.; Hirata, H. Redox regulation of cellular signalling. Cell. Signal. 1999, 11, 1–14. [Google Scholar]
- Wang, X.; Sharma, R.K.; Gupta, A.; George, V.; Thomas, A.J.; Falcone, T.; Agarwal, A. Alterations in mitochondria membrane potential and oxidative stress in infertile men: A prospective observational study. Fertil. Steril. 2003, 80, 844–850. [Google Scholar] [CrossRef]
- Cummins, J.M.; Jequiers, A.M.; Kan, R. Molecular biology of human male infertility: Links with aging, mitochondrial genetics and oxidative stress. Mol. Reprod. Dev. 1994, 37, 345–362. [Google Scholar] [CrossRef]
- Kim, W.H.; Park, W.B.; Gao, B.; Jung, M.H. Critical role of reactive oxygen species and mitochondrial membrane potential in Korean mistletoe lectin-induced apoptosis in human hepatocarcinoma cells. Mol. Pharmacol. 2004, 66, 1383–1396. [Google Scholar] [CrossRef]
- Voisine, C.; Craig, E.A.; Zufall, N.; von Ahsen, O.; Pfanner, N.; Voos, W. The protein import motor of mitochondria: Unfolding and trapping of preproteins are distinct and separable functions of matrix Hsp70. Cell 1999, 97, 565–574. [Google Scholar] [CrossRef]
- Haupt, S.; Berger, M.; Goldberg, Z.; Haupt, Y. Apoptosis—The p53 network. J. Cell Sci. 2003, 116, 4077–4085. [Google Scholar] [CrossRef]
- Janicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar]
- Charles, A.K.; Hisheh, S.; Liu, D.; Rao, R.M.; Waddell, B.J.; Dickinson, J.E.; Rao, A.J.; Dharmarajan, A.M. The expression of apoptosis related genes in the first trimester human placenta using a short term in vitro model. Apoptosis 2005, 10, 135–140. [Google Scholar] [CrossRef]
- Lee, W.R.; Shen, S.C.; Lin, H.Y.; Hou, W.C.; Yang, L.L.; Chen, Y.C. Wogonin and fisetin induce apoptosis in human promyeloleukemic cells, accompanied by adecrease of reactive oxygen species, and activation of caspase 3 and Ca2+-dependent endonuclease. Biochem. Pharmacol. 2002, 63, 225–236. [Google Scholar]
- Chinnaiyan, A.M.; O’Rourke, K.; Lane, B.R.; Dixit, V.M. Interaction of CED-4 with CED-3 and CED-9: A molecular framework for cell death. Science 1997, 275, 1122–1126. [Google Scholar] [CrossRef]
- Cheng, E.H.Y.; Kirsch, D.G.; Clem, R.J.; Ravi, R.; Kastan, M.B.; Bedi, A.; Ueno, K.; Hardwick, J.M. Conversion of Bcl-2 to a Bax-like death effector by caspases. Science 1997, 278, 1966–1968. [Google Scholar]
- Sample Availability: Not available.
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shang, L.-H.; Li, C.-M.; Yang, Z.-Y.; Che, D.-H.; Cao, J.-Y.; Yu, Y. Luffa echinata Roxb. Induces Human Colon Cancer Cell (HT-29) Death by Triggering the Mitochondrial Apoptosis Pathway. Molecules 2012, 17, 5780-5794. https://doi.org/10.3390/molecules17055780
Shang L-H, Li C-M, Yang Z-Y, Che D-H, Cao J-Y, Yu Y. Luffa echinata Roxb. Induces Human Colon Cancer Cell (HT-29) Death by Triggering the Mitochondrial Apoptosis Pathway. Molecules. 2012; 17(5):5780-5794. https://doi.org/10.3390/molecules17055780
Chicago/Turabian StyleShang, Li-Hua, Chun-Mei Li, Zhao-Yang Yang, De-Hai Che, Jing-Yan Cao, and Yan Yu. 2012. "Luffa echinata Roxb. Induces Human Colon Cancer Cell (HT-29) Death by Triggering the Mitochondrial Apoptosis Pathway" Molecules 17, no. 5: 5780-5794. https://doi.org/10.3390/molecules17055780